
Abstract
TRPA and TRPV ion channels are members of the transient receptor potential (TRP) cation channel superfamily, which mediates various sensory transductions. In Caenorhabditis elegans, the TRPV channels are known to affect chemosensation, while the TRPA-1 channel is associated with thermosensation and mechanosensation. We examined thermosensation, chemosensation, and osmosensation in strains lacking TRPA-1 or TRPV channels. We found that TRPV channel knockout worms exhibited similar behavioral deficits associated with thermotaxis as the TRPA-1 channel knockout, suggesting a dual role for TRPV channels. In contrast, chemosensation responses, assessed by both avoidance reversal behavior and NaCl osmosensation, were dependent on TRPV channels but seemed independent of TRPA-1 channel. Our findings suggest that, in addition to TRPA-1 channel, TRPV channels are necessary for thermotaxis and may activate, or modulate, the function of TRPA-1 channels. In contrast, TRPA-1 channels do not have a dual responsibility, as they have no functional role in odorant avoidance or osmosensation.
Keywords
Introduction
The transient receptor potential (TRP) cation channels belong to a gene superfamily which is present in all eukaryotes. These channels open and close in response to various stimuli, such as temperature, mechanical force, chemical, and osmotic pressure.1,2 TRPV channels have been associated with chemosensation, pain sensation, and the detection of osmotic pressure differences, 3 while TRPA channels have been associated with thermosensation and mechanosensation.4,5 TRPV-1 channel also functions as a heat sensor in vertebrates. 6 Accumulating evidence suggest a potential interaction between TRPA and TRPV channels. For example, TRPA-1 and TRPV-1 are known to be present in the human airway and are both involved in respiratory diseases. 7 11 Coexpression of TRPV-1 with TRPA-1 in vitro results in the modifications of the function of TRPA-1. 12 However, whether or not these channels interact in vivo and the potential effects of such interaction on animal physiology and behavior remain unclear. In this study, we use the nematode Caenorhabditis elegans as the model system to examine the functional overlap between TRPA-1 and TRPV channels and to determine the effects of interaction between these channels on different behaviors. Our results suggest that, in addition to TRPA-1 channel, TRPV channels are necessary for thermotaxis and may activate, or modulate, the function of TRPA-1 channels, while TRPA-1 channels do not have a dual responsibility as they have no functional role in odorant avoidance or osmosensation.
Materials and Methods
C. Elegans Strains
Three strains were provided by the Caenorhabditis Genetics Center, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440): wild-type Bristol strain N2, TRPA-1 channel knockout RB1052 trpa-1(ok999) IV, and TRPV triple-channel knockout FG125 ocr-2(ak47) osm-9(ky10) IV ocr-1(ak46) V. This latter strain was selected due to the complex expression patterns of TRPV channels and the probable redundancy of TRPV channels in C. elegans.13,14
Worms were maintained at 16° C on nematode growth medium (NGM) with an OP50 strain of Escherichia coli as the primary food source. The N2 strain was used in control experiments to validate both naive and learned responses.
Thermotaxis assay
A thermotaxis assay was performed to examine whether training worms to associate food with a lower temperature could be achieved with or without TRPA-1 or TRPV channels. First, the worms were trained in a 16°C incubator for 24 hours with an abundance of bacterial food. They were then transferred to the middle of larger test plates with temperature 26°C and 15°C setup at two opposite ends (experimental setup demonstrated in Fig. 1A), containing NGM without a bacterial food lawn for the thermotaxis assay. The NGM chunk used to transport them was removed after 15 minutes, and the temperature gradient was checked for every 15 minutes. The worms were allowed a total of 60 minutes to migrate on a lane in the test plate to their preferred temperature zone. A paralytic, 0.5 M sodium azide (NaN3) was added at both ends of the plate. The number of worms was counted, and the following equation was used to quantify the behavior: thermotaxis learning index = 100 × (worms moving toward 15°C - worms moving toward 26°C)/(worms moving toward 15°C + worms moving toward 26°C).

Thermotaxis behavior of C. elegans. (
Chemosensory assay
This assay was performed by presenting an individual worm with a repellant odorant and measuring the time it took to initiate a reversal movement. A sterilized platinum wire was dipped in a 1:10 dilution of 2-octanone in ethanol, or in 1-octanol, and then placed immediately in front of an isolated, freely moving adult worm on an agar plate. The time elapsing until the initiation of a reversal movement was recorded. 15
Osmotaxis assay
This assay was performed following the method of Kunitomo et al. 16 Briefly, a NaCl concentration gradient was established by placing two agar blocks at opposite ends of a Petri dish containing 50 mM NaCl in the absence of bacteria for 14 hours, with one agar block containing 0 mM NaCl and the other containing 150 mM NaCl. The worms were cultivated on a 25 mM NaCl NGM plate until transferred to the gradient plate and allowed to forage for 40 minutes. A total of 1 μL of paralytic 0.5 M sodium azide (NaN3) was added at both ends of the plate to trap worms as they arrived. The number of worms was counted, and the following equation was used to quantify the behavior: osmotaxis index = 100 × (worms moving toward 0 mM NaCl - worms moving toward 150 mM NaCl)/(worms moving toward 0 mM NaCl + worms moving toward 150 mM NaCl).
Statistical analysis
All values are reported as means ± SEM. Statistical differences were derived from one-way analysis of variance with post hoc Tukey's HSD test using IBM SPSS.
Results
We compared naive and learned behaviors of two knockout strains of C. elegans with those of wild-type worms. One strain, RB1052, is the TRPA-1 channel knockout. The other strain, FG125, is the triple TRPV channel (osm-9, ocr-1, and ocr-2) knockout.
A thermotaxis assay was used to test if training the worms to associate food with a lower temperature could be achieved with or without TRPA-1 or TRPV channels. We compared worms that went through training at 15°C for a 24-hour period and untrained wild-type worms. The thermotaxis index of the trained wild-type N2 worms was more than twofold that of the untrained N2 worms (P < 0.05; Table 1 and Fig. 1B), indicating that the thermotaxis training was successful. With the knockout strains, the thermotaxis indices of both RB1052 and FG125 strains were significantly lower than the learning index of the wild type (P < 0.05; Table 1 and Fig. 1C). Additionally, there was no significant difference between the two knockout strains (P = 0.946).
Behavioral measurements of C. elegans.
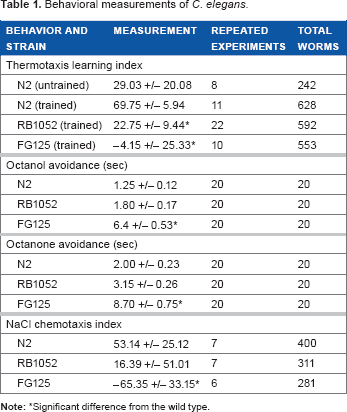
Significant difference from the wild type.
To assess chemosensory responses, two volatile odorants, such as 1-octanol and 2-octanone, both of which are repellant odorants to C. elegans, were used to induce avoidance reversal movement. When naive worms were presented with 1-octanol (Table 1 and Fig. 2A), the FG125 strain showed a significant increase in time to initiate reversal compared to the wild type (P < 0.001). However, the RB1052 strain only showed a slight increase in time to initiate reversal, which is not significantly different from the wild type (P = 0.47). Similar results were observed with 2-octanone (Table 1 and Fig. 2B). FG125 strain showed a significant increase in time to initiate reversal compared to the wild type (P < 0.001), but RB1052 strain only showed a slight increase in time to initiate reversal, which is not significantly different from the wild-type (P = 0.21).

Behavioral tests related to the chemical sensing of C. elegans. FG125 (TRPV knockout) strain showed a significant difference in reversal time when presented with 1-octanol (
We also used an NaCl osmosensory assay to examine whether the worms’ ability to associate food source with osmotic pressure of NaCl in the NGM could be achieved with or without the TRPA-1 or TRPV channels. All three strains were cultivated with 25 mM NaCl and ample bacterial food and then tested on an osmotic pressure gradient. The results showed that wild-type worms preferentially moved toward the cultivation osmotic pressure. The TRPA-1 knockout, RB1052, worms showed a lesser preference, but it was not significantly different from the wild type (P = 0.21; Table 1 and Fig. 2C). In contrast, the TRPV knockout, FG125, worms actually moved away from the side with the cultivation osmotic pressure. The osmotaxis index of FG125 strain was significantly different from the osmotaxis indices of both the wild-type and RB1052 worms (P < 0.001 and P < 0.01, respectively; Table 1 and Fig. 2C).
Discussion
In this study, we used several behavioral assays to examine the effects of TRPA-1 or TRPV channels on the behaviors of C. elegans, which are controlled by different neurocircuits. As expected, loss of TRPA-1 and TRPV channels completely abolished the behaviors related to thermosensation and osmosensation, respectively. Interestingly, loss of TRPV channels also greatly reduced associative learning regarding the temperature of food source. In contrast, loss of TRPA-1 channel did not have a statistically significant effect on the behaviors related to chemosensation, including both avoidance response to chemical repellent and NaCl osmotic pressure detection.
Thermotaxis in C. elegans is mainly determined by the AFD sensory neuron and AIY interneuron. 17 Some studies also reported possible roles of sensory neurons, such as AWC and ASI, and of interneurons, such as AIY, AIZ, and RIA. 18 Among these neurons, it is known that AWC expresses osm-9, 19 but whether or not these neurons express other TRPV channels is not clear. Therefore, one explanation for our results is that TRPA-1 and TRPV channels are coexpressed on the membrane of the neurons in the thermotaxis circuitry and that TRPV channels may modulate or even determine the functions of the TRPA-1 channel. Alternatively, TRPV channels may also have roles other than conducting ionic currents to modulate TRPA channel function. For example, the OCR-2 channel may have a direct nuclear function in neuronal cells. 20 Deficiency of ocr-2 reduced the expression of the diacetyl receptor ODR-10 in AWA neurons, and osm-9 mutant and ocr-1/ocr-2 double mutant completely abolished it. 21 Furthermore, C. elegans knockouts of osm-9 responded normally to cold shock, 14 suggesting the functional redundancy of TRPV channels, or different pathways to process cold shock or ambient cool temperature. Identification of the expression pattern of TRPA-1 and TRPV channels in the thermosensory neurons, together with more types of temperature related experiments, will further elucidate this mechanism.
The reversal movement to avoid chemical repellent is mediated by sensory neurons of the amphid, particularly the ASH and ASK neurons, 22 both of which express some TRPV channels (ASH: osm-9 and ocr-2;19,23 ASH: osm-9 19 ). In particular, ASH neuron was found to be involved in the nociceptive pathway to induce avoidance behavior. 24 OCR-2 and OSM-9 channels are colocalized in the sensory cilia and plasma membrane of the following four pairs of chemosensory neurons: ADF, AWA, ASH, and ADL. The TRPA-1 channel is also expressed in other sensory neurons, including ASH, OLQ, and IL1 neurons, and may be responsible for other behavioral responses, such as reversal movement and head withdrawal reflex in response to nose touch. 5 Therefore, TRPA-1 and TRPV channels are coexpressed in at least the ASH neuron related to chemosensation. However, in our experiments, the loss of TRPA-1 function did not significantly reduce the responses to chemorepellents or to NaCl osmosensation, suggesting that either TRPA-1 is not essential in the neurons controlling these responses or the system has other mechanisms to compensate the loss of TRPA-1 channel.
Author Contributions
Conceived and designed the experiments: JT, VN, FOL, and HL. Analyzed the data: JT, VN, and HL. Wrote the first draft of the article: HL. Contributed to the writing of the article: HL, JT, and FOL. All authors reviewed and approved of the final manuscript.