
Abstract
Synaptic plasticity critically depends on reciprocal interactions between neurons and glia. Among glial cells, microglia represent approximately 10% of the total brain cell population serve as the brain's resident macrophage, and help to modulate neural activity. Because of their special role in the brain's immune response, microglia are involved in the pathological progression of neurodegenerative disorders such as Alzheimer's disease (AD). However, microglia also are surveyors of the brain's health and continuously contact dendritic spines to regulate structural synaptic changes. This review summarizes our current understanding of neuronal-microglial signals that affect neural function at the synapse. Here, we examine the role of microglia in neuronal synapses in pathological brains and specifically focus on in vivo studies using 2-photon microscopy. Furthermore, because the role of microglia in AD progression is controversial, we outline the interaction between neurons and microglia in pathological conditions such as AD.
Introduction
Microglia reside in the central nervous system (CNS), where they are the predominant immune cells. First described nearly a century ago by Pio del Rio Hortega, 1 microglia constitute 20% of all glial cells and approximately 10% of all cells in the brain. 2 Microglia typically have a distinct ramified morphology and are distributed throughout the CNS in a nonoverlapping manner.2,3 During development, microglia descend from fetal macrophages. Fetal macrophages have a mesodermal origin, and, during embryogenesis and early postnatal development, they infiltrate the brain where they proliferate locally.4,5 These cells have a round morphology without processes and, therefore, are referred to as “amoeboid microglia.”6,7 Amoeboid microglia are highly mobile and proliferative phagocytic cells that play an important role in neuronal maturation. 8 After infiltrating the brain parenchyma, amoeboid microglia develop their typical ramified morphology and weakly express molecules associated with macrophage and phagocytic functions. Therefore, microglia in this developmental stage are traditionally defined as “resting microglia.”4,9–11 Unlike other bone marrow–derived macrophages, microglia are not generally replaced by bone marrow–derived monocytes except when the CNS is exposed to irradiation. 12 In spite of their name, resting microglia are highly dynamic and constantly survey their microenvironment by extending and retracting their processes at an average velocity of 1.47 mm/minute.13,14 These microglial processes physically contact either astrocytes or neurons, and, through their combined efforts, they can monitor the entire brain parenchyma within a few hours. Thus, microglia play a major role in developing neuronal synapses.8,14 In circumstances where CNS homeostasis is perturbed, such as infection, trauma, ischemia, or neurodegenerative disease, resting microglia change their morphology and become activated. 10 With these morphological changes, complex processes become simplified, and ramified microglia revert to the amoeboid shape. Once activated, microglia function as brain macrophages by increasing their phagocytic capacity, proliferating, acting as antigens for T-cells, and releasing factors and compounds, such as proinflammatory or anti-inflammatory cytokines and chemokines.7,9
Most of our knowledge about microglial function is based on in vitro studies that examined dissociated cells and brain slices. Although the in vitro approach provides valuable information about microglial function and behavior, this paradigm does reveal microglia behavior as it occurs in the intact CNS, especially cell-to-cell interactions. Furthermore, microglia readily activate and undergo morphological changes in in vitro preparations, 9 which mimics injury rather than the normal state. Therefore, in vivo studies using living animals are ideal for investigating microglia functions in the context of a healthy, intact brain. 15 In particular, technological advances including 2-photon laser scanning microscopy 16 allows real-time observation of microglial behavior in transgenic mice with microglia expressing green fluorescent protein (GFP).14,17 In this review, we highlight key findings from in vivo 2-photon microscopy studies regarding interactions between microglia and neurons in the healthy CNS and in Alzheimer's disease (AD).
Microglia and Synaptic Spines
Jung et al 17 genetically engineered transgenic mice to express enhanced green fluorescent protein (EGFP) under the Cx3crl locus (CX3CR1-GFP mice), which is the site where microglia are labeled with GFP. Using the 2-photon microscope and a thin-skulled transcranial approach, resting microglia in the adult cortex can be observed to be constantly surveying their microenvironment.13,14 Real-time imaging has revealed that microglia rapidly extend and retract their long cellular processes at an average rate of 1.47 μm/minute, 14 and the velocity of this process is greater than that for that of processes in other brain cells (ie, neurons at 0.2 μm/minute and astrocytes at > day). Considering that both resting and inactive ramified microglia are thought to exist in a healthy brain, the above observation was quite striking and prompted researchers to characterize the physical interactions between microglia and synaptic structures. As a result, double-transgenic mice expressing EGFP in both microglia and neurons (Iba-1-GFP/Thyl-GFP M line) have been established to visualize both microglia and neurons simultaneously for in vivo 2-photon imaging.18–20 Two-photon microscopy has revealed that microglia briefly (~5 minute) and repeatedly contact presynaptic terminals and dendritic spines in the somatosensory and visual cortices at a frequency of approximately 1 structure/hour. During these interactions, microglia appeared to be enlarged, and these morphological changes have been confirmed using 3-D reconstructions of serial section electron microscopy (SSEM) of the visual cortex of juvenile mice (4 weeks of age, which is the critical period for experience dependent synaptic plasticity) 21 in situ. 22 These structural changes appear to be activity-dependent, since the frequency of interaction between neurons and microglia was reduced by lowering the body temperature or injecting tetrodotoxin (TTX) into both retinae. 18 In addition, following cerebral ischemic injury, microglial contact with synaptic structures increased in duration, which in some cases resulted in synaptic destruction. 18 These data suggest that microglia can detect conditions at synapses and contribute to synaptic pruning during ischemic injury-induced remodeling.
Tremblay et al 22 described in detail the interactions between microglia and neurons. CX3CR1-GFP/Thyl-YFP mice with microglia expressing EGFP and neurons expressing yellow fluorescent protein (YFP) were used to visualize neurons and microglia simultaneously with 2-photon microscopy. In these experiments, experience-dependent changes in microglial interactions were observed during the critical period for the mouse visual system, which includes postnatal days (PND) 21 through 30. Changes in the number and sizes of dendritic spines in the visual cortex during the critical period are closely associated with the development of visual perception, such as direction selectivity and ocular dominance.23–25 Using 3-D reconstruction of SSEM, the appositions of microglia to dendritic spines, synaptic terminals, and synaptic clefts were confirmed in the visual cortex of juvenile mice during the critical period. Microglial processes preferentially interacted with smaller, developing dendritic spines. Real-time imaging with 2-photon microscopy revealed that the spines contacted by microglia were more frequently eliminated than noncontacted spines (24% vs 7%) during normal visual experience. 22 Following light deprivation, which was established by housing animals in complete darkness for 6 consecutive days during the critical period, the microglial processes became less motile and changed their preference from developing dendritic spines to larger ones that were deteriorating. However, these changes could be reversed with reexposure to light for 2 days. Collectively, these data demonstrate that microglia participate in synaptic pruning in response to sensory experience.
Microglial Synaptic Pruning
Abnormal synaptic pruning during critical periods of development leads to an increased synaptic density of immature dendritic spines. 26 This phenomenon is observed in various developmental or genetic disorders, such as fragile X syndrome in humans. 27 Therefore, elimination of unwanted synapses during development is critical for healthy neural circuits. Schafer et al showed that during developmental pruning in the mouse visual cortex, microglia engulf synapses of retinal ganglion cells in the lateral geniculate nucleus. 28 Using stimulated emission depletion (STDE) microscopy and immunogold electron microscopy techniques (immuno-EM), Paolicelli et al 29 demonstrated that microglia contribute to synaptic pruning during development. In their study, they confirmed that either PSD95 (postsynaptic component) or SNAP25 (presynaptic component) was colocalized with GFP-labeled microglia in the mouse hippocampus at PND 15, which indicated that microglia engulf synaptic components. Moreover, phagocytic inclusions that displayed ultrastructural synaptic features were observed within microglial cell bodies and processes; thus, microglial phagocytosis of dendritic spines likely persists during normal aging and throughout adulthood. 30
Immune molecules, fractalkine/fractalkine receptors (CX3CL1/CX3CR1), and classical complement cascades are proposed as potential mediators for microglia interactions at synapses, including synaptic pruning during development.28,29,31,32 CX3CR1 expressed specifically on the surface of microglia33,34 in the developing CNS appears to play an important functional role in synaptic pruning. In diseased states, such as AD, Parkinson's disease (PD), and amyotrophic lateral sclerosis (ALS), fractalkine (CX3CL1) and its receptor (CX3CR1) promote phagocytosis of apoptotic and injured neurons, affect the number of microglia, and recruit microglia to the injured site.17,35 Although the extent of colocalization of PSD95-immunoreactivity and GFP in CX3CR1-knockout (CX3CR1-KO) mice14,18 and littermate controls was similar, CX3CR1-knockout mice exhibited an increase in PSD95 immunoreactivity on PND 15, not PND28. 28 Paolicelli et al 28 showed a significant reduction in microglia density in the brains of CX3CR1 KO mice compared with littermate controls during postnatal weeks 2 to 3, and, in the period, the dendritic spine density of hippocampal CA1 pyramidal neurons were significantly enhanced. This indicated that microglial engulfment of synapses through CX3CR1 is a critical component of synaptic pruning in the developing brain. Due to inappropriate synaptic pruning by microglia, CX3CR1-KO mice showed an immature phenotype in excitatory synaptic transmission and exhibited enhanced long-term depression in the CA1 region of hippocampal slices. 28 These animals also showed a decreased latency of pentylenetetrazole (PTZ)-induced seizures, which indicated that proper synaptic pruning by microglia is required for neuronal circuit maturation during postnatal development. 28
The classical complement proteins C1q and C3 also might mediate microglial functions, including synaptic pruning during postnatal development. These complement proteins clear cellular debris or pathogens that have been tagged by immune cells for phagocytosis. 31 C1q and C3 are localized within immature synapses in the developing dorsolateral retinogeniculate nucleus (dLGN) of the thalamus, a region that is frequently examined for activity-dependent synaptic pruning during development.36,37 Microglia eliminate synapses that have been tagged in a complement receptor 3 (CR3)-dependent manner. 29 During the peak period of synaptic remodeling in the postnatal retinogeniculate system, a presynaptic element (ie, vGlut2) was observed within microglia by immuno-EM. Phagocytosis of synaptic inputs was decreased in mice devoid of C3 or CR3, which resulted in an increased synaptic density and deficits in segregation of eye-specific axonal projection to the retinogeniculate nucleus (LGN). These effects were maintained even in adulthood in CR3KO mice, which indicated a microglial contribution to synaptic pruning by using a complement system in the healthy brain. 31 In addition, CR3-depended engulfment of neurites was confirmed using in vitro coculture systems of neurons and microglia. 38 Taken together, these data suggest that microglia play a role in the structural plasticity in both the developing and healthy adult brain via the fractalkine/fractalkine receptor and the classical complement cascades.
Neuronal Activity and Microglia-Neuron Interactions
Findings from studies using in vitro culture assays indicate that microglia have receptors for neurotransmitters, neuropeptides, and neuromodulators, which suggests that microglia can respond to neural activity.7,39,40 Studies that have modulated electrical activity through ion channel blockers have yielded inconsistent results regarding microglia motility focusing on microglia. 41 A decrease in neural activity in the mouse cortex resulting from TTX application had no effect on the motility of microglial processes, whereas increased neural activity resulting from bicuculline (GABA [gamma-aminobutyric acid] A receptor blocker) increased motility. 14 Another study showed that a reduction in neural activity in the visual cortex induced by either intravitreal TTX injection or a body temperature reduction (to 32°C) decreased the frequency of microglia-synapse contact from approximately 1/h to 0.4/h. In these conditions, the speed of microglial motility was not affected. 18 In addition, microglial motility was decreased by GABA application in an ex vivo retinal explant model, whereas microglial motility increased with glutamate application. 42 These inconsistent outcomes might result from differences among brain regions, microglial heterogeneity among regions, or differences in pharmacological methods. Although there is some discrepancy among studies, microglial responses to neuronal activity has not yet been examined. Further research through in vivo studies will likely reveal the underlying mechanisms of microglial responses to neuronal activity, as well as identify the regional specificity of microglial motility and synaptic contacts.
Microglia can secrete signaling molecules, including neurotransmitters, cytokines, and extracellular matrix proteins, which can influence synaptic functions and activity (Fig. 1). 43 48 Therefore, microglia likely directly contribute to shaping synapses. A recent study reported the direct contribution of microglia activation on synaptic transmission in isolated hippocampal neurons. 45 In this study, microglia activation induced with lipopolysaccharide (LPS) application increased spontaneous excitatory postsynaptic currents and resulted in seizure-like bursting. Interestingly, this effect was achieved via the astrocyte purinergic receptor (P2Y1R) activated by ATP released from microglia. Astrocyte activation subsequently promoted glutamate release, which stimulated AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptors and increased the frequency of excitatory post-synaptic potentials through presynaptic metabotropic glutamate receptors (mGluR5). These results suggest that microglia can influence neural activity and the concept of the tripartite synapse (presynapse, spine, and astrocytes) should be modified to that of quadripartite synapses to better describe synaptic transmission. More importantly, these results provide insight into microglial contributions to synaptic dysfunction in neurological diseases, since aberrant microglial functions have been observed in diseases such as AD and PD.

Signaling crosstalk between neurons and microglia under physiological conditions. This is a simplified list of the neuron-microglia signaling pathway.
Interaction of Microglia and Neurons in AD
Whether such microglial contributions to synaptic pruning and maturation of neuronal circuits in the healthy CNS are compromised in the pathological brain is unclear. Pathological microglial functions have been implicated in several neurodegenerative diseases. 49 52 Among these neurodegenerative diseases, AD is one of the most extensively studied for microglial influences on amyloid plaques and synaptic plasticity. AD is the most common form of clinically recognized dementia in elderly people. First described by Alois Alzheimer in 1907, 53 AD is a fatal and progressive disease clinically characterized by memory impairment and cognitive decline. 54 Pathological hallmarks of AD are accumulation of senile plaques that are enriched with amyloid β (Aβ) peptide and neurofibrillary tangles that contain hyperphosphorylated microtubule-associated protein tau.55,56 Although accumulation of microglia in senile plaques has been observed in patients with AD and animal models of AD, 57 the exact microglial contributions to AD pathogenesis are still unknown. Many studies have focused on the relationship between plaque formation and microglial contributions. Until now, the role of microglia in AD progression is controversial. In early AD, microglial accumulation may delay disease progression by promoting clearance of Aβ before senile plaques form. However, as the disease progresses, proinflammatory cytokines produced in response to the amyloid plaques inhibit Aβ clearance, and then promote Aβ accumulation, which subsequently contributes to neuronal death.58,59 A number of in vivo 2-photon studies have examined microglial contributions to AD progression and especially to neuronal death.60,61 Fuhrmann et al 62 genetically crossed mice with AD and transgenic mice (3XTg, CX3CR1-GFP, and Ty1-YFP) to visualize microglia and neurons in cortical layer III with 2-photon imaging. Compared with 3XTg:CX3CR1+/-: Ty1-YFP mice, mice lacking the CX3CR1 receptor (3XTg:CX3CR1-/-: T y 1 - Y F P) showed greater microglia-mediated neuronal loss in addition to attenuated microglial migratory velocity. 62 These results indicate a critical role for the CX3CR1 chemokine receptor in microglia-mediated neuronal loss in AD transgenic mice. In contrast to the observation that CX3CR1 deficiency is beneficial for neuronal loss in 3XTg mice, CX3CR1 deficiency was found to be neither neuroprotective nor neurotoxic in CX3CR1-/-:CRND8 Tg mice. 63 This discrepancy might result from the different mutations in the 2 AD animal models. Whether the CX3CR1 molecular pathway of microglia is involved in preventing neuronal loss or is just recruited to the insult site is still debatable. Longitudinal studies with different AD animal models are necessary to understand the precise mechanism of neuronal-microglial communication in AD pathology.
Conclusion
In this review, we highlighted the key findings of recent studies regarding microglial contribution to neuronal processes in the healthy CNS and in AD pathology by focusing on in vivo studies. Resting microglia are highly dynamic and constantly survey the brain parenchyma by extending and retracting processes. In both the developing and adult nervous systems, microglia play an important role in brain immunity. In addition to having immune functions and clearing debris by phagocytosis, both resting and challenged microglia play an active role in neuronal circuit homeostasis (Fig. 2). Microglial interactions with synapses can affect the maturation of presynaptic and postsynaptic spines and influence synaptic transmission (Table 1). In vivo studies using 2-photon microscopy have revealed microglial influences on neuronal viability in pathological conditions such as AD. Microglia accumulation in senile plaques has been demonstrated in AD, but the exact role of microglia in AD pathogenesis remains unknown. Nonetheless, it seems plausible that elucidating the molecular mechanisms and functional significance of microglia-synapse interactions may offer new avenues for AD treatment.
Physiological roles of microglia in neuronal circuits. Schematic diagram indicating the physiological functions of microglia in neuronal circuits. Microglia play a role in maintaining neuronal circuits during development ( Overview of synapse-related microglia.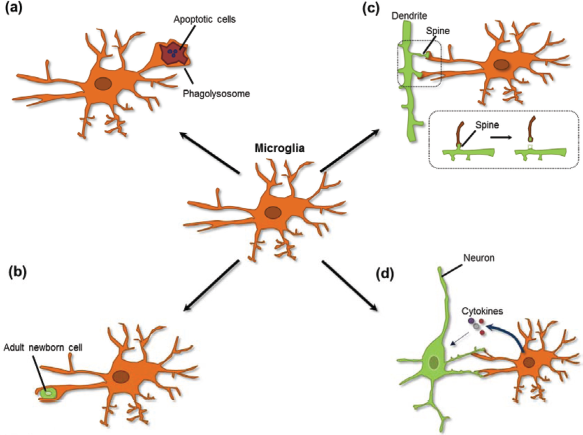

Author Contributions
Supervised the study: IM-J. Reviewed and edited the manuscript: IM-J. Reviewed and approved of the final manuscript: KHK and SMS. All authors reviewed and approved of the final manuscript.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.