
Abstract
De novo neurosteroidogenesis from cholesterol occurs in the brain of various avian species. However, the biosynthetic pathways leading to the formation of neurosteroids are still not completely elucidated. We have recently found that the avian brain produces 7α-hydroxypregnenolone, a novel bioactive neurosteroid that stimulates locomotor activity. Until recently, it was believed that neurosteroids are produced in neurons and glial cells in the central and peripheral nervous systems. However, our recent studies on birds have demonstrated that the pineal gland, an endocrine organ located close to the brain, is an important site of production of neurosteroids de novo from cholesterol. 7α-Hydroxypregnenolone is a major pineal neurosteroid that stimulates locomotor activity of juvenile birds, connecting light-induced gene expression with locomotion. The other major pineal neurosteroid allopregnanolone is involved in Purkinje cell survival during development. This paper highlights new aspects of neurosteroid synthesis and actions in birds.
Keywords
Introduction
Steroids supplied by peripheral steroidogenic glands regulate a variety of important brain functions during development, which persist into adulthood in vertebrates. Because steroids are lipid soluble, peripherally secreted steroid hormones can cross the blood-brain barrier and act on the brain through intracellular receptors that regulate the transcription of specific genes. Accordingly, the brain has been considered as a target site of peripheral steroid hormones in vertebrates. In contrast to this classical concept, studies conducted over the past two decades have demonstrated that the central and peripheral nervous systems have the capacity of synthesizing steroids de novo from cholesterol, the so-called neurosteroids (See reviews by Baulieu, 1 Tsutsui et al,2,3 Compagnone and Mellon, 4 Mellon and Vaudry, 5 Tsutsui et al,6,7 Tsutsui and Mellon, 8 and Do-Rego et al 9 ).
Baulieu and colleagues 10 18 originally demonstrated the formation of neurosteroids in the brain of mammals. It is now known that the brain of nonmammalian vertebrates also possesses several kinds of steroidogenic enzymes and produces a variety of neurosteroids (See reviews by Tsutsui et al,2,3 Mellon and Vaudry, 5 Tsutsui et al,6,7 Tsutsui and Mellon, 8 and Do-Rego et al 9 ). In birds, biosynthesis of neurosteroids has been reported in galliform bird species such as the Japanese quail Coturnix japonica2,6,19–26 and in passeriform bird species such as the zebra finch Taeniopygia guttata. 27 35 The formation of several neurosteroids from cholesterol is now also documented in various species of amphibians 36 47 and fish. 48 51 Therefore, de novo synthesis of neurosteroids from cholesterol in the brain appears to be conserved across vertebrate species (See reviews by Baulieu, 1 Tsutsui et al,2,3 Compagnone and Mellon, 4 Mellon and Vaudry, 5 Tsutsui et al,6,7 Tsutsui and Mellon, 8 and Do-Rego et al 9 ).
However, the biosynthetic pathways leading to the formation of neurosteroids in vertebrates are still not completely elucidated (See review by Tsutsui et al 7 ). In fact, Tsutsui and colleagues recently identified 7α-hydroxypregnenolone as a novel bioactive neurosteroid in the brain of the Japanese quail 52 and the Japanese red-bellied newt Cynops pyrrhogaster. 40 Importantly, 7α-hydroxypregnenolone acts on brain tissue as a novel neuronal modulator to stimulate locomotor activity of quail 52 and newts. 40 It was also found that cytochrome P450 7α-hydroxylase (cytochrome P4507α, gene name Cyp7b) catalyzes pregnenolone to produce 7α-hydroxypregnenolone in the brain of these species.52,53 It was further demonstrated that melatonin acts on cytochrome P4507α-expressing neurons to regulate 7α-hydroxypregnenolone synthesis, thus regulating diurnal locomotor activities in quail. 52
Until recently, it was believed that neurosteroids are produced only in neurons and glial cells in the central and peripheral nervous systems. Now there is evidence that in the juvenile chicken and quail, the pineal gland, an endocrine organ located close to the brain, actively produces a variety of neurosteroids de novo from cholesterol.54,55 Notably, 7α-hydroxypregnenolone and allopregnanolone (3α,5α-tetrahydroprogesterone, that is, 3α,5α-THP) are major neurosteroids secreted by the pineal gland.54,55 Importantly, the avian pineal gland produces 7α-hydroxypregnenolone that stimulates locomotor activity in light-dependent and circadian time-dependent manners. 54 On the other hand, allopregnanolone produced by the pineal gland prevents cell death of Purkinje cells in the cerebellum during development. 55
Based on new findings obtained by the studies on birds, this review highlights the advances in our understanding of the biosynthesis and biological actions of 7α-hydroxypregnenolone, a newly discovered bioactive neurosteroid, in the avian brain. Because the effect of 7α-hydroxypregnenolone on locomotion may be through neuromodulation, this review also describes recent findings in songbirds and quail, where neurosteroids have been implicated to have rapid neuromodulatory effects that influence song production and processing or sexual behavior of birds. Finally, this review describes what are currently known about the biosynthesis and biological actions of pineal 7α-hydroxypregnenolone and allopregnanolone in birds.
Classical Concept of Neurosteroidogenesis in the Avian Brain
Birds have served as excellent animal models for the investigation of neurosteroidogenesis. Tsutsui and colleagues analyzed neurosteroids formed from cholesterol using the Japanese quail and demonstrated that the brain of this bird possesses cytochrome P450 side-chain cleavage enzyme (P450scc, gene name Cyp11a), 3β-hydroxysteroid dehydrogenase/Δ
5
-Δ
4
-isomerase (3β-HSD, gene name Hsd 3b), 5β-reductase (gene name Srd5b), cytochrome P450 17α-hydroxylase/c17,20-lyase (P45017α, yase, gene name Cyp17), 17β-hydroxysteroid dehydrogenase (17β-HSD, gene name Hsd17b), and so on, and produces pregnenolone, progesterone, epipregnanolone (3β,5β-tetrahydroprogesterone, that is, 3β,5β-THP), androstenedione, testosterone, and estradiol-17β from cholesterol (Fig. 1) 2,6,19–26 The expression and activity of cytochrome P450 aromatase (P450arom, gene name Cyp19), which converts testosterone into estradiol-17β, have also been demonstrated in the quail brain (Fig. 1).
56
67
Schlinger and colleagues independently performed similar studies to demonstrate neurosteroidogenesis in the brain of zebra finches.
27
35
The formation and metabolism of neurosteroids from cholesterol is now established in the brain of birds.
Classical biosynthetic pathways for neurosteroids in the avian brain. The arrows indicate the biosynthetic pathways of neurosteroids identified previously in the quail brain. De novo neurosteroidogenesis in the brain from cholesterol appears to be a conserved property across vertebrates. P450scc, cytochrome P450 side-chain cleavage enzyme; 3β-HSD, 3β-hydroxysteroid dehydrogenase/Δ
5
-Δ
4
-isomerase; P45017α, lyase, cytochrome P450 17α-hydroxylase/c17,20-lyase; 17β-HSD, 17β-hydroxysteroid dehydrogenase; P450arom, cytochrome P450 aromatase. See the text for details.
As summarized in Figure 1, it appears that the avian brain possesses a variety of steroidogenic enzymes and produces pregnenolone, progesterone, epipregnanolone, androstenedione, testosterone, and estradiol-17β from cholesterol. The discovery of these neurosteroids in the avian brain has expanded our knowledge of the sources of active steroidal molecules, the time-course of their actions in the brain, and the kinds of brain functions in which neurosteroids have significant functions (See review by Tsutsui 68 ). Studies of avian neurosteroids are currently of great interest to many researchers.
Discovery of 7α-Hydroxypregnenolone, a Novel Bioactive Neurosteroid, in the Avian Brain and Its Biological Action on Locomotion
Identification of 7α-hydroxypregnenolone
Recently, 7α- and 7β-hydroxypregnenolone have been discovered in the avian brain as novel pregnenolone metabolites (Fig. 2).
52
Subsequently, it has been demonstrated that 7α-hydroxypregnenolone is converted from pregnenolone by cytochrome P4507α (Fig. 2).
52
Identification of 7 α- and 7β-hydroxypregnenolone in the avian brain and stimulatory action of 7α-hydroxypregnenolone on locomotor activity in birds. (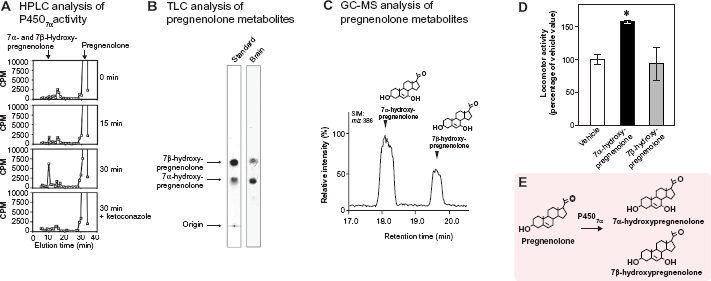
Based on a preliminary finding that the quail brain actively produces unknown neurosteroids from pregnenolone, Tsutsui and colleagues sought to identify these neurosteroids from the adult quail brain by using biochemical techniques including high-performance liquid chromatography (HPLC), thin-layer chromatography (TLC) and gas chromatography-mass spectrometry (GC-MS) analyses. 52 Quail brain homogenates were incubated with tritiated pregnenolone and radioactive metabolites were analyzed by reversed-phase HPLC. Several nonradioactive steroids were used as reference standards for HPLC analysis, and 7α-hydroxypregnenolone and its stereoisomer, 7β-hydroxypregnenolone, exhibited the same retention time of the radioactive peak (Fig. 2A). 52 The HPLC peak fraction was collected and subjected to TLC to separate the isomers. Quail brain homogenates produced two metabolites from 3 H-pregnenolone corresponding to the positions of the 7α- and 7β-hydroxypregnenolone standards by TLC analysis (Fig. 2B). 52 The metabolites of pregnenolone were further analyzed by GC-MS. Based on GC-selected ion monitoring (SIM) analysis (m/z 386), the metabolites had retention times that were identical to those of 7α-hydroxypregnenolone and 7β-hydroxypregnenolone, respectively (Fig. 2C). 52
Identification of cytochrome P4507α and 7α-hydroxypregnenolone formation
7α-Hydroxypregnenolone is synthesized from pregnenolone through the enzymatic activity of cytochrome P4507α (Fig. 2E). To demonstrate that 7α-hydroxypregnenolone is synthesized in the brain, it is necessary to show that the brain expresses cytochrome P4507α. A 2,341-bp full-length cDNA encoding a putative cytochrome P4507α was identified from the quail brain. 52 The enzymatic activity of this putative quail cytochrome P4507α was demonstrated in the homogenates of COS-7 cells transfected with the putative quail cytochrome P4507α cDNA. 52 Combination of HPLC and GC-MS analyses revealed that the homogenate converted pregnenolone into 7α-hydroxypregnenolone. Both 7α- and 7β-hydroxypregnenolone are clearly present in the quail brain, although it is still unclear whether cytochrome P4507α can also convert pregnenolone into 7β-hydroxypregnenolone (Fig. 2). 52 The production of 7α-hydroxypregnenolone in the brain may be a conserved property of vertebrates because this neurosteroid has been identified in the brain of newts 40 and mammals. 69 72 Recently, a cDNA encoding cytochrome P4507α was identified in the newt brain. 53 The homogenate of COS-7 cells transfected with the newt cytochrome P4507α cDNA indeed converted pregnenolone into 7α-hydroxypregnenolone. 53
The biosynthesis and concentrations of 7α- and 7β-hydroxypregnenolone in different brain regions of the quail of both sexes were compared by HPLC and GC-MS analyses. 52 The two neurosteroids were found at the highest concentration in the diencephalon, and their concentrations were very low in other brain regions. 52 The biosynthetic activities and concentrations of 7α- and 7β-hydroxypregnenolone in the diencephalon were found to be much higher in males than females. 52 Such a sexual dimorphism of cytochrome P4507α only occurs in the diencephalon. 52 Similarly, there are sex differences in 3β-HSD and cytochrome P450arom in the avian brain.33,34,56
Biological action of 7α-hydroxypregnenolone on locomotor activity
It is well known in birds 52 as well as in other vertebrates73,74 that locomotor activity of males is higher than that of females. We found that there were clear sex differences in the synthesis and concentration of diencephalic 7α- and 7β-hydroxypregnenolone. 52 It may be that these neurosteroids play a role in the control of locomotor activity of males. Because the male quail displays a robust locomotor activity rhythm when held under typical light/dark lighting schemes,75,76 this bird serves as an excellent animal model to demonstrate the biological action of 7α- and 7β-hydroxypregnenolone. Both of the neurosteroids were administered intracerebroventricularly (ICV) to the male quail during night, when the activity is low, to examine whether they could affect locomotor activity. 52 Thirty minutes after administration of 7α-hydroxypregnenolone, locomotor activity was measured by using an implantable telemetry system. 52 A stimulatory effect of 7α-hydroxypregnenolone was observed in male quail (Fig. 2D). 52 In contrast, 7β-hydroxypregnenolone did not influence the locomotor activity (Fig. 2D). 52 It thus appears that 7α-hydroxypregnenolone acts as a novel bioactive neurosteroid to stimulate locomotor activity in male quail (See reviews by Tsutsui et al 77 81 ). A similar stimulatory effect of 7α-hydroxypregnenolone on locomotor activity has been shown in male newts.40,82
From sex differences in 7α-hydroxypregnenolone synthesis, concentration, and locomotor activity in quail, 52 it is considered that this neurosteroid plays an essential role in the control of locomotor activity in males. 52 Consistent with this notion, it has been shown that the cytochrome P450 inhibitor ketoconazole decreases locomotor activity in male quail. 52 Unlike males, 7α-hydroxypregnenolone administration does not affect locomotor activity in females, 52 suggesting that the receptor for 7α-hydroxypregnenolone is not present or inactive in the female.
Mode of action of 7α-hydroxypregnenolone
It is important to clarify the mode of action of 7α-hydroxypregnenolone on locomotor activity in birds and other vertebrates. Tsutsui and colleagues have first indicated that 7α-hydroxypregnenolone acts as a neuronal modulator to stimulate locomotor activity of male newts through the dopaminergic system. 40 7α-Hydroxypregnenolone increased the concentration of dopamine in the male newt brain, especially in the rostral brain region including the striatum, which is known to be involved in the regulation of locomotor behavior. 40 In addition, 7α-hydroxypregnenolone increased dopamine release from cultured male brain in vitro. 40 The effect of 7α-hydroxypregnenolone on locomotion was abolished by administration of haloperidol or sulpiride, two dopamine D2 receptor antagonists. 40
In the male quail brain, the expression of Cyp7b mRNA was localized in several diencephalic regions, such as the nucleus preopticus medialis (POM), the nucleus paraventricularis magnocellularis (PVN), the nucleus ventromedialis hypothalami (VMN), the nucleus dorsolateralis anterior thalami (DLA), and the nucleus lateralis anterior thalami (LA).
52
Dopaminergic neurons that are located in the mesencephalic region, including the ventral tegmental area (VTA) and the substantia nigra (SN), project to the telencephalon, in particular in the striatum in birds.83,84 Importantly, the telencephalic region is enriched with dopamine D1 and D2 receptors in birds.85,86 Accordingly, 7α-hydroxypregnenolone actively synthesized in the diencephalon may act on dopamine neurons localized in the VTA and SN to stimulate dopamine release from their termini in the striatum and increase locomotor activity in male quail as in male newts (See reviews by Tsutsui et al
77
81
and Haraguchi et al
82
) (See Fig. 3A).
Mode of action of 7α-hydroxypregnenolone on locomotor activity and melatonin action on 7α-hydroxypregnenolone synthesis in quail. (
The acute stimulatory action of 7α-hydroxypregnenolone on locomotor activity strongly suggests that this neurosteroid acts through a nongenomic rather than a genomic mechanism in quail. 52 It has been reported in the rat that progesterone metabolite allopregnanolone modulates locomotion 87 and dopamine release88,89 via a nongenomic pathway. It is hypothesized that the neuromodulatory action of allopregnanolone is mediated through γ-aminobutyric acid type A (GABAA) receptors, since allopregnanolone is a potent allosteric modulator of GABAA receptors90,91 and dopaminergic neurons are regulated by GABAergic transmission. 92 Similarly, pregnenolone can also act via nongenomic mechanisms by binding to GABAA and N-methyl-D-aspartate (NMDA) receptors to enhance neuronal excitability.91,93 Whether the acute actions of 7α-hydroxypregnenolone on dopamine release and locomotor activity in quail are also mediated through GABAA and/or NMDA receptors, or through an unknown membrane receptor, remains to be determined.
Diurnal changes in 7α-hydroxypregnenolone synthesis and its regulatory mechanisms
To clarify the functional significance of 7α-hydroxypregnenolone in the regulation of locomotor activity, diurnal changes in both locomotor activity and diencephalic 7α-hydroxypregnenolone concentrations were analyzed in the male quail exposed to a daily photoperiod of 16 h/8 h light/dark cycles (lights on at 07:00 am, off at 11:00 pm). Locomotor activity of males was much higher than that of females from the time of lights on until noon but decreased to female levels thereafter. 52 These changes in locomotor activity in males were directly correlated with 7α-hydroxypregnenolone concentrations in the diencephalon, the maximum value occurring at 11:00 am when locomotor activity was high. 52 Furthermore, administration of ketoconazole suppressed locomotor activity at 11:00 am. 52 Thus, the increase in diencephalic 7α-hydroxypregnenolone may account for the higher locomotor activity in males. As mentioned above, the lower level of 7α-hydroxypregnenolone synthesis and concentration in the female diencephalon suggests that this neurosteroid may not be involved in the control of locomotor activity in females.
Melatonin may regulate the biosynthesis of 7α-hydroxypregnenolone in the diencephalon and thereby influence locomotor activity because melatonin is known to be involved in the regulation of locomotor activity in birds. 94 100 A series of experiments were thus carried out to investigate the possible involvement of melatonin in the regulation of diurnal changes in 7α-hydroxypregnenolone production in male quail. 52 Concomitant pinealectomy (Px) and orbital enucleation (Ex) provoked a marked increase in the production and concentration of 7α-hydroxypregnenolone and stimulated the expression of Cyp7b mRNA in the quail diencephalon. 52 Reciprocally, melatonin administration to Px/Ex quail decreased the production and concentration of 7α-hydroxypregnenolone and inhibited the expression of Cyp7b mRNA in the diencephalon. 52 The inhibitory effect of melatonin on 7α-hydroxypregnenolone synthesis was abrogated by luzindole, a melatonin receptor antagonist. 52 It thus appears that melatonin secreted by the pineal gland and eyes may act as an inhibitory factor of 7α-hydroxypregnenolone synthesis in the quail brain (Fig. 3B). This mechanism may account for the results of earlier studies indicating that melatonin treatment reduces locomotor activity in quail,99,101 sparrows, and owls. 99
It is well established that the nocturnal secretion of melatonin depends on the duration of the dark period, 102 the onset of melatonin secretion occurring soon after the onset of darkness. 103 Therefore, the increase in 7α-hydroxypregnenolone production observed in the brain of male quail during the light period can likely be accounted for by the decrease in endogenous melatonin secretion (Fig. 3B). Since 7α-hydroxypregnenolone stimulates locomotor activity in male quail, this neurosteroid may play a crucial role in diurnal changes in locomotor activity through the action of melatonin.
In birds and other vertebrates in general, there is a circadian rhythm in locomotor activity controlled by daily rhythm of melatonin secretion.94–100,104 However, until recently, the neuroendocrine mechanisms mediating this behavioral action of melatonin remained totally unknown. The discovery of the function of 7α-hydroxypregnenolone in mediating the action of melatonin on diurnal locomotor rhythmicity is an important step in understanding these mechanisms. 52 A similar mechanism may underlie the regulation of diurnal locomotor rhythms in other vertebrates, because 7α-hydroxypregnenolone is also produced in the brain of newts 40 and mammals 69 72 (See reviews by Tsutsui et al 77 81 ).
Discovery of Neuromodulatory Effects of Neuroestrogens in the Avian Brain
As described above, the brain of quail and other birds possesses cytochrome P450arom, which converts testosterone into estradiol.56–62,105 Cytochrome P450arom and estrogen receptors are both expressed in several brain regions including the preoptic area that is involved in the control of reproductive behaviors in birds.56–62,105 We detected, biochemically, the formation of estradiol from progesterone in the quail diencephalon including the preoptic area. 20
There is evidence for activation of territorial behavior by neurosteroids in the song sparrow Melospiza melodia.106,107 It is known that territorial behavior of this species is expressed in the nonbreeding season, although circulating testosterone levels are low. 108 Because the brain of zebra finch expresses 3β-HSD and cytochrome P450arom,34,35,67 these steroidogenic enzymes may produce estrogens from dehydroepiandrosterone originated from the peripheral gland during the nonbreeding season. Because the brain of zebra finches also expresses cytochrome P450scc and cytochrome P45017α,lyase, 28 31 dehydroepiandrosterone may also be produced de novo from cholesterol in the brain of these birds. More research is needed to evaluate the function of neurosteroids produced in the brain from cholesterol de novo and the role of central metabolism of steroids originally coming from the periphery. As in zebra finches, song sparrows expressed 3β-HSD and cytochrome P450arom in the brain.67,109,110 Cytochrome P450arom is elevated in the non-breeding season in the brain. 109 3β-HSD is expressed and active in the song sparrow brain.67,110 3β-HSD is also elevated during the non-breeding season.
There are several reports showing changes in neurosteroid formation in relation to social interactions. A recent study showed that within the caudomedial nidopallium marked changes in estradiol occurred when males were exposed to females or to conspecific zebra finch song. 111 Estrogens produced in the local brain region are thought to rapidly strengthen auditory encoding and guide song preference in a songbird. 112 Changes in estradiol formation were reduced by exposure to fadrozole, an inhibitor of cytochrome P450arom, or to glutamate as in quail hypothalamus. 66 These findings suggest rapid control of cytochrome P450arom activity by glutamatergic inputs. 66 In quail hypothalamic explants, cytochrome P450arom undergoes Ca2+-dependent phosphorylation that reduces cytochrome P450arom activity within minutes.63,65 Treatments of these explants with K+ or with glutamate receptor agonists produce a similar rapid inhibition of cytochrome P450arom activity. 63 These results suggest that voltage-gated Ca2+ channels serves as a key regulatory signal for rapid estrogen production.
Synaptic estrogen formation in the brain is becoming clear in songbirds and other birds113,114 as in mammals. 115 Compartmentalization of cytochrome P450arom within presynaptic boutons is considered to be crucial to provide sex- and song-specific estrogenic signals in the songbird brain.114,116
Biosynthesis of Neurosteroids in the Pineal Gland and Biological Actions of Pineal Neurosteroids in Birds
Neurosteroidogenesis in the pineal gland
Until recently, it was generally believed that neurosteroids are produced only in the central and peripheral nervous systems. However, our recent studies in chickens 54 and quail 55 have demonstrated that the pineal gland, an endocrine organ located close to the brain, actively synthesizes neurosteroids de novo from cholesterol (Fig. 4). In fact, the steroidogenic acute regulatory protein (StAR, gene name StAR) and cytochrome P450scc were both expressed in the pineal gland of juvenile chickens 54 and juvenile quail (Fig. 4). 55 Immunohistochemistry with cytochrome P450scc antibodies showed intense staining in cells forming follicular structures in the quail pineal gland. 55 Incubation of the pineal glands of quail chicks with tritiated cholesterol led to the formation of a radioactive metabolite that exhibited the same retention time as pregnenolone by HPLC analysis. 55 The occurrence of pregnenolone in the pineal gland was also demonstrated by GC-MS analysis. 55
Subsequently, the expressions of several key steroidogenic enzymes, including cytochrome P4507α, 3α-HSD, 3β-HSD, 5α-reductase, 5β-reductase, cytochrome P45017α,lyase, 17β-HSD, and cytochrome P450arom have been demonstrated in the pineal gland of both juvenile chickens and juvenile quail (Fig. 4).54,55 To clarify the biosynthetic pathways of neurosteroids in the pineal gland, biochemical studies combined with HPLC and GC-MS analyses were further conducted. Pineal gland homogenates from quail chicks were incubated with tritiated pregnenolone and subsequent reversed-phase HPLC analysis detected the formation of 7α- and/or 7β-hydroxypregnenolone (Fig. 4).
55
In addition, progesterone, allopregnanolone and/or epipregnanolone, androstenedione, testosterone, 5α- and/or 5β-dihydrotestosterone, and estradiol-17β were produced from the precursor pregnenolone (Fig. 4).
55
Although isomers such as 7α-and 7β-hydroxypregnenolone; allopregnanolone and epipregnanolone; and 5α- and 5β-dihydrotestosterone were not separated from each other by HPLC analysis, the formation of these neurosteroids in the pineal gland was demonstrated by GC-MS analysis.
55
Derivatives of synthetic 7α- and 7β-hydroxypregnenolone, progesterone, allopregnanolone, epipregnanolone, androstenedione, testosterone, 5α- and 5β-dihydrotestosterone, estradiol-17β, and the purified nonradioactive steroids produced by the pineal gland were applied to GC-SIM analysis, which showed the same mass spectral characteristics: m/z 386 for 7α- and 7β-hydroxypregnenolone, m/z 510 for progesterone, m/z 514 for allopregnanolone and epipregnanolone, m/z 482 for androstenedione, m/z 680 for testosterone, m/z 486 for 5α- and 5β-dihydrotestosterone, and m/z 664 for estradiol-17β.
55
Unlike HPLC analysis, GC-MS analysis was capable of separating several pairs of isomers: 7α- and 7β-hydroxypregnenolone; allopregnanolone and epipregnanolone; and 5α-and 5β-dihydrotestosterone.
55
As summarized in Figure 4, the neurosteroids produced in the pineal gland were thus identified as 7α- and 7β-hydroxypregnenolone, progesterone, allopregnanolone, epipregnanolone, androstenedione, testosterone, 5α- and 5β-dihydrotestosterone, and estradiol-17β.
55
These data provide the first evidence for de novo neurosteroidogenesis in the pineal gland in any vertebrate class.
Identified biosynthetic pathways for neurosteroids in the avian pineal gland. The arrows indicate the biosynthetic pathways of neurosteroids identifed in the pineal glands of juvenile quail. The pineal gland actively produces a variety of neurosteroids de novo from cholesterol. 7α-Hydroxypregnenolone and allopregnanolone are major products secreted by the pineal gland. P450scc, cytochrome P450 side-chain cleavage enzyme; P4507α cytochrome P450 7α-hydroxylase; 3β-HSD, 3β-hydroxysteroid dehydrogenase/Δ
5
-Δ
4
-isomerase; 3α-HSD, 3α-hydroxysteroid dehydrogenase/Δ
5
-Δ
4
-isomerase; P45017α, lyase, cytochrome P450 17α-hydroxylase/c17,20-lyase; 17β-HSD, 17β-hydroxysteroid dehydrogenase; P450arom, cytochrome P450 aromatase. See Haraguchi et al
55
and the text for details.
Identification of major neurosteroids synthesized in the pineal gland
To identify major neurosteroids synthesized in the pineal gland, the pineal glands from quail chicks were cultured in medium 199 with tritiated pregnenolone. HPLC analysis revealed that pregnenolone was converted primarily into 7α- and/or 7β-hydroxypregnenolone and allopregnanolone and/or epipregnanolone in the pineal gland. 55 HPLC analysis and real-time PCR in the pineal gland revealed that the synthesis of 7α- and/or 7β-hydroxypregnenolone and the expression of Cyp7b mRNA occur in both sexes of adult and juvenile quail, but they are significantly higher in juveniles than in adults. 55 Allopregnanolone and/or epipregnanolone synthesis and Srd5a mRNA expression were also higher in juveniles than in adults of both sexes. 55 The synthesis of 7α- and/or 7β-hydroxypregnenolone and the expression of Cyp7b mRNA were higher in the pineal gland than in the cerebellum and diencephalon. 55 Allopregnanolone and/or epipregnanolone synthesis and Srd5a mRNA expression were also higher in the pineal gland than in the cerebellum and diencephalon. 55
The pineal glands of quail chicks were cultured and the release of major neurosteroids was analyzed by GC-MS. Significant amounts of 7α-hydroxy-pregnenolone and allopregnanolone were found to be released from the pineal gland into the culture medium unlike 7β-hydroxypregnenolone and epipregnanolone (Fig. 4). 55 In sum, 7α-hydroxypregnenolone and allopregnanolone appear to be the major neurosteroids secreted from the pineal gland (Fig. 4).
Light-dependent synthesis of pineal 7α-hydroxypregnenolone and its biological action on locomotion
The original finding that the chicken pineal gland actively produces 7α-hydroxypregnenolone came from the analysis of light-dependent regulation of the circadian clock. 54 The circadian clock is the internal time-measuring system that controls daily rhythms of physiology and behavior even in the absence of external time cues. The phase of the circadian clock is adjusted by environmental stimulus such as light and food in a time-of-day-dependent manner (See review by Hirota and Fukada 117 ). For example, a light pulse given at early night and late night induced phase delay and advance, respectively, while the one at subjective daytime caused no significant phase shift. Such a phase-dependent light response of the circadian clock is conserved across species, but its mechanism still remains to be solved. The chick pineal gland is one of the best organs to work on this issue, because it expresses intrinsic photoreceptive opsins, such as pinopsin, 118 which confer light-sensitivity on the pineal circadian clock governing rhythmic production of melatonin (See review by Fukada and Okano 119 ).
To approach the molecular mechanism of the light-dependent phase-shift of the circadian clock, genes induced by a light pulse at different times of a day in the chicken pineal gland were searched by GeneChip analysis by comparing dark-reared juvenile chicks with those exposed to light at various times of the day: daytime, early night, or late night. This comprehensive transcriptome analysis revealed that a light pulse at early night induced a number of genes involved in cholesterol biosynthesis that are the targets of a transcription factor, sterol regulatory element- binding protein (SREBP). 54 In addition to the target gene expression, the light pulse at early night also stimulated the formation of the active form of SREBP transcription factor. Noticeably, the light response of SREBP-target genes was parallel to that of E4bp4. E4bp4 encodes a transcription factor that represses a core clock gene Per2, hence, associated with the phase-delay of the chick pineal clock (Fig. 5).120,121 E4bp4 turned out to be a new target gene of SREBP, revealing a new role of SREBP in the photic input pathway of the circadian clock. 54
The photic induction of a series of genes involved in cholesterol biosynthesis suggested a possible production of cholesterol (and its derivatives) in the pineal gland for physiological response to the light. The analysis of neurosteroidogenesis eventually revealed that the chick pineal gland actively produces and secretes 7α-hydroxypregnenolone (Fig. 5).
54
In accordance with the transcriptional changes in response to the light pulse, 7α-hydroxypregnenolone production was stimulated at a specific time of the day, that is, it was activated by a light pulse given at early night but not at late night and daytime.
54
Furthermore, the locomotor activity of dark-reared juvenile chicks was stimulated by light exposure more strongly at early night than at late night and daytime.
54
Intriguingly, the light-dependent stimulation of the locomotor activity at early night is reduced by Px.
54
Collectively, the pineal production of 7α-hydroxypregnenolone is stimulated by light in a time-of-day-dependent manner under the control of the circadian clock, and these unexpected properties may be essential for regulation of locomotor activity (Fig. 5).
Light-stimulated production of pineal cholesterol biosynthetic genes that is associated with light-stimulated production of 7α- hydroxypregnenolone in chickens. Light induction of a full set of genes governing cholesterol biosynthesis is mediated by light activation of SREBP, sterol regulatory element-binding protein, which in parallel activates transcription of E4bp4.
54
E4BP4 is a transcription factor that regulates the circadian clock through repression of clock gene Per2.120,121 SREBP-mediated induction of a series of cholesterol biosynthetic genes would up-regulate cholesterol synthesis and lead to stimulation of 7α-hydroxypregnenolone, a stimulator of locomotion. See Hatori et al
54
and the text for details.
Biological action of pineal allopregnanolone in Purkinje cell survival
Because the two major pineal neurosteroids, 7α-hydroxypregnenolone and allopregnanolone, are abundantly released from the pineal gland of juvenile birds, 55 not only pineal 7α-hydroxypregnenolone but also pineal allopregnanolone may play important roles in the brain of birds during development.
In birds, the pineal gland is located near the cerebellum (Fig. 6A). The cerebellar cortex has been used as an excellent model to study synaptic formation and transmission of neural networks because it forms relatively simple neuronal networks as compared with those of other brain regions. The Purkinje cell is a principal cerebellar neuron that integrates the process of memory and learning. It is known that in birds and mammals, Px induces cell loss in the brain including Purkinje cells during development.122,123 This observation suggests that allopregnanolone and/or 7α-hydroxypregnenolone secreted by the pineal gland may be involved in Purkinje cell survival during development. To test this hypothesis, Haraguchi et al
55
conducted a series of experiments in the male juvenile quail. Px decreased the concentration of allopregnanolone in the cerebellum and induced apoptosis of Purkinje cells, whereas administration of allopregnanolone to Px quail chicks increased the concentration of allopregnanolone in the cerebellum and prevented apoptosis of Purkinje cells.
55
In contrast to allopregnanolone, administration of 7α-hydroxypregnenolone to Px quail chicks did not rescue Purkinje cell death.
55
Haraguchi et al
55
further indicated that pineal allopregnanolone reaches the cerebellar Purkinje cells by diffusion as shown by injection of
3
H-allopregnanolone close to the pineal lumen (Fig. 6B). Thus, allopregnanolone secreted by the pineal gland is considered to be an important factor for Purkinje cell survival during development (Fig. 6B). Although 7α-hydroxypregnenolone did not facilitate Purkinje cell survival, this neurosteroid is involved in the regulation of locomotion in birds52,54 as mentioned above.
Neuroprotective action of pineal allopregnanolone on Purkinje cell survival during cerebellar development in quail. (
It is well known that caspase-3 plays an important role in Purkinje cell death in vertebrates.124,125 Caspase-3 is a crucial mediator of apoptosis 124 in vertebrates including birds.125,126 Importantly, Px increased the number of Purkinje cells that expressed active caspase-3, a key protease in apoptotic pathway, in quail chicks and administration of allopregnanolone to Px quail chicks decreased the number of Purkinje cells expressing active caspase-3. 55 These findings indicate that the neuroprotective effect of pineal allopregnanolone on Purkinje cells is associated with the decrease in caspase-3 activity during development. Accordingly, pineal allopregnanolone exerts antiapoptotic effects in Purkinje cells by suppressing the activity of caspase-3 during development (Fig. 6B). This is a new function of the pineal gland for the prevention of Purkinje cell death in the developing cerebellum.
It was generally accepted that the pineal gland transduces photoperiodic changes to the neuroendocrine system by executing rhythmic secretion of melatonin. However, the formation of neurosteroids in the pineal gland was, until recently, unknown in vertebrates. Our recent studies provide new evidence that the pineal gland is a major neurosteroidogenic organ and produces allopregnanolone far more abundantly than other brain regions. Importantly, pineal allopregnanolone acts on cerebellar Purkinje cells to prevent their programmed cell death during development. This is a paradigm shift of neurosteroid formation and action by the discovery of pineal allopregnanolone that facilitates neuronal survival in the cerebellum, because it was generally believed that neurosteroids are produced only in neurons and glial cells in the brain and other nervous systems.
Conclusions
Studies conducted over the past two decades have demonstrated that the brain of birds has the capacity of synthesizing various neurosteroids de novo from cholesterol. It appears, however, that the biosynthetic pathways leading to the formation of neurosteroids in the avian brain are still incompletely elucidated. 7α-Hydroxypregnenolone, a newly discovered neurosteroid produced by cytochrome P4507α in the avian brain, acts as an important neuromodulator to increase locomotor activity. The stimulatory action of 7α-hydroxypregnenolone may be mediated by the dopaminergic system. Melatonin acts on neurons expressing cytochrome P4507α to regulate 7α-hydroxypregnenolone synthesis, thus inducing diurnal locomotor changes. In this way, 7α-hydroxypregnenolone-producing neurons may play a pivotal role in the integration of circadian information that affects locomotor activity in birds. On the other hand, until recently, it was generally believed that neurosteroids are produced in neurons and glial cells in the brain and other nervous systems. However, there is now evidence that, in the juvenile chicken and quail, the pineal gland, an endocrine organ located close to the brain, actively produces a variety of neurosteroids de novo from cholesterol. 7α-Hydroxypregnenolone is a major pineal neurosteroid that stimulates locomotor activity of juvenile birds, connecting light-induced gene expression with locomotion. The other major pineal neurosteroid allopregnanolone prevents cell death of Purkinje cells by suppressing the activity of caspase-3 during cerebellar development. Interaction of brain and pineal neurosteroids in the regulation of brain functions deserve further investigations in birds.
Author Contributions
Conceived and designed the experiments: KT, YF. Analyzed the data: KT, SH, KI, HM, TU, MH, TH, YF. Wrote the first draft of the manuscript: KT, YF. Contributed to the writing of the manuscript: KT, SH, KI, HM, TU, MH, TH, YF. Agree with manuscript results and conclusions: KT, SH, KI, HM, TU, MH, TH, YF. Jointly developed the structure and arguments for the paper: KT, SH, KI, HM, TU, MH, TH, YF. Made critical revisions and approved final version: KT, SH, KI, HM, TU, MH, TH, YF. All authors reviewed and approved of the final manuscript.
Funding
This work was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Science and Culture, Japan (22132004 and 22227002 to KT and 24227001 to YF).
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.
Footnotes
Acknowledgments
We are grateful to the following collaborators, Masahiro Matsunaga, Saori Suzuki, Sakurako Hara, Masayuki Kusaka, Yuko Suzuki and Hubert Vaudry.