
Abstract
Neuroinflammation is found both in the brain of humans suffering from Parkinson's disease and in animal models of disease. It is suggested to be involved in the pathogenesis of the disease. In the present study, in order to study the effects of antioxidants on neuroinflammation, microglial phenotypes were evaluated in rats fed with diets containing bilberries, blueberries, or crowberries at 1 and 4 weeks following striatal injection of 6-hydroxydopamine. The dopamine innervation was visualized using antibodies raised against tyrosine hydroxlase (TH) in the striatum and in the globus pallidus. One week post-lesion, the expression of Iba1-positive cells, a general microglial marker, was significantly increased in the striatum of all animals fed with antioxidant-enriched diets compared to control-diet fed animals, while the diameter of the TH-negative zone was similar in all animals. At four weeks post-lesion, the Iba1-positive microglia was significantly reduced in animals fed with antioxidant-enriched diets. The diameter of the TH-negative zone was significantly reduced in animals fed bilberry and crowberry. The expression and distribution of ED1-positive cells was similar to that of Iba1-positive cells found in the lesioned areas. A cell division marker Ki67 revealed that few microglia were proliferating in crowberry-treated animals. Otherwise dividing cells were associated with blood capillary cells. Although the antioxidant level should be equal in the entire brain, no regeneration was found in globus pallidus, suggesting the mechanism promoting regeneration in the striatum is not effective in the globus pallidus. In conclusion, diets rich in bilberries and crowberries and with high contents of antioxidants stimulate an early phase of accumulation of reactive migroglia that fades at longer time points i.e. promotes regeneration of the striatal dopamine system.
Introduction
Neuroinflammation is proposed to be a contributing factor in neurodegenerative disorders. The presence of activated microglia in the brains of patients suffering from Parkinson's disease was frst demonstrated in 1988. 1 Activated microglia produce a variety of noxious compounds such as proinflammatory cytokines and, consequently, the levels of several cytokines, such as tumor necrosis factor alpha (TNFa) and interleukin-1β (IL-1β), are elevated in the brains of parkinsonian patients. 2 4 Similar changes in levels of TNFα are found in animal models of Parkinson's disease, wherein the dopamine neurons in the substantia nigra express TNFα receptor 1, suggesting that TNFα affects the nigrostriatal dopamine system. 5 7 These cytokines can exert direct toxic effects but may also induce toxic effects indirectly, for instance, via the inducible form of nitric oxide synthase.8,9
To suppress neuroinflammation, antioxidant supplementation is efficient.10,11 Antioxidant treatment may involve a diet high in antioxidants, for example, a diet rich in fruits and vegetables. Epidemiological studies have demonstrated that fruits and vegetables are beneficial to health and that they can lower the risk of neurological disorders.2,12–16 An antioxidant-enriched diet reduces neuroinflammation, improves behavioral declines during aging, and is neuroprotective in animal stroke models. 17 21 The mechanism for improvement of the motor system in aging has been attributed, at least in part, to enhanced dopamine release, especially after treatment with a diet enriched with blueberries. 19 Blueberry-enriched diets can also affect survival of grafted dopamine neurons, as well as induce regeneration of striatal dopamine nerve fibers after a dopamine lesion. 22 24
The rat model of Parkinson's disease that utilizes striatal administration of 6-hydroxydopamine (6-OHDA) results in a degenerated zone of dopamine nerve fibers surrounding the injection track, followed by slow, progressive degeneration of the ventral mesencephalic dopamine neurons. 25 Feeding the animals with a diet enriched with blueberries rescues neither the dopamine neurons nor the degree of striatal dopamine degeneration. 24 However, the neuroinflammatory response is changed such that an initial, transient infiltration of reactive microglia is present. In comparison, in animals that have received control diet the microglia are slowly activated over time. The rapid onset of activated microglia was not expected and it is still elusive if such a fast onset of activated microglia is a common event when treated with antioxidant-enriched diet. The main antioxidative compounds in blueberries are the anthocyanins, which have antiinflammatory qualities due to the inhibition of lipid peroxidation and cyclooxygenase 1 and 2 activity. 26 29 There are several berries that have a similar profile of anthocyanins as blueberry (Vaccinium corymbosum), for example, bilberry (Vaccinium myrtillis) and crowberry (Empetrum nigrum). Crowberry has the highest contents of anthocyanins among these berries. 30 This study was performed in order to investigate the effects of blueberries, bilberries, and crowberries on the nigrostriatal dopamine system, and the consequences of a dopamine lesion on the inflammatory response.
Materials and Methods
Animals
Female Sprauge-Dawley rats (150 g; Scanbur B&K, Sollentuna, Sweden) were housed on a 12-hour light/dark cycle at constant temperature (21°C) with free access to water and food-pellets. The rats were fed with R-34 diet (n = 10) or R-34 diet supplemented with 2% blueberry (n = 10), bilberry (n = 9), or crowberry (n = 10) throughout the study. The 6-hydroxydopamine lesions were performed four weeks after special diets were given and the rats were sacrificed one or four weeks after the lesion. All experiments were approved by the ethics committee for Northern Sweden (approval # A24/08, A68/12).
Striatal dopamine lesions
Four weeks after the antioxidant-enriched diets, the rats were anesthetized with 4% isofluran (Baxter Medical AB, Kista, Sweden) and mounted in a sterotaxic frame. A unilateral injection of 6-OHDA (Sigma, Stockholm, Sweden) was performed into the right dorsal striatum. The following coordinates were used: 1.0 mm anterior and 2.8 mm lateral to bregma, and 5.0 mm below the dural surface. A dose of 20 μg 6-OHDA, dissolved in 4 μl saline containing 0.02% ascorbic acid, was injected into the dorsal striatum at a rate of 1 μl/min using a Hamilton syringe. The cannula was left in place for 2 minutes following infusion. One week or 4 weeks post striatal dopamine depletion, the rats were anesthetized with pentobarbital and transcardially perfused with 50 ml of Ca2+ -free Tyrode solution, followed by 200 ml of 4% paraformaldehyde in 0.1 M phosphate buffer (pH = 7.4). The brains were then dissected and postfixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH = 7.4) for between 1 and 1.5 hours and then rinsed in 10% sucrose in 0.1 M phosphate buffer.
Immunohistochemistry
The brains were rapidly frozen in gaseous CO2. Coronal cryostate sections (14 μm) were collected from the striatum and then thawed onto gelatine-coated glass slides. The sections were washed 2 × 15 min in 0.1 M phosphate buffered saline (PBS; pH = 7.4), followed by incubation for 48 hours at 4°C with the primary antibodies. After a 3 × 10 minutes wash in 0.1 M PBS, the sections were incubated for 1 hour at room temperature with secondary antibodies. Following a 3 × 10 min wash in 0.1 M PBS, the sections were finally mounted in 90% glycerol in 0.1 M PBS. All antibodies were diluted in 0.3% Triton-X-100 in 0.1 M PBS and incubations were performed in a humidified chamber.
Microglial cells were detected with the primary antibodies Iba1 (rabbit anti-human, 1:1000, cat # 019-19741, Wako Chemicals, Neuss, Germany), OX-6 (mouse anti-rat, diluted 1:200, cat # MCA46R, Serotec, Hamar, Norway) and ED1 (mouse anti-rat, diluted 1:100, cat # MAB1435, Chemicon-Millipore, Solna Sweden). Antibodies raised against Ki67 (mouse anti-human, diluted 1:200, cat # NCL-Ki67-MMI, Immunkemi F&D AB, Järfälla, Sweden) were utilized to visualize dividing cells. The dopamine denervation was evaluated using antibodies against tyrosine hydroxylase (TH; rabbit anti-rat, diluted 1:300, cat # P40101, Pel-Freez, Arkansas, USA). The secondary antibodies used were Alexa Fluor A594-conjugated antibodies (goat anti-rabbit, diluted 1:500, cat # A-11037) and Alexa Fluor A488-conjugated antibodies (goat anti-mouse, diluted 1:200, cat # A-11029), both obtained from Invitrogen (Oregon, USA).
Image analysis
The diameter of the TH-negative zone following striatal 6-OHDA injection was measured as the distance between the TH-positive area on each side of the injection trace. The densities of TH-positive nerve fibers in the globus pallidus and microglial cells were measured and expressed as mean grey values. The densities of Iba1-, OX-6-, and ED1-positive microglia were determined in dopamine-depleted striata at the level of injection trace and in the globus pallidus. Images were captured with a CCD camera (ProgRes C14, Jenaoptic) with a 20x objective and analyzed using the NIH Image (Image macro) software. The optical density of the immunohistochemical images was determined on binary images in a standardized frame and expressed as grey value set from 0 to 255, where 0 represents 100% black and 255 100% white. All measurements were performed on blind-coded slices over three brain sections for each parameter and collected from each brain and brain area. The mean value from each brain area was calculated and further analyzed.
Statistical analysis
Two-factor ANOVA was performed to detect differences between treatments over time. To detect differences between treatments at each time point, one-factor ANOVA was performed followed by additional Bonferroni post hoc comparison. To determine differences within the treatment, a t-test was performed. All results are expressed as mean ± SEM. The significance level was set at P < 0.05.
Results
Regeneration of TH-positive nerve fibers
TH-immunohistochemistry was used to visualize the dopaminergic innervation in the striatum and in the globus pallidus. In all animals, a TH-negative zone was found in the striatum. The trace from the injection needle was visible at the center of the TH- negative area. The border between the TH-dense area and the TH-negative zone was distinct in all animals at one week, whereas at 4 weeks post-lesion the borderline of the TH-positive innervation was less distinct, especially in animals treated with diets supplemented with berries. The interaction effect between time and treatment significantly affected the size of the TH-negative zone (F = 3.180, P = 0.038, two-factor ANOVA). One week post-lesion, no differences were found when comparing the size of the lesion between controls and any of the treatments, while the size of the TH-negative diameter was significantly reduced in animals fed with bilberries compared to controls 4 weeks post-lesion (F3,15 = 3.478, P = 0.043; Fig. 1). The diameter of the TH-negative zone was significantly reduced 4 weeks post-lesion compared with the size measured at 1 week when comparing within bilberry (t7 = 8.159, P = 0.0001) and crowberry diets of treatment (t8 = 3.114, P = 0.014; Fig. 1). Thus, bilberry and crowberry treatments improved regeneration by sprouting from spared nerve fibers, as revealed by measuring the diameter of the TH-negative zone over the injection track.
TH- and ED1-immunohistochemistry of the injured striatum (
TH-immunohistochemistry in the globus pallidus revealed that few scattered nerve fibers were present in the dopamine-denervated side (Fig. 1). Thus, the TH-positive nerve fibers in the globus pallidus had degenerated 1 week post-lesion, which was true for all treatments. Statistical evaluations on TH-positive nerve fiber density, as revealed by mean grey value, demonstrated similar density in the globus pallidus independent of time after the lesion or given diet. Thus, no regeneration was found in the globus pallidus (Fig. 1).
Iba1-positive microglia is increased after antioxidant treatment
Iba1-immunohistochemistry, a pan-microglia marker, was used to investigate the general microglial profile in the dopamine-depleted striatum (Fig. 2). In all animals, Iba1-immunoreactivity revealed an even distribution over the brain, with increased density over the injured area in the striatum and in the globus pallidus. In the lesioned striatum, Iba1-positive cells covered the entire needle track with a dense cluster in the center of the injection site (Fig. 2). The density of Iba1-positive cells in the denervated striatum was affected both by diet and time (time effect F = 9.361, P = 0.005, diet effect F = 6.624, P = 0.001, interaction effect of time and diet F3,38 = 10.465, P = 0.0001, two-factor ANOVA; Figs. 2 and 3). The density of Iba1-positive microglia over the injection track was significantly higher in all animals given an antioxidant-enriched diet compared to controls 1 week post-lesion (F3,18 = 11.016, P = 0.0001, one- factor ANOVA). This increased density of Iba1-positive microglia was significantly reduced 4 weeks post-lesion in treated animals when comparing to their respective 1-week time points (bilberry t7 = 3.592, P = 0.009, blueberry t8 = 2.350, P = 0.047, and crowberry t8 = 3.291, P = 0.011), while the density of Iba1-positive cells was significantly increased in controls (t7 = 4.576, P = 0.003; Fig. 3A). Thus, 4 weeks post-lesion, all animals had a similar density of Iba1-positive microglia over the center portion of the injection track.
Iba1- and OX-6-immunoreactivity in the striatum of control (
Peripherally to the needle track and still within the TH-negative zone in the striatum, Iba1-positive cells were less dense (Fig. S1). Four weeks post-lesion, the distribution appeared more restricted to a smaller area in the animals fed with antioxidant-enriched diet than in the controls (Fig. 2). Furthermore, the morphology of the Iba1-positive cells located in the center of the injection site appeared activated, displaying rounded cell bodies and no processes. The cells at the periphery of the TH-negative zone had longer ramified processes, although the cells were larger than those on the intact side. This was true for all animals. However, in those fed an antioxidant-enriched diet, the presence of rounded cells appeared much more pronounced than in the controls, where the Iba-1-positive cells possessed cellular processes (Fig. 3C–F). Furthermore–-and particularly in bilberry-fed animals–- these rounded Iba1-positive cells were located into the myelinated fiber tracts in the striatum, while in the controls Iba1-positive cells were not frequently found in the fiber tracts (Fig. 3E and F).
The density of Iba1-positive microglia in the striatum was significantly higher in animals treated with bilberry, blueberry, and crowberry enriched diets when comparing to animals fed with control diet 1 week post-lesion (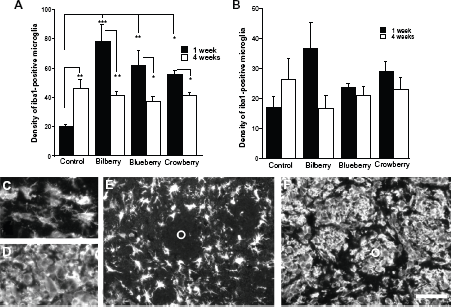
In the globus pallidus, no significant difference in density of Iba1-positive cells was found between the different treatments of animals, at any time point (Fig. 3B).
The presence of ED1-positive cells is affected by antioxidant-enriched diet
The microglial phenotype was further investigated since the morphology of the Iba1-positive cells suggested that the microglial activity differed between the cells localized in the center of the injection site and cells located peripherally to the TH-negative zone (Fig. 1A–C). ED1-immunoreactivity revealed co-reactivity with Iba1 (Fig. S1) and the presence of positive cells was found both 1 week and 4 weeks post-lesion. In the striatum, the location of ED1- positive cells was restricted to the TH-negative zone in all animals. Diet, but not time, had an impact on the density of ED1-positive cells in the dopamine-depleted striatum, and an interaction effect between diet and time was found (diet effect F3,38 = 8.058, P = 0.0001; interaction effect of time and diet F3,38 = 3.985, P = 0.017; two-factor ANOVA). One week post-lesion, the highest density of ED1-positive cells was found in the close vicinity of the injection track in animals given the bilberry-enriched diet, while the ED1-positive cells had a more widespread location in the controls (Fig. 1). In animals with high density of ED1-postive cells, the cells were evenly distributed in the striatum, both within and between the myelinated fiber bundles, while in the control animals the myelinated fiber bundles were mostly devoid of ED1-positivity (Fig. 4C and D). This accumulation of ED1-positive cells in the myelinated fiber bundles had been cleared at 4 weeks post-lesion. Thus, the distribution was similar to the presence of Iba1-positive cells. Measurements of the cell density over the injection site revealed a significant increase in bilberry-treated animals when comparing to controls 1 week post-lesion (F3,18 = 5.824, P = 0.008, one-factor ANOVA, Bonferroni; Fig. 4A). Furthermore, the increase in ED1-positive cells in bilberry-treated striatum was also significantly higher than in crowberry-treated animals (P = 0.0039) 1 week post-lesion. Four weeks post-lesion, ED1-positive cells were also clustered along the injection trace in the controls; however, this did not change the density compared to the 1-week time point (Figs. 1 and 4). A reduction was found in bilberry-treated animals between 1 week and 4 weeks post-lesion (t7 = 5.420, P = 0.001), and the myelinated fiber bundles were cleared from ED1-positive cells. The lowest density of ED1-positive cells was found in the striatum of blueberry-fed animals, which was significantly lower than in animals fed with both bilberry- and crowberry-enriched diets but not in controls (F3,18 = 6.146, P = 0.006 one-factor ANOVA; bilberry P = 0.007; crowberry P = 0.039; Bonferroni).
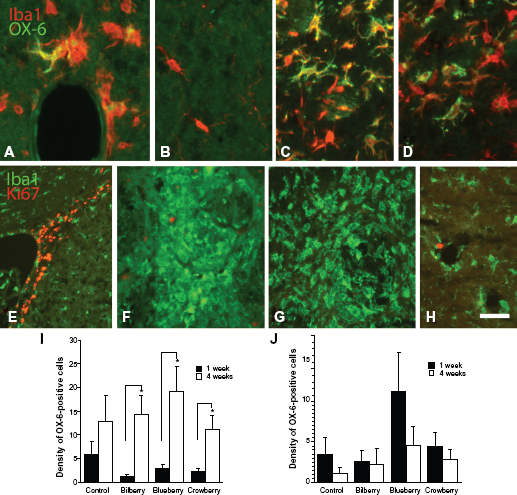
In the globus pallidus, ED1-positive cells were found both 1 week and 4 weeks post-lesion ipsilateral to the lesion, while no ED1-positive cells were found in control sides (Figs. 1 and 4). At the 1 week time point, the density of ED1-positive cells were significantly higher in animals fed with a bilberry-enriched diet compared to controls or any other treatment (F3,17 = 13.263, P = 0.0001, one-factor ANOVA, Bonferroni; Fig. 4). However, this increase was reduced at 4 weeks compared to the one week time point (t7 = 2.403, P = 0.047). Four weeks post-lesion, no difference was found in any of the animal treatments.
The density of ED1-positive microglia was significantly increased in both the striatum (
The presence of OX-6-positive microglia increases over time
MHC class II-positive microglia were visualized utilizing OX-6-immunohistochemistry. One week post-lesion, the majority of the cells were, regardless of diet, localized to the periphery of the high-density zone of Iba1-positive microglia. Four weeks post-lesion, the cells were also observed in the injection track (Fig. 2). No OX-6-positive cells were detected peripherally to the TH-negative area or in the intact hemisphere (Fig. 5B). Time significantly affected the density of OX-6-positive microglia (F1,38 = 21.488, P = 0.0001, two-factor ANOVA), whereas treatment did not. The density of OX-6-positive cells in the injection site of the striatum was significantly increased 4 weeks compared to the 1 week time point in all animals treated with an antioxidant-enriched diet, but not in controls (bilberry t7 = -2.802, P = 0.026; blueberry t8 = -2.892, P = 0.020; crowberry t8 = -3.031, P = 0.016; Fig. 5I). In all animals, there was a sharp borderline between the presence and absence of OX-6-positive cells 4 weeks post-lesion (Fig. 2). OX-6- as well as Iba1-positive cells were found clustered around capillary walls (Fig. 5A). In the globus pallidus, the density of OX-6-positive cells showed similar levels at both the early and late time points, regardless of diet (Fig. 5J). No OX-6-positive cells were found in the control side.
Iba1- and OX-6-positive microglia in the striatum of control (
Microglia accumulation is not due to cell division
A cell division marker Ki67 was employed in attempts to study if all microglia that were invaded at 1 week post-lesion were accumulated by cell proliferation. The results revealed that no Ki67-positive nuclei were found in the dense Iba1-positive area around the injection site in the striatum, in either controls or animals treated with bilberry- and blueberry-enriched diets (Fig. 5G). However, in all animals fed with crowberry-enriched diet, a few, widely spread Ki67-positive cells were found in the microglia-invaded area (Fig. 5F). In all animals, few Ki67-positive nuclei were found associated with the blood capillary endothelial cells (Fig. 5H). In all animals, Ki67-positive cells were located in the subventricular zone, which was used as a positive control for the otherwise negative striatum (Fig. 5E).
Discussion
The results from this study demonstrate that dopaminergic regeneration occurs in the striatum of animals fed with bilberry- and crowberry-enriched diets, while no regeneration of TH-positive nerve fibers was detected in the globus pallidus. Together with the striatal regeneration, an overall increase in density of microglia was found in the striatum of animals treated with antioxidant-enriched diets compared to controls, as revealed by the pan-microglial marker Iba1 one week post-lesion. This increased density was reduced at the 4 week time point, while the density of Iba1-positive cells was increased between 1 week and 4 weeks post-lesion in control animals. The microglial density was most pronounced in the striatum of animals treated with a bilberry-enriched diet one week post-lesion. This early increase of Iba1-positive microglia in bilberry-fed animals was similar to the density of ED1-positive cells, while the presence of OX-6-positive cells was increased over time in all animals, independent on treatment. Furthermore, the ED1-positive cells were preferably found in the center of the dopamine denervated zone in the striatum one week post-lesion, while OX-6-positive cells were located to the periphery of the denervated zone.
The antioxidant treatment had no rescuing effect on the TH-positive nerve fibers, since the loss of TH-positive nerve fibers was similar in all animals and regions of the brain evaluated one week post-lesion. Although, the specific cell death of the dopamine neurons was not evaluated here, earlier results have demonstrated no rescuing effect, while other polyphenol-rich treatments have documented rescuing effects on the cell death of dopamine neurons.24,31–34 However, the TH-negative zone was significantly smaller in the striatum of animals treated with bilberry- and crowberry-enriched diets 4 weeks post-lesion, suggesting that regeneration had occurred in animals given berries with the highest levels of anthocyanins. 30 While the improvement of TH-positive nerve fibers was found in the striatum 4 weeks post-lesion, no such effect was documented in the globus pallidus. Thus, the positive effect of antioxidant-enriched diets in the striatum had no such effect in the globus pallidus, although it seems likely that the antioxidants should reach levels similar in the whole brain since anthocyanins can cross the blood-brain barrier and can be localized to several brain regions. 35 Therefore, these results suggest that the effect produced by bilberries and crowberries is not one of rescuing. Rather, it suggests that the effect is that of promoting regeneration of the dopamine system by the sprouting of spared nerve fibers through mechanisms that are effective in the striatum but not in the globus pallidus. In general, it is hard to achieve regeneration in the globus pallidus as reported by other studies concerning graft-induced regeneration, wherein the striatum attracts and the globus pallidus repels dopamine regrowth.36,37
In dopamine-depleted striatum, microglia becomes activated 3 days post-lesion, as determined by the morphology. 38 In the present study a moderate increase in overall microglia, as found with Iba1- immunoreactivity, was observed in control-fed animals, with an increased expression over time. Treatment with an antioxidant-enriched diet significantly increased the density of microglia at the early time point. Additionally, their morphology became more rounded and reactive. In contradiction to our study, other investigators have demonstrated that the microglia is less activated by antioxidant treatments. 39 41 However, those studies report the microglia activation at later time points than one week, which does not necessarily contradict our results, since at the later time point the density is lower than at the earlier time point. Furthermore, most cells showing an early response were positive to ED1, while the number of OX-6-positive microglia increased at the later time point in the present study. Since there was no difference in the presence of OX-6- positive microglia between controls and animals fed with antioxidant-enriched diets, the activation of the MHC class II pathway was likely not what produced the regenerative effect. Thus, heavy early microglia infiltration (represented by the ED1-positive cells) in the striatum in antioxidant-treated animals resulted in regeneration of TH-positive nerve fibers. The reason for this effect is still elusive, but one mechanism involved might be reactive microglia producing proinflammatory cytokines such as TNFa. The presence of TNFa soon after an injury such as striatal dopamine lesion has rescuing effects on the dopamine nerve fibers, while a slow increase of TNFa is devastating. 42 This dual effect of TNFa is mediated through different TNFα receptors.43,44 Furthermore, the ED1-positive cells are correlated to phagocytotic activity. 45 Thus, heavy initial infiltration by ED1-positive cells clear up debris to create a milieu permissive for regeneration.
Independent of treatment, most of the reactive microglia expressed ED1. The recruitment of microglia to the injured striatum or globus pallidus is thought to come mainly from innate microglia. However, recruitment from peripheral bone marrow cells might also be possible, especially since 6-OHDA may injure the blood-brain barrier (BBB). 46 Interestingly, there was only a minor proliferating activity in the striatal endothelial cells, as revealed by Ki67-immunoreactivity, where newly formed endothelial cells are correlated to BBB dysfunction. 47 However, it is probably unlikely that these few newly formed blood vessels can give rise to such as high a concentration of the ED1-positive cells as that found in the injured area of animals given antioxidant- enriched diet. On the other hand, the reactive microglia was not accumulated by cell division of microglia. While it is known that TNFa levels are increased after a dopamine depletion, high levels of TNFa may mediate vasogenic edema followed by infiltration of bone marrow cells.7,42,48,49 Thus, further studies are needed to clarify the location where the accumulation of ED1-positive cells are derived.
Besides the involvement of microglia in the regenerative process after antioxidant treatment, the polyphenols themselves may have direct actions on regeneration. For instance, the 6-OHDA lesion induces increased influx of iron to the dopamine neurons.50,51 However, flavonoids act as chelators to iron and inhibit oxidative stress. 52 Therefore, the presence of antioxidants may reduce the toxic environment in the lesioned area, which thus becomes permissive for dopaminergic regeneration.
In conclusion, diets enriched in bilberries or crowberries, ie, those with high contents of antioxidants, promote an early phase of reactive migroglia that fades at longer time points and promotes regeneration of the striatal dopamine system.
Author Contributions
Both authors conceived and designed the experiments, contributed in writing the manuscript, and approved the final manuscript.
Funding
This study was supported by the Swedish Research Council grant #09917, the Umeå University Medical Faculty Foundations, and National Institutes of Aging grant #AG4418.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including but not limited to the following: authorship and contributorship, conflicts of interest, privacy and confidentiality and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest.
Supplementary Figure

TH- and lba1-immunohistochemistry (A and B) in the striatum of control (A) and bilberry-fed (B) animals 4 weeks post-lesion. iba1-positive microglia are sparse distributed over TH-positive areas in the striatum as well as in the periphery of the TH-negative zone (A and B). Some iba1-positive cells co-express ED1-immunoreactivity (c). notes: Scale bar A and B = 100 μm; c = 20 μm.