
Abstract
L-Tryptophan (L-TRP) is a nutritionally essential amino acid and the kynurenine (KYN) pathway is the major route of L-TRP catabolism. Besides being synthesized for proteins, L-TRP and its metabolites have critical roles for the functions of nervous and immune systems. Many researches show that optimal amounts of L-TRP in diets depend on species, developmental stages, environmental factors and health status. We have shown that KYN pathway-related enzyme activities vary among species, tissue and cell types in physiological conditions. Furthermore, the response of these enzyme activities to systemic and/or central nervous system immune activation and inflammation depends on species and cell types. Thus, it is very important to choose appropriate animal species and cell types in which to evaluate the physiologic and pathologic effects of increased KYN pathway metabolism. We believe that understanding L-TRP metabolism among species and cell types provides a better idea for analysis of human pathological condition.
Keywords
Introduction
The kynurenine (KYN) pathway is the major route of L-tryptophan (L-TRP) catabolism and an anabolic source of nicotinamide-containing nucleotide. 1 In extrahepatic tissues, the first enzyme of the KYN pathway is indoleanime 2,3-dioxygenase-1 (IDO1), rather than tryptophan 2,3-dioxygenase (TDO). 2 This enzyme has a much broader substrate profile for indoleamine-containing compounds and its expression is strongly influenced by the state of the immune system. The IDO1 activity is potently induced by several proinflammatory cytokines and endotoxin. In contrast, TDO activity is increased by TRP and its analogues via actions at a distinct allosteric activation site. 2 Substantial increases in the TRP-KYN pathway metabolites occur in human brain, blood and systemic tissues during immune activation. Many animal model studies on the KYN pathway metabolism during immune activation and normal conditions have used nonhuman primates,3–5 gerbils,6–9 guinea pigs,10,11 mice12–14 and rats. 9 In the field of neuroscience, the rat is one of the most frequently used animal species; however, immunologists generally use mice. It is well known that IDO1 induction in lung following immune stimulation by LPS is substantially less in rats than in mice and other species. 15 On the other hand, in the transient global brain ischemia model, rats and gerbils have been used widely to investigate the molecular mechanism of selective neuronal death because of the vascular anatomy in the brain. Therefore, it is very important to well understand species difference and select animals for analysis of human pathological condition. Furthermore, studies in vitro have shown that not all human cells are capable of directly synthesizing quinolinic acid (QUIN) from L-TRP. The activities of kynurenine pathway enzymes and the production of kynurenine metabolites are depending on cell types. 16 IDO1 is the most important regulatory enzyme for KYN pathway, and kynurenine 3-monooxygenase (KMO), kynureninase and 3-hydroxyanthranic acid oxidase are also important determinates of whether a cell can make QUIN. Indeed, stimulation by proinflammatory cytokines have resulted in large increases of the IDO1 activity in most cell types, although the accumulated amounts of QUIN are very different. It has been shown that blood macrophages and monocyte-derived cells produced the largest amount of QUIN in accordance with the highest activities of KMO and kynereninase compared to other cell types. 16
L-TRP plays versatile roles in nutrition and physiology, particularly food intake, neurological function and immunity. Optimal amounts of L-TRP in diets likely depend on species, developmental stages, environmental factors and health status. We studied species and cell type differences in IDO1 activity and kynurenine pathway-related enzymes in response to systemic immune activation by LPS or IFN-γ, and central nervous system (CNS) inflammation. Here, we show that KYN pathway enzyme activities and metabolite concentrations vary with cell types and species.
Species Differences in TRP-KYN Pathway Metabolism in Physiological Conditions
In the CNS, several KYN metabolites are neuroactive and may play potential roles in inflammatory neurological diseases.17–20 As shown in Figure 1, L-KYN is converted to anthranilic acid (AA), 3-hydroxykynurenine (3-HK), and other metabolites in the systemic tissues. Furthermore, 3-HK is metabolized to 3-hydroxyanthranilic acid (3-HAA), which is a bioprecursor of QUIN. Our group measured serum and cerebrospinal fluid (CSF) L-KYN and AA concentrations in different animal species.
21
The concentrations of serum L-KYN in rabbits were 4 to 17 times higher than that those in the other species studied. CSF L-KYN concentrations were also the highest in rabbits. There were no large species differences in the ratio of L-KYN concentrations CSF/serum. Compared with L-KYN, serum AA concentrations in rabbits were only slightly higher than those in the other species. AA concentrations in CSF were the highest in rats and lowest in rabbits. Consequently, the ratio of AA/KYN in rat CSF was 198-fold higher than that in rabbit CSF. In addition, the AA ratio in CSF/serum in humans and rats were extremely high compared with those of rabbits and guinea pigs. Thus, it is likely that KYN pathway metabolism in the CNS varies with species, although the cause of this difference remains to be determined. Previous studies have suggested that L-KYN, AA, and 3-HK, at least in rat brain, are derived from the periphery;
22
however, it is possible that these species differences depend on the blood-brain permeability and/or transport capacity into the brain cells for certain KYN pathway metabolites. Our results suggested that blood-brain barrier permeability of KYN metabolites’ levels in rats and humans were higher than those of mice, gerbils, and rabbits. The principal bioprecursor of 3-HAA was shown to be AA rather than 3-HK in rat brain;
23
however, it is speculated that L-KYN is mainly metabolized to 3-HAA through 3-HK in the brain except in some species. We also determined tissue KYN pathway enzymes, IDO1, TDO, KMO, and kynureninase in rabbits, rats, gerbils, and mice (Table 1).
21
The activities of these enzymes vary among species and tissues. Lung IDO1 activity in rabbits was 146–516 times higher than the activity in other species, and rabbit liver KMO activities were lower by one order of magnitude compared to those of others. Furthermore, brain KMO activities were 12.3–23.2 times higher in gerbil than those on other species. We also showed that brain KMO activities in gerbils were much higher than other species,
8
although the physiological significance of this difference in gerbil brain is still unclear. KMO is predominantly a mitochondrial enzyme, whereas the other enzymes of the KYN pathway are predominantly located in cytosol extracts of the brain. It is possible that different subcellular as well as cellular localization of these enzymes or substrates is involved in the regulation of KYN pathway metabolism in the brain. Thus, both studies suggested that the different activities of KYN pathway enzymes, IDO1, kynureninase, and KMO, which convert L-TRP to L-KYN, L-KYN to AA, and 3-HK to 3-HAA, resulted in these species differences. In summary, KYN pathway enzyme activities and metabolite concentrations vary by species in normal conditions.
Schematic overview of the kynurenine (KYN) pathway. L-Tryptophan (L-TRP) is the extracellular fluid is transported into cells by a high-affinity tryptophan transporter. The first rate-limiting enzyme indoleanime 2,3-dioxygenase 1 (IDO1) catalyses the initial enzymatic step in the KYN pathway leading to the synthesis of a number of KYN metabolites. IDO1 is induced by several proinflammatory cytokines; therefore, KYN metabolism is increased during many inflammatory conditions. By contrast, glucocorticoid hormones increase transcription of tryptophan 2,3-dioxygenase (TDO) and peripheral degradation of L-TRP via the KYN pathway. The biological function and induction mechanism of IDO2 are still unclear and controversial. IDO1 is an important regulatory enzyme in the production of L-KYN in a broad spectrum of cell types. Low activity of kynurenine 3-monooxygenase (KMO) in some cells restricts the capacity to synthesize QUIN from L-TRP. Activities of tissue kynurenine pathway enzymes in rabbit, rat, gerbil, and mouse.

Species Heterogeneity in Response to Brain Injury and Systemic Immune Activation
Effects of systemic LPS administration on lung and brain IDO1 activity and hepatic TDO activity in gerbils, Sprague-Dawley rats and C57BL/6 J mice.
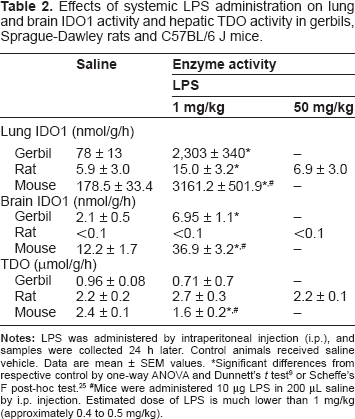
Significant differences from respective control by one-way ANOVA and Dunnett's t test 9 or Scheffe's F post-hoc test. 25
Mice were administered 10 μg LPS in 200 μL saline by i.p. injection. Estimated dose of LPS is much lower than 1 mg/kg (approximately 0.4 to 0.5 mg/kg).
Different KYN Pathway Enzymes Activities in Various Human Cell Types
Effects of LPS, IFN-γ and TNF-α on L-KYN and QUIN production in different cell types.

Significant differences from respective control by one-way ANOVA and Mann-Whitney test. 16
Effects of IFN-γ on KYN pathway enzyme activities in different cell types.

Significant differences from respective control by one-way ANOVA and Mann-Whitney test. 16
A New Enzyme in the Kynurenine Pathway; Indoleamine 2,3-Dioxygenase-2
As shown in previous sections, it has been believed that the first and rate-limiting step in the KYN pathway is performed by either of two enzymes, TDO or IDO1, depending on the tissue and cell type. Recently, a gene with homology to IDO1 was reported 26 and then subsequently demonstrated to be an enzyme with the ability to catabolize tryptophan.27–29 The enzyme has been referred to as indoleamine 2,3-dioxygenase-like protein, indoleamine 2,3-dioxygenase-2 or proto-indoleamine 2,3-dioxygenase (INDOL1, IDO2, protoIDO) on the basis of its structural similarity to IDO1 and its enzymatic activity. In mice, the IDO2 protein is predominantly expressed in the kidney followed by epididymis, testis, liver and brain.27,30 On the other hand, the IDO1 protein is expressed in various organs, like the brain, lung, placenta, epididymis and kidney under normal physiological conditions. Although both IDO proteins are expressed in the epididymis, IDO1 protein is located to the principal and apical cells of the mouse caput epididymis whereas the IDO2 immunoreactivity is in the tail of the spermatozoa. 27 Murine IDO1 protein is also located primarily in the renal vasculature of the kidney. 27 In contrast, IDO2 is located to the kidney tubules. The different patterns of expression suggest that there is no functional redundant amongst the these enzymes in the epididymis and kidney,27,30 but the role of each enzyme remains to be defined. In addition, expression of the proteins in response to stimuli differs. IDO1 is induced systemically dependent or independent on the presence of IFN-γ 25 while IDO2 mRNA expression is unchanged or down-regulated. 27 The Km for L-TRP of the human IDO2 protein was approximately 500-1,000 times higher than that of the mammalian IDO1 enzyme. 31 Furthermore, IDO activity of human DCs expressing both IDO1 and IDO2 is inhibited by IDO1-specific siRNA. 32 These studies indicate that IDO2 is functionally inactive with regard to the TRP metabolism and suggest that IDO2 might be a pseudo-gene for IDO1. However, the biological role of IDO2 is still controversial. We found that depletion of TRP and formation of KYN is detectable at physiological levels of TRP when mouse and human IDO2 enzymes are expressed and assayed in mammalian cells27,28 (Saito K, September, 2012). IDO2-like enzymes from lower vertebrates also show significant enzymatic activity when transfected into mammalian cells. 33 These findings might suggest that post-translational modifications of the protein are required for optimal activity. In additional, there is a possibility that different cofactors or conditions may be necessary for measuring IDO2 activity in vitro assay. The lack of an appropriate in vitro assay is a problem for biochemical characterization of IDO2.
Conclusions
We conclude that the KYN pathway enzyme activities and metabolite concentrations in serum and CSF vary with species in normal and pathologic conditions. In addition, immune activation in gerbils, induced by ischemic brain injury or systemic administration of LPS, replicates the increases in QUIN levels in human brain and systemic tissues in response to inflammation. In contrast to gerbils, neither LPS administration nor ischemic brain injury in rats had any significant effects on QUIN concentrations. Although different stimuli or strains of rats can increase brain and systemic tissue IDO1 activity and QUIN levels, it appears that the rat does not replicate human responses to immune activation. Taken together, these findings emphasize the importance of choosing an appropriate animal species in which to evaluate the physiological and pathophysiological effects of increased KYN pathway metabolism. Furthermore, L-KYN accumulated in several cell types; however, production of QUIN from L-TRP is accelerated in specific cell types, eg, macrophages, monocytes, microglia and hepatic cells. KYN pathway of TRP metabolism is involved in a lot of biological processes, including the immune system. We believe that further findings on TRP metabolism and its enzyme regulation in organisms under both physiologic and pathophysiologic conditions might provoke the implementation of novel therapeutic strategies by targeting the KYN pathway.
Funding
Part of this work was supported by Grant-in-aid for Scientific Research from Smoking Research Foundation. 13th ISTRY Satellite Symposium and the publication fee of this proceeding were supported by The International Council on Amino Acid Science (ICAAS).
Author Contributions
Wrote the first draft of the manuscript: YM. Contributed to the writing of the manuscript: KS. Jointly developed the structure and arguments for paper: YM, KS. Made critical revisions and approved final version: YM, KS. All authors reviewed and approved of the final manuscript.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Eethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.