
Abstract
Introduction
CX3CL1 (fractalkine), a membrane-bound chemokine that induces both the adhesion and migration of leukocytes, is involved in the recruitment of cells to tissues undergoing inflammatory responses. To explore the regulation of CX3CL1 in inflammatory bone diseases, we examined CX3CL1 expression in osteoblasts.
Methods
Human osteoblasts isolated from the femora of rheumatoid arthritis patients were incubated in the presence or absence of various inflammatory stimuli. Culture supernatants were collected, and soluble CX3CL1 levels were determined with an enzyme-linked immunosorbent assay (ELISA). The expression of CX3CL1 mRNA transcripts in osteoblasts was examined using the quantitative TaqMan real-time polymerase chain reaction.
Results
The combination of tumor necrosis factor (TNF)-α and interferon (IFN)-γ induced dramatic increases in levels of both soluble CX3CL1 protein and mRNA transcripts. CX3CL1 expression in osteoblasts was decreased by the addition of interleukin(IL)-4 or IL-17 but was increased when stimulation by IFN-γ and IL-17 was supplemented with IL-1β In addition, expression was decreased when TNF-α was added.
Conclusions
Multiple cytokines, including IL-17, are able to either increase or decrease the expression of CX3CL1 by human osteoblasts.
Keywords
Introduction
The pathology of rheumatoid arthritis (RA) is characterized by the infiltration of several inflammatory cell types into the pannus and joint fluid, followed by tissue destruction. Chemokines and other inflammatory mediators appear to play key roles in the pathogenesis of RA, and the coordinated production of chemokines and proinflammatory cytokines is likely to be important in the orchestration of the inflammatory response.1,2 Several cellular components of the joint contribute to the cytokine/chemokine network. Although autoreactive T cells, B cells and synovial cells (including synovial fibroblasts and macrophages) have crucial roles in pannus formation and arthritis formation, bone- derived cells such as osteoblasts (OBs), osteocytes and osteoclasts are also recognized as important cellular mediators of bone erosion and destruction in RA. 3 Several chemokines are highly expressed in bone erosive lesions. 4 The cells involved in bone formation (e.g. OBs) express a variety of chemokines, and OBs appear to be major regulators of bone remolding under both normal and pathological conditions.
During inflammatory processes, OBs exhibit prominent induction of cytokines and chemokines including TNF-α, IL-6, CXCL8, CXCL1, CCL2, CXCL9, CXCL10, CXCL11, ICAM-1, VCAM-1 and angiopoietin-1.5–9 Several proinflammatory cytokines such as TNF-α, IFN-γ, IL-6 and transforming growth factor-β have been shown to upregulate the expression of these molecules in OBs. Nevertheless, the exact profile of OB-derived chemokine expression remains unclear.
CX3CL1 (also known as fractalkine), is a member of the chemokine family that is synthesized mainly by endothelial cells (ECs). 10 The soluble form of CX3CL1 reportedly exerts a chemotactic effect on monocytes, NK cells and T lymphocytes. CX3CL1 acts as an adhesion molecule via its receptor CX3CR1 to promote the firm adhesion of a subset of leukocytes to ECs under conditions of physiological flow.11–12 Thus, CX3CL1 appears to possess immunoregulatory properties that affect inflammatory/immune cell-EC interactions and inflammatory responses. Indeed, several studies have implicated CX3CL1 in a variety of inflammatory disorders, including glomerulonephritis, systemic sclerosis and systemic lupus erythematosus.13–16 In particular, CX3CL1 may play important roles in RA and rheumatoid vasculitis.17–20 Then, we examined the relation between other cytokines (IL-1β, IL-4 and IL-17) and CXCL1 that took part in the inflammation of the rheumatoid arthritis.
Materials and Methods
Reagents
Complete medium consisted of DMEM (Nissui Pharmaceutical Co., Tokyo, Japan) supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 µg/ml streptomycin, and 10% heat-inactivated FBS (Gibco Laboratories, Grand Island, NY). TNF-α, IFN-γ, IL-1β, IL-4 and IL-17 were purchased from Genzyme/Techne (Cambridge, MA).
Preparation of Human osteoblasts
Human OBs were purified from metaphyseal trabecular bones in the proximal femora of RA patients (
Enzyme-Linked Immunosorbent assay
Soluble CX3CL1 was quantified using a double-ligand enzyme-linked immunosorbent assay (ELISA) modified from a previously described assay.16,20 Monoclonal murine anti-human CX3CL1 (Genzyme/Techne, 4 µg/ml) was used as the primary antibody (Ab), and the secondary Ab was biotinylated polyclonal goat anti-CX3CL1 (Genzyme/Techne, 0.25 µg/ml). This assay detects the chemokine domain of human CX3CL1, and the sensitivity limit is ~150 pg/ml. 20
Isolation of Total Rna and Real-Time Polymerase Chain Reaction(Pcr)
Total RNA was extracted from human OBs using TRIzol reagent (Invitrogen, Carlsbad, CA). One-microgram samples of total RNA were reverse transcribed into cDNA by incubation with TaqMan RT reagents (Applied Biosystems, Foster City, CA), first for 120 min at 37 °C and then for 10 min at 25 °C. Real-time PCR was conducted with an ABI Prism® 7900 sequence detection system (Applied Biosystems). The reaction mixture included 40 ng cDNA, which was amplified using AmpliTaq Gold DNA polymerase (Applied Biosystems) as described previously. 9 For detection of CX3CL1 (Applied Biosystems, catalog number 159394) and ribosomal RNA (rRNA) expression, appropriate Assays-on-Demand™ primers and probes (Applied Biosystems) were used in the PCR. For quantification, the number of target mRNA copies per rRNA copy was calculated, and values were expressed as -fold increases over control (culture medium alone). 20
Statistical Analysis
Data are expressed as the means ± SEM and were compared by analysis of variance. Post-hoc Student's t-tests were conducted to determine differences between specific groups. Values of
Results
Expression and Secretion of ob-Derived cx3cl1, and Regulation by cytokines
We previously reported that the expression and secretion of CX3CL1 by RA OBs are increased by co-stimulation with TNF-α and IFN-γ Here we show that the expression and secretion of CX3CL1 by RA OBs are regulated by IL-4 (20 ng/ml) (medium, 0 pg/ml; TNF-α+ IFN-γ, 4539 ± 101.1 pg/ml; TNF-α+ IFN-γ+ IL-4 0.2 ng/ml, 2643.3 ± 125.9 pg/ml; TNF-α+ IFN-γ+ IL-4 2 ng/ml, 2380 ± 116.8 pg/ml; TNF-α+ IFN-γ+ IL-4 20 ng/ml, 2144 ± 10.0 pg/ml; Figs. 1A and B). The time and concentration dependences of this effect were evaluated.
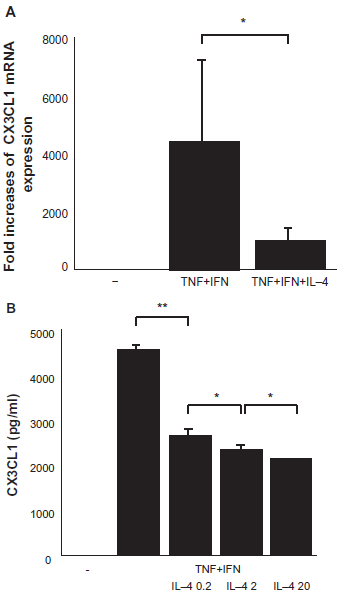
Expression and secretion of CX3CL1 by OBs, and effects of TNF-α, IFN-γ and IL-4. Cultured OBs from RA patients were incubated for 24 h as indicated with TNF-α and IFN-γ. The effect of adding IL4 was confirmed. (
IL-1β and TNF-α are both factors that regulate the inflammatory response. We performed co-stimulation experiments with IFN-γ. The expression and secretion of CX3CL1 by RA OBs were increased by co-stimulation, although there was little response to single stimulation with TNF-α. The time and concentration dependences of this effect were evaluated. Neither IL-1β (20 ng/ml) nor IFN-γ (100 units/ml) significantly affected CX3CL1 secretion from RA OBs compared to treatment with medium alone. When applied together, graded concentrations of different combinations of IL-1β and IFN-γ induced significant concentration- dependent secretion of CX3CL1 from RA OBs (medium, 0 pg/ml; IL-1β+ IFN-γ, 1103.3 pg/ml; Fig. 2).
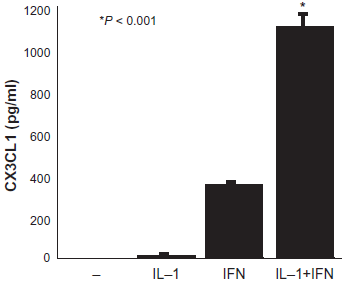
Secretion of CX3CL1 from OBs and the effects of IL-1β and IFN-γ. Cultured OBs from RA patients were incubated for 24 h with IL-1β and IFN-γ as Indicated.
Expression and Secretion of ob-Derived cx3cl1, and Regulation by il-17
The expression and secretion of CX3CL1 by RA OBs were suppressed by IL-17 (20 ng/ml) in response to co-stimulation with TNF-α and IFN-γ (medium, 0 pg/ml; TNF-α+ IFN-γ, 5003 ± 217.8 pg/ml; TNF-α+ IFN-γ+ IL-17 0.2 ng/ml, 3557.5 ± 257.9 pg/ml; TNF-α+ IFN-γ+ IL-17 2 ng/ml, 3104.2 ± 219.4 pg/ml; TNF-α+ IFN-γ+ IL-17 20 ng/ml, 1807.8 ± 311.5 pg/ml; Figs. 3A and 3B). This effect was concentration-dependent. By contrast, the expression and secretion of CX3CL1 by RA OBs increased with the addition of IL-17 (20 ng/ml) in response to co-stimulation with IL-1β and IFN-γ (medium, 0 pg/ml; IL-1β+ IFN-γ, 500 ± 34.4 pg/ml; IL-1β+ IFN-γ+ IL-17 2 ng/ml, 657.7 ± 48.6 pg/ml; IL-1β+ IFN-γ+ IL-17 20 ng/ml, 755 ± 66 pg/ml; Figs. 4A and 4B). This effect was also concentration-dependent.
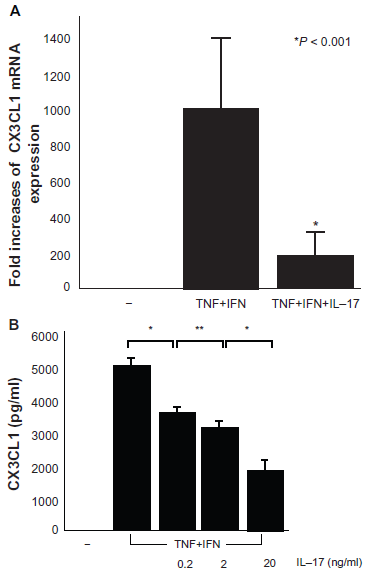
mRNA expression (
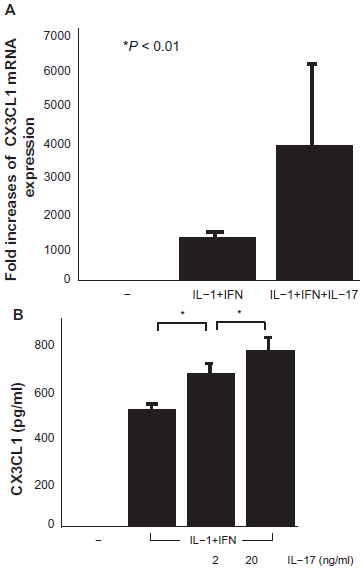
mRNA expression (
Discussion
The present study clearly demonstrates that OBs from RA patients are a prominent cellular source of CX3CL1 in response to a variety of cytokines. The influence of TNF-α and IFN-γ on OBs has also been reported by other groups. In 1987, Sato K et al used OB-like cells (MC3T3-E1) and reported the influence of TNF-α on the resorption of bone, highlighting its effects on human OBs. 22 The influence of TNF-α and IFN-γ on the remodeling of bone has also been reported previously. Gowen M et al reported the effects of TNF-α and recombinant human IFN-γ on the growth and function of cultured human OB-like cells, demonstrating that TNF stimulates bone cell proliferation and prostaglandin production while inhibiting 1,25-dihydroxyvitamin-stimulated alkaline phosphatase activity and osteocalcin release. 23
In this study, we show that IL-1β has a dynamic state similar to that of TNF-α, and in combination with IFN-γ, expression of CX3CL1 more than TNF-α. There was little expression of CX3CL1 induced by IL-1β alone, but expression was induced in combination with IFN-γ, similarly to effects observed for TNF-α with IFN-γ. Thus, IL-β was weaker than TNF-α at inducing the expression of CX3CL1. There was a previous report on the influence of IL-1β on OBs that used the osteoblastic cell line MC3T3-E1 to investigate the role of IL-1 in the regulatory mechanisms of bone remodeling. 24 IL-4 is a cytokine with known anti-inflammatory effects. The expression of CX3CL1 was reduced in OBs. We did not evaluate the response of OBs to IL-4, but IL-4 has been found to induce a concentration-dependent increase in the expression of alkaline phosphatase stimulated by 1,25- dihydroxyvitamin D3, a marker of differentiated OB activity. 25 This time, we do not examine blocking experiments of monoclonal antibody to the cytokines. It is necessary to be going to do this examination in the future.
Recent reports have focused on the role of IL-17 in RA. After Bettelli et al reported on Th17 and regulatory T cells in 2006,
26
examinations of the effects of IL-17 under various conditions have been carried out. IL-17 is highly produced by RA synovium.27,28 Furthermore, IL-17R mRNA is expressed by mouse osteoblastic cells and fetal mouse long bones, and IL-17 in combination with TNF-α, but not IL-1β, increases osteoclastic resorption
The effect of anti-IL-1β is comparable to that of anti-TNF-α in RA, possibly providing a clue as to why IL-17 appears to play a different role. IL-1β takes part in the activation of the osteoclast. On the other hand production of IL6 is accelerated through a human synovial cells for TNF-α. IL-6 acts on Th17, and production of IL-17 is accelerated. CXCL1 participates to destruction of a joint. It is suggested that it is controlled by IL-17 through TNF-α. Future studies of IL-17 will be needed to clarify this point.
The synergistic induction of CX3CL1 expression by various cytokines such as TNF-α, IFN-γ, IL-1β and IL-17 raises the question of which cytokine acts as the dominant inducer of CX3CL1.
In conclusion, there are still many uncertain points about the role of IL-17. A possible role for IL-17 in the regulation of CX3CL1 and the pathophysiology of RA is suggested by our results.
Disclosure
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers of this paper report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material.