
Abstract
This study was aimed at gaining an insight into immune mechanisms of differential susceptibility to autoimmunity of individuals sharing the same major histocompatibility complex by studying arthritis-susceptible Lewis (LEW) and arthritis-resistant Wistar Kyoto (WKY) rats (both RT.1l) using the adjuvant arthritis (AA) model of rheumatoid arthritis (RA). Lymph node cells (LNC) and synovium-infiltrating cells (SIC) of LEW and WKY rat subjected to an arthritogenic challenge were tested. The frequency of T helper 17 (Th17) and T regulatory (Treg) cells was determined by flow cytometry, whereas serum and spleen adherent cell (SAC)-derived supernatant were analyzed for specific cytokines and chemokines. We observed that WKY rats are not deficient in generating a Th17 response to the arthritogenic challenge in LNC (periphery); however, the Th17/Treg ratio is markedly reduced in the joint (target organ) of WKY versus LEW rats because of reduced Th17 levels therein in WKY rats. These results suggest differential and selective decrease in Th17 cell migration into the joints of WKY rats. Interestingly, serum levels of chemokines RANTES and MCP-1 were reduced in WKY rats. Furthermore, WKY rats showed reduced serum IL-1β level in vivo but no defect in IL-1β production by SAC in vitro, suggesting an effective in vivo regulation of IL-1β response. We also unraveled the role of interferon-γ (IFNγ), which we have previously reported to be increased in WKY versus LEW rats, in regulation of IL-1β. Thus, reduced Th17/Treg ratio in the target organ (joints) and decreased systemic IL-1β might contribute to the AA-resistance of WKY rats; whereas the converse factors render LEW more vulnerable to AA.
Keywords
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease of multifactorial origin. Susceptibility to RA is reported to be most strongly linked with major histocompatibility complex (MHC) haplotype. 1 However, the non-MHC genes as well as environmental factors also influence the induction and development of RA.2,3 Animal models of RA share characteristics of human RA, and have contributed extensively both to understanding of the pathogenesis of RA and to advances in the treatment of RA. 4 However, the precise mechanisms of disease susceptibility/resistance have yet to be understood fully. Furthermore, many of the previous studies in rodent models are based on testing strains having disparate MHC haplotypes.5,6 Testing of mouse/rat strains of the same MHC haplotype is vital for drawing more accurate conclusions regarding immunological factors influencing disease susceptibility. In this study, as described below, we have filled this gap for the rat adjuvant-induced arthritis (AA) model of RA by using rat strains of the same MHC haplotype.7,8 Another important aspect of disease induction and progression is the balance between T helper 17 (Th17) and T regulatory (Treg) cells. Th17 cells play a vital role in disease induction in arthritis, while Treg cells possess immunoregulatory activity. It has been reported by some investigators that RA patients are deficient in Treg in the peripheral blood,9,10 thereby also affecting Th17/Treg balance; but other investigators failed to observe this difference in their studies. 11 Regardless, there is barely any information in the comparative Th17/Treg balance in arthritis-susceptible versus arthritis-resistant rat strains in the AA model. In this study, we have addressed this important component of disease susceptibility/resistance in defined rat strains. Also tested were serum levels of pro-inflammatory cytokines such as IL-1β and IL-17, which play an important role in arthritis pathogenesis.12,13
Inbred Lewis (LEW) (RT.11) rats develop AA following subcutaneous (s.c.) injection of heat-killed Mycobacterium tuberculosis H37Ra (Mtb). The Wistar Kyoto (WKY) rat strain has the same MHC haplotype (RT-11) as the LEW rat but is resistant to AA. 14 Furthermore, the Mtb-immunized WKY rats are not deficient in T-cell proliferative response or antibody response to the disease-related antigen, mycobacterial heat shock protein 65 (Bhsp65). 15 Therefore, we reasoned that comparative studies on immune responsiveness of these two rat strains might provide insights into AA-susceptibility/resistance. We have previously reported the quantitative difference in the levels of expression and kinetics of pro-inflammatory vs. immune regulatory cytokines by lymph nodes cells (LNC) in response to Bhsp65 in LEW and WKY rats;14–16 a prominent finding then was increased IFNγ response to Bhsp65 in LNC of WKY compared to LEW rats. However, there is no information on the levels of either the cytokines in circulation or the frequency of T helper 17 (Th17)/T regulatory (Treg) cells in LEW vs. WKY rats.
In the present study, we examined the frequency of Th17 and Treg as well as the Th17/Treg ratio in the periphery (lymph node) and the target organ (joints) of LEW and WKY rats following Mtb injection. We also tested IL-17 and IL-1β in sera of these rats and the role of IFNγ in regulation of IL-1β. In addition, as inflammasome activation contributes to the production of mature (active) IL-1β,17,18 we examined the state of inflammasome activation in these rat strains. Inflammasome is a group of multimeric protein complexes that consist of a NOD-like receptor (NLR) sensor molecules (NLRP1, NLRP3), the adaptor protein ASC and caspase-1. Formation of active inflammasome complex can be triggered by recognition of a molecular pattern during infection, tissue damage, or metabolic imbalance. The activated inflammasome complex activates caspase-1, which then releases active IL-1β by proteolytic cleavage of pro-IL-1β.18,19 Furthermore, inflammasome activation plays an important role in the pathogenesis of several chronic inflammatory disorders. 20
Our results revealed that AA-resistant WKY rats are not defective in generating a Th17 response in the periphery (lymph nodes) following Mtb injection, but the level of Th17 cells is significantly reduced in the joints of these rats compared to arthritic LEW rats. On the contrary, WKY rats have increased levels of Treg in the periphery, but similar levels of Treg in the joints compared to LEW rats. Furthermore, WKY rats have significantly reduced serum IL-1β and chemokines RANTES (Regulated on Activation, Normal T Cell Expressed, and Secreted) and MCP-1 (Monocyte Chemoattractant Protein-1) compared to LEW rats. We suggest that the above factors contribute in part to AA-resistance of WKY rat; conversely, increased Th17 in the joints, reduced Treg in the periphery, and increased circulating levels of chemokines and IL-1β render LEW rats more vulnerable to AA compared to WKY rats.
Materials and methods
Animals
Male Lewis (LEW/SsNHsd) (=LEW) (RT.11) and Wistar Kyoto (WKY/NHsd) (=WKY) (RT.11) rats aged 5–6 weeks were purchased from Harlan Sprague-Dawley (HSD) (Indianapolis, IN, USA). These rats were housed in the vivarium of the University of Maryland School of Medicine, Baltimore, MD, USA (UMB). Maintenance of these rats and all experimental procedures performed on them were in accordance with the guidelines of the institutional animal care and use committee (IACUC) of UMB.
Induction of adjuvant arthritis (AA) and grading of its severity
Heat-killed M. tuberculosis H37Ra (Mtb) (Difco Laboratories, Detroit, MI, USA) was suspended in mineral oil (Sigma-Aldrich, St. Louis, MO, USA). Groups of LEW and WKY rats were immunized with 200 µL of Mtb-mineral oil suspension (1.5 mg/rat) each at the base of the tail (s.c.). Beginning the day after immunization, these rats were observed regularly for the clinical signs of arthritis in the paws. The severity of arthritis in each paw was graded on a scale of 0 to 4 as described elsewhere.21,22 The maximum attainable arthritic score was 4 for each paw and 16 per rat. The hind paws were photographed and also subjected to radiographic examination to assess bone damage as described elsewhere. 23
Preparation of lymph node cells (LNC) and stimulation with PMA and ionomycin for cytokine testing
LEW rats were euthanized on day 10 (onset) or day 18 (peak) after Mtb immunization. The draining lymph nodes (para-aortic, inguinal, and popliteal) were harvested from these rats.
14
Likewise, WKY rats were euthanized at the corresponding time points and their draining lymph nodes were harvested. Thereafter, a single cell suspension of LNC was prepared, and the cells were washed three times with Hank’s balanced salt solution (HBSS) (Life Technologies, Grand Island, NY, USA). These LNC (2.5 × 106 cells/well of a 6-well plate in 2 mL of complete RPMI
Preparation of synovial-infiltrating cells (SIC) and their stimulation with PMA and ionomycin for cytokine testing
Hind paws of Mtb-immunized LEW and WKY rats were harvested on day 10 or day 18. Total SIC were collected by opening the joints using a sterile surgical blade. 14 These SIC were washed 3–4 times with HBSS and then cultured and stimulated as described above for LNC. 22
Flow cytometric analysis of SIC and LNC
SIC and LNC were stimulated with PMA and ionomycin as described above and then incubated with 10% normal rat serum for 15 min. The T cells among SIC and LNC were surface-stained with anti-CD3 APC, anti-CD4 FITC, and/or anti-CD25 PE (all from eBioscience, San Diego, CA, USA). 22 For intracellular staining, cells were fixed and permeabilized using the Fixation/ Permeabilization Kit (eBioscience, San Diego, CA, USA) followed by staining with anti-rat/mouse IL-17A eFlour 450 (eBioscience, San Diego, CA, USA) and/or anti-rat IFNγ (BioLegend, San Diego, CA, USA). 22 Foxp3 was stained using anti-mouse/rat Foxp3 eFlour 450 (eBioscience). Using LSRII flow cytometer (BD Bioscience) (Flow Cytometry Shared Service, UMB), lymphocytes were acquired after setting “gates” for size (forward scatter) and cell complexity (side scatter) and further gated for CD3+ and CD4+ T cells. Thereafter, CD4+IL-17A+ gates and CD25+Foxp3+ gates were used to quantify Th17 and Treg cell frequency, respectively. Th17/Treg ratio was calculated using percentage of cell numbers. Appropriate isotype controls were used to establish each positive signal. Data analysis was performed using FlowJo software (Tree Star).
Preparation of spleen adherent cells (SAC) and their re-stimulation with Mtb for cytokine testing
Spleens were harvested from LEW and WKY rats on day 10 or day 18 after Mtb injection, as well as from naïve rats. A single cell suspension of spleen cells was prepared and the cells were washed 3–4 times with HBSS. These spleen cells were placed in 60 × 15 mm culture dishes (Corning Incorporated Corning, NY, USA) at 37°C in complete RPMI. After 90 min, the non-adherent cells were removed by washing with DMEM, yielding the SAC that remained attached to the bottom of the culture dish. 21 These SAC (approximately 1.5–2.0 × 106 cells per dish) were re-stimulated with or without sonicated Mtb (10 µg/mL) in the presence or absence of different concentrations of IFNγ (PeproTech, Rocky Hill, NJ, USA) for 1–24 h. These cells were used then used for cytokine testing as described below.
Multiplex analysis of serum and SAC culture supernatant for cytokines and chemokines
Blood samples were collected from LEW and WKY rats on day 10 or day 18 after Mtb injection, as well as from naïve rats by tail vain bleeding. Then serum was prepared from these samples. Serum samples and/or SAC culture supernatants were analyzed for cytokines (IL-1β and IL-17) and chemokines (RANTES and MCP-1) using a Multiplex assay (Cytokine Core Facility, UMB). The assays were read using the Luminex system following the manufacturer’s recommendations.
Western blot analysis
Cell lysates of SAC were prepared using cell lysis buffer (Cell Signaling Technology, Danvers, MA, USA). The protein concentration of the lysates prepared from SAC was measured using a BCA protein assay kit (Pierce). The proteins (20 μg) in the lysate supernatant were separated by SDS-PAGE (4–20%). Thereafter, the proteins were transferred onto a PVDF membrane. The membrane was first blocked with 5% milk in Tris-buffered saline containing 0.05% Tween, followed by probing with anti-IL-1β antibody (Abcam, Cambridge, MA, USA). Then the same membrane was re-probed with an antibody against tubulin. Protein bands were visualized with chemiluminescent HRP antibody detection reagents (Denville Scientific Inc, Denville, NJ, USA) following incubation with secondary anti-rabbit antibody (Pierce, Rockford, IL, USA) according to the manufacturer’s directions.
Preparation of RNA, cDNA, and quantification of IL-1β mRNA expression using qRT-PCR
RNA was isolated from SAC using trizol reagent (Invitrogen corporation laboratory services, Carlsbad, CA, USA) and then cDNA was prepared from RNA using iScript cDNA synthesis kit (Bio-Rad laboratories, Hercules, CA, USA). The cDNA thus prepared was amplified in an ABI Prism 7900HT cycler (Applied Biosystems, Foster City, CA, USA) by qRT-PCR using IL-1β primer and SYBR green PCR master mix (Applied Biosystems Inc., Foster City, CA, USA). The mRNA levels of the genes of interest were normalized to the hypoxanthine-guanine phosphoribosyltransferase (HPRT) gene, and the relative gene expression levels were determined. “Fold increase” over mRNA levels of untreated cells was then determined. Primers were designed using the Primer Express 2.0 Program (Applied Biosystems, Foster City, CA, USA) and then obtained from Sigma.
Statistical analysis
The data were expressed as mean ± SEM. Student t-test and ANOVA were used as appropriate to assess the significance of differences using GraphPad Prism version 4.0. A P value of <0.05 was considered statistically significant.
Results
We have previously reported that LEW rats are susceptible, while WKY rats are resistant to Mtb-induced AA. 15 We validated this disease-susceptibility/resistance profile again before undertaking a mechanistic study to this effect. We immunized groups of LEW and WKY rats with Mtb s.c. and then observed these rats for 25 days thereafter for signs of AA. The signs of arthritis started appearing in LEW rats after about 8 days of Mtb injection and reached a peak level at about days 18–20 followed by gradual regression (Figure 1a). Photographs of hind paws are shown in Figure 1b. In LEW rats, the joint damage in the form of bone destruction was evident in X-ray photographs (Figure 1c). In contrast, WKY rats failed to develop clinical signs of arthritis after Mtb injection throughout the observation period (Figure 1a and b). No bone damage was observed in X-ray photographs of hind paws of WKY rats (Figure 1c).
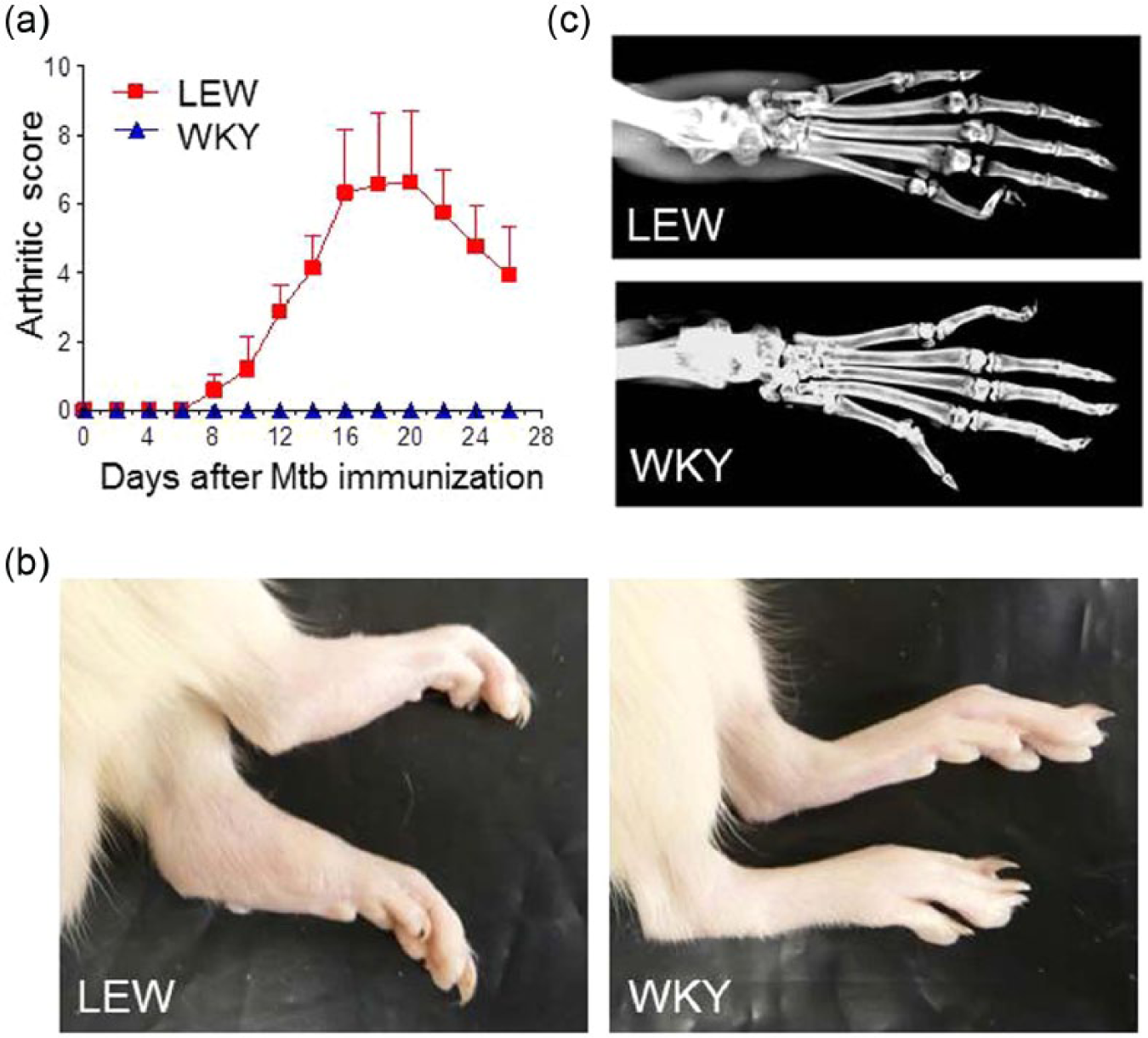
LEW/WKY rats display differential susceptibility to AA. WKY rats are highly resistant to AA induction and are protected against arthritic inflammation and joint damage compared to LEW rats. (a) Age- and sex-matched LEW and WKY rats (n = 6 per group) were immunized with heat-killed Mtb s.c. for arthritis induction. Thereafter, these rats were observed regularly and graded for signs of arthritis from day 0 through day 25. (b) Photographs of the hind paws (LEW, left; WKY, right) and (c) X-ray images of a representative LEW (top) and WKY (bottom) rat on day 18 after Mtb-injection are shown.
We tested the frequency of Th17 and Treg in the lymph node cells (LNC) (periphery) and synovial-infiltrating cells (SIC) (target organ) of LEW and WKY rats on day 10 (onset) and day 18 (peak) after Mtb injection (Figure 2a). In the case of Th17 cells in the periphery (lymph nodes) (top panel), there was a trend towards an increase in the frequency of these cells in both LEW as well as WKY rats on days 10 and 18 of Mtb immunization. Furthermore, at each phase, Th17 frequency in LNC of LEW and WKY rats was comparable. In the case of Th17 cells in the target organ (joints) (bottom panel), there was an increase in the frequency of these cells in arthritic LEW rats with the progression of AA. In contrast, the WKY rat joints showed much lower frequency of Th17, which did not change much in the period following Mtb injection. Specifically, the frequency of Th17 cells in the joints of arthritic LEW rats at days 10 and 18 was significantly (P <0.05) higher than that of WKY rats. Similarly, the frequency of a subset of IFNγ+IL-17+ T cells (double producers) (Figure 2a) was significantly (P <0.05) higher in the joints of LEW rats compared to that of WKY rats with the progression of AA in the former. A similar trend was observed for the double producers in LNC of LEW vs. WKY rats, but the difference was not statistically significant.

The frequency of Th17, Treg, and IFNγ+ IL-17+ T cells, as well as the Th17/Treg ratio in LEW and WKY rats on different days after Mtb injection. WKY rats are not deficient in generating a Th17 response to Mtb injection, yet are protected from AA. The Th17/Treg ratio is comparable in the lymph nodes of the two rat strains but significantly reduced in the joints of WKY rats compared to LEW rats. (a) Lymph node cells (upper panel, n = 10–12 per group) and synovial-infiltrating cells (SIC) from the joints (lower panel, n = 3–4 sets of pooled SIC from 2–3 rats) were prepared from Mtb-immunized LEW and WKY rats at the indicated time points after Mtb injection. Then, the cells were analyzed by flow cytometry for the frequency of Th17, Treg, and IFNγ+ IL-17+ T cells (double producers). (b) Changes in Th17/Treg ratio was derived from the frequency of Th17 and Treg cells in LEW and WKY rats at the respective time points. (*, P <0.05 when comparing LEW and WKY at each time point).
In contrast to Th17 cells, the frequency of Treg (Figure 2a) in the lymph nodes (top panel) of WKY rats was higher than that of LEW rats at each of the three phases of AA tested. A similar trend was observed in the joints (bottom panel), but the difference was not statistically significant. We also determined the Th17/Treg ratio in the samples from these tissues at the three phases of AA (Figure 2b). The Th17/Treg ratio showed an increase in LNC as well as the joints of LEW rat during the progression of the disease. In comparison, at each stage, the ratio of these cells in LNC in WKY was similar to that of LEW rats, but in the joint the Th17/Treg ratio was significantly reduced in the case of WKY rats at each phase tested compared to LEW rats.
The levels of cytokines IL-1β and IL-17 (Figure 3a) and chemokines RANTES and MCP-1 (Figure 3b) were determined in sera from LEW and WKY rats collected on day 10 (onset) and day 18 (peak) of AA. Also tested was serum from naïve rats. The levels of IL-1β and RANTES at the onset and peak phase, and that of MCP-1 at the peak phase were significantly increased in LEW rats compared to those of WKY rats at the corresponding time points. IL-17 levels in LEW and WKY rats were comparable at both time points tested. The levels of these cytokines/ chemokines in sera of naïve rats were either below detection levels or not significantly different between the two rat strains.

Serum levels of key pro-inflammatory cytokines and chemokines in LEW and WKY rats. AA-resistant WKY rats show reduced serum levels of IL-1β, RANTES, and MCP-1 compared to AA-susceptible LEW rats. Serum was prepared from the blood samples collected from Mtb-immunized LEW (filled bar) and WKY (open bar) rats at the indicated time points after Mtb injection, as well as from naïve rats. Then the sera were analyzed for (a) cytokines IL-1β and IL-17, and (b) chemokines RANTES and MCP-1 using a Multiplex assay. The results obtained from two sets of pooled serum samples from 3–6 rats tested in triplicate are shown. (*, P <0.05 when comparing LEW and WKY at each time point; BD, below detection.)
We then tested in vitro IL-1β production by SAC of LEW and WKY rats. SAC were prepared from the two rat strains at the onset phase of AA after Mtb injection and then restimulated with or without Mtb for different time points. Cell lysates were prepared and analyzed by Multiplex for IL-1β (Figure 4a) or western blot for pro-IL-1β and active (mature) IL-1β (Figure 4b). Although serum IL-1β levels (in vivo) were different between LEW and WKY rats (Figure 3a, top panel), surprisingly IL-1β level in the culture supernatants from both the rat strains were found to be comparable (Figure 4a). These results were confirmed by western blot analysis (Figure 4b). The levels of pro-IL-1β and mature (active) IL-1β in SAC of both the LEW and WKY rats showed an increase with time post-Mtb stimulation, but there was no difference either in pro-IL-1β or in active IL-1β production between the two strains.
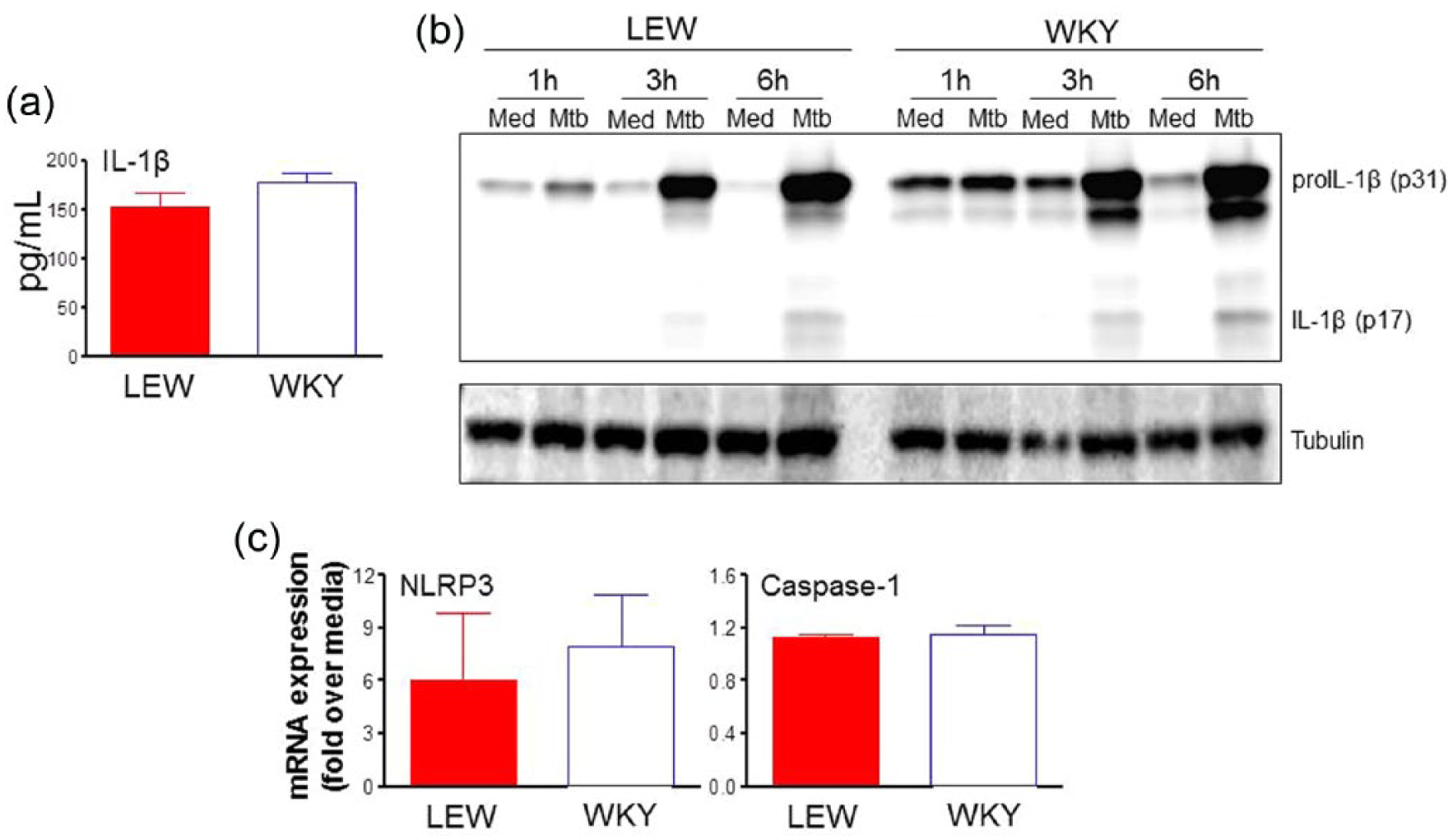
Assessment of the production of IL-1β in vitro by spleen adherent cells (SAC) of LEW and WKY rats. WKY rats were not deficient in IL-1β production by SAC exposed to Mtb in vitro. The level of IL-1β in the culture supernatant and the status of inflammasome activation in WKY rats were comparable to that of LEW rats. SAC from Mtb-immunized LEW and WKY rats were prepared at about the onset of AA. (a) Cells were restimulated in vitro for 24 h with Mtb sonicate (10 μg/mL). Thereafter, cell-free culture supernatants were collected, and the levels of IL-1β were measured by a Multiplex assay (n = 3). (b) SAC were restimulated in vitro for 1–6 h with or without Mtb sonicate. Lysates were prepared from these cells and the levels of pro-IL-1β (p31) and mature IL-1β (p17) were analyzed by western blot. Tubulin was used as a loading control. A representative blot is shown. (c) SAC were restimulated for 6 h and then RNA was isolated and analyzed for NLRP3 and caspase-1 mRNA expression by quantitative RT-PCR (n = 3). Values were normalized to the respective hypoxanthine-guanine phosphoribosyltransferase (HPRT) mRNA levels and were expressed as “Fold over medium”.
To further explore any differences in inflammasome activation in LEW and WKY rats, we tested the mRNA expression of inflammasome-related molecules, NLRP3 and caspase-1, in SAC of LEW and WKY rats. Again, there was no significant difference in the expression of either NLRP3 or Caspase-1 (Figure 4c). Finally, we tested the influence of IFNγ on IL-1β production by SAC. We observed that IFNγ treatment decreased the expression of IL-1β mRNA in SAC (Figure 5) as tested by qRT-PCR (top panel) and by western blot analysis (bottom panel). The above results suggests that IFNγ suppresses IL-1β primarily by inhibiting the production of pro-IL-1β (which is known to be driven by NF-kB pathway) without any measurable effect on inflammasome activation (which is required for the cleavage of pro-IL-1β into active IL-1β) (Figure 4b and c).

IFNγ suppresses IL-1β production in SAC. We have previously shown that WKY rats have increased IFNγ levels in response to antigen (Bhsp65) than LEW rats. 15 Here, we show that IFNγ can inhibit IL-1β response in vitro, supporting the finding of reduced serum (in vivo) level of IL-1β (shown in Figure 3). SAC were prepared and stimulated as described above but in the presence or absence of different concentrations of IFNγ for 6 h. (Top panel) RNA was isolated and analyzed for IL-1β mRNA expression by quantitative RT-PCR. (Bottom panel) Cell lysates from a representative experiment were analyzed by western blot for the levels of pro-IL-1β (p31) and active (mature) IL-1β (p17). Results shown are triplicates of a representative experiment. Similar results were obtained in repeat experiments.
Discussion
In this study, we examined the frequency of Th17 and Treg as well as Th17/Treg ratio in the periphery (lymph nodes) and the target organ (the joints) in AA-susceptible LEW rats and AA-resistant WKY rats following a potentially arthritogenic challenge (Mtb injection). We observed significant differences in these immune parameters in the two rat strains. The frequency of Th17 and Th17/Treg ratio were significantly higher in the joints of LEW rats than that of the WKY rats. Although the frequency of Th17 showed an increase in the periphery (lymph node) with the progression of disease in LEW rats, it was not different from WKY rats at any time post-Mtb injection. In contrast, Treg frequency in the lymph nodes of WKY rats was significantly (P <0.05) higher than that in LEW rats at different phases of AA. A similar trend was observed for the joint-infiltrating cells but the difference did not reach statistical significance. This difference in Treg frequency in the draining lymph nodes, which are the primary site of pathogenic T cell activation in response to Mtb challenge prior to migration of these cells into the joints, suggests that the increased levels of Treg in WKY rats (compared with LEW rats) might help attenuate the pathogenic activity and/or subsequent migration of Th17 into the joints of this rat strain. In regard to Th17 cells, our analysis in AA revealed two subsets, one producing IL-17 and the other producing both IL-17 and IFNγ (double producers or double positive cells). The latter subset of T cells has also been reported in other autoimmune diseases.24–26 It has been proposed that IL-17+ IFNγ+ T cells are more pathogenic than IL-17+ T cells for inducing tissue inflammation. 26 However, another perspective on IL-17+ IFNγ+ T cell subset relates to spontaneous downregulation of inflammation. With the progression of inflammation induced by IL-17, there is gradual induction of IFNγ, an inhibitor of IL-17, to control ongoing inflammation. 26 Additional studies in different autoimmune diseases/models are warranted to further clarify the role of IL-17+ IFNγ+ T cells in the disease process.
As mentioned above, upon examining the frequency of Th17 in LEW vs. WKY rats, we observed comparable levels in the lymph nodes but marked difference in the joints (LEW>>WKY). These results suggest that there might be differential migration and retention of Th17 in the joints in these two rat strains. However, the precise factors influencing the differential migration and retention of pathogenic Th17 cells into the joints of LEW vs. WKY rats are not fully defined. In this context, increased production of chemokines such as RANTES are shown to induce chemotaxis of CD4+ T cells to the inflamed joints, 27 and elevated levels of chemokines in the serum and synovial fluid of arthritic LEW rats have been observed in a previous study, but that study did not involve WKY rats. 21 In the present study, we compared the serum RANTES and MCP-1 levels in LEW and WKY rats. As expected, LEW rats showed significantly higher levels of both RANTES and MCP-1 than WKY rats. These results suggest that differential chemokine production by the two rat strains following Mtb injection might be one of the factors influencing T cell migration into the joints of LEW rats, and thereby contribute to the difference in AA-susceptibility of the two rat strains.
To further understand the differential AA-susceptibility of LEW and WKY rats, we tested the serum levels of pro-inflammatory cytokines IL-1β and IL-17, which play an important role in arthritis pathogenesis,12,13 at different time points post-Mtb injection. Serum level of IL-1β was significantly reduced in WKY compared to that of LEW rats, whereas IL-17 levels were comparable in the two rat strains. The observed lack of difference in serum IL-17 level is reinforced by the results of our previous study showing that IL-17 response in the draining lymph node cells of Mtb-immunized LEW and WKY was comparable. 14 Therefore, we focused our attention to IL-1β. To evaluate the reasons for the difference in serum levels (in vivo) of IL-1β in Mtb-immunized LEW and WKY rats, we undertook two approaches: (1) testing the in vitro production of IL-1β; and (2) cytokine (IFN-γ)-based regulation of IL-1β. We first examined whether WKY rats might be defective in IL-1β production. For this purpose, we tested the production of IL-1β in antigen-presenting cells (using spleen adherent cells; SAC) of LEW and WKY rats. Pro-IL-1β is known to be produced through the activation of NF-kB and this precursor form of the cytokine is then converted to active IL-1β by caspase-1 upon activation by inflammasome. 17,18 We observed no difference in the production of IL-1β and the expression of inflammasome-related molecules NLRP3 and caspase-1 in SAC of LEW and WKY rats. These findings suggest that antigen-presenting cells (APCs) of LEW and WKY rats behave similarly to Mtb exposure in vitro. In this regard, the observed difference in serum levels (in vivo) of IL-1β in LEW vs. WKY rats could be because of in vivo regulation as discussed below.
In our quest to further explain the reduced serum level of IL-1β in WKY rats compared to LEW rats, we considered the responses of the two rat strains for other cytokines. We have previously shown that IFNγ levels are much higher in WKY than in LEW rat following Mtb injection. 15 Previous studies by others and us have described the Th17-inhibitory activity of IFNγ.14,28 In this study, we highlighted another arm of IFNγ-mediated events, namely IFNγ-driven inhibition of IL-1β response. A difference in IFNγ production by LEW and WKY rats might be one of the reasons for reduced levels of IL-1β in the serum of WKY. Thus, the development of AA, or the resistance to AA, following Mtb injection is most likely a result of a difference in cytokine (IL-1β) regulation and not because of a defect in IL-1β production. Our conclusion regarding IFNγ-mediated inhibition of IL-1β is based on using SAC. However, considering the significance of synovial-infiltrating cells and the draining lymph node cells in the pathogenesis of AA, it would be pertinent to examine these cells as well besides testing SAC as described above. We plan to undertake these experiments in our ongoing studies in LEW and WKY rats.
Previous studies by other investigators have elaborated upon different factors that influence susceptibility/resistance to autoimmunity. For example, the role of specific non-MHC genes such as Th1-type cytokines and chemokines in disease susceptibility/resistance has been highlighted in experimental model of uveitis. 29 Similarly, Th1 and Th17 responses have been shown to be associated with susceptibility/resistance to experimental autoimmune encephalomyelitis (EAE). 30 Other studies in EAE have described the role of regulatory B and T cells; 31 of the influence of antibodies to the autoantigen (myelin); 32 of the polarization of antigen-presenting cells to the inflammatory phenotype; 33 and of the nuclear IkB family protein IkBNS 34 in susceptibility/resistance to that disease. Similarly, differences in antigen processing of specific autoantigenic determinant were shown to affect susceptibility/resistance to type 1 diabetes in mice. 35 In another study, nucleotide polymorphisms at the TNFAIP3/A20 locus have been reported to be associated with inflammatory and autoimmune diseases such as systemic lupus erythematosus. 36 Our present study based on the AA model has unveiled two additional important factors that influence susceptibility/resistance to arthritis, namely Th17/Treg balance and IFNγ-mediated regulation of IL-1β. Taken together, it is apparent that distinct factors affect the outcome of the disease-inducing stimuli in different diseases, both animal models and patients with autoimmunity. Understanding of particular factors in a given disease can help develop disease-specific and novel interventions for therapeutic purposes.
Footnotes
Acknowledgements
We thank Brian Astry, Joseph Stains, Kari-Ann Shirey, Ferenc Livak, Eric Wang, and Lisa Hester for helpful discussions.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported in part by the National Institutes of Health (NIH), USA grant no. R01AT004321).