
Abstract
Objective
To better understand the effects of simvastatin (SS) on the expression and secretion of two chemokines, CXCL10 and CX3CL1, by osteoblasts, and to test whether inhibition of isoprenoid intermediates of cholesterol biosynthesis were involved in the effects of SS.
Methods
Human osteoblasts were incubated in the presence or absence of the inflammatory cytokines tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ), with and without SS (0.1–100 μM). Culture supernatants were then collected, and expression of CXCL10 and CX3CL1 mRNA in osteoblasts was examined using quantitative TaqMan real-time polymerase chain reaction. The levels of CXCL10 and CX3CL1 were measured using enzyme-linked immunosorbent assays.
Results
At a high concentration (100 μM), SS inhibited expression and secretion of the chemokines and showed cytotoxity, whereas at lower concentrations (0.1–1 μM) SS stimulated the expression and secretion of the chemokines. Expression and secretion of CXCL10 or CX3CL1 from osteoblasts were induced by stimulation with TNF-α and IFN-γ. In addition, SS exerted a biphasic effect on the evoked induction of CXCL10 and CX3CL1. Chemokine expression and secretion was also assayed in the presence of mevalonate (MEV), geranylgeranyl pyrophosphate (GGPP) or farnesyl pyrophosphate (FPP). MEV abolished both the inhibitory effect of high-dose SS and the stimulatory effect of low-dose SS. On the other hand, GGPP abolished only the inhibitory effects of high-dose SS, and FPP had no effect at all.
Conclusions
These findings suggest that osteoblasts are an important cellular source of CXCL10 and CX3CL1, and that statins such as SS may modulate the inflammatory process in bone tissues to inhibit bone resorption and stimulate bone formation through biphasic modulation chemokine synthesis.
Introduction
Chemokines and proinflammatory cytokines appear to play key roles in a variety of immune/inflammatory responses.1,2 The maintenance of skeletal homeostasis is a dynamic process driven by the coordinated cellular activities of osteoblasts (OBs), osteocytes and osteoclasts. 3 Moreover, cells involved in bone formation (e.g. OBs) express a variety of chemokines that likely play key roles during bone remodeling under both normal and pathological conditions. 4 For example, we and others have shown that CX3CL1 (fractalkine) and CXCL10 (interferon-inducible protein 10) are expressed and secreted by OBs.5,7
Inhibitors of 3-hydroxy-3-methylglutaryl coenzyme A (HMG CoA) reductase inhibitors (statins) are widely used for lowering serum cholesterol concentrations, and provide an important and effective approach to the treatment of hyperlipidemia and arteriosclerosis. 8 In addition, several recent studies have shown that statins such as simvastatin (SS) also affect inflammation and immune function by modulating the expression and function of inflammatory molecules, including cytokines and metaloproteinase expressed by endothelial cells and macrophages.9,11 Furthermore, it was recently reported that statins modulate the inflammatory process in bone tissues and osteogenic cells, thereby inhibiting bone resorption, stimulating bone formation, and modulating angiogenic factor synthesis.12,15 Since osteoblast-derived chemokines have important roles in during bone remodeling under both normal and pathological conditions, in the present study we examined the expression and secretion of CXCL10 and CX3CL1 by OBs and their modulation by SS.
Materials and Methods
Reagents
Complete medium consisted of Dulbecco's modified Eagle's medium (DMEM; Nissui Pharmaceutical Co., Tokyo, Japan) supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin and 10% heat-inactivated fetal bovine serum (FBS; Gibco Laboratories, Grand Island, NY). Tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ) were purchased from Genzyme/Techne (Cambridge, MA). SS was kindly provided by Merck &Co., Inc, Whitehouse Station, NJ. For experimentation, SS was dissolved in ethanol and converted to the open acid form. To test whether inhibition of isoprenoid intermediates of cholesterol biosynthesis is involved in the effect of SS, chemokine secretion was also examined in the presence of mevalonate (MEV; 100 μM, Sigma Aldrich, St. Louis, MO), geranylgeranyl pyrophosphate (GGPP; 10 μM; Sigma-Aldrich) and farnesyl pyrophosphate (FPP; 10 μM; Sigma-Aldrich).
Preparation of Human Osteoblasts
Human OBs were purified from metaphyseal trabecular bones collected from proximal femora of post-traumatic patients (
Evaluation of Cytotoxicity
The effect of SS on cell viability was evaluated based on trypan blue exclusion and release of lactate dehydrogenase (LDH). For trypan blue assays, the indicated concentrations of SS were added to OBs, after which the cells were then incubated for up to 24 h, detached using trypsin, and resuspended in 1 ml of DMEM. Trypan blue solution (Gibco, Grand Island, NY, USA) was then added to a final concentration of 0.2% and incubated for 1 min. Cells positive for trypan blue were counted using a hemocytometer under a light microscope (Nikon) and were considered dead. In addition, the amount of LDH released into the culture medium was determined using an automatic autoanalyzer system (Hitachi, Tokyo, Japan).
Enzyme-Linked Immunosorbent Assays
CXCL10 and CX3CL1 were quantified using double ligand enzyme-linked immunosorbent assays (ELISAs) that were modifications of an assay described previously.17,18 Monoclonal murine anti-human CXCL10 (1 μg/ml; Genzyme/Techne, Cambridge, MA) or CX3CL1 (4 μg/ml; Genzyme/Techne) served as the primary Ab, and biotinylated polyclonal goat anti-CXCL10 (0.1 μg/ml; Genzyme/Techne) and anti-CX3CL1 (0.25 μg/ml; Genzyme/Techne) served as the respective secondary Abs. The sensitivity limit for the CXCL10 and CX3CL1 ELISAs were ∼50 pg/ml and ∼150 pg/ml, respectively.
Isolation of Total RNA and Real-Time Polymerase Chain Reaction (PCR)
Total RNA was extracted from human OBs using TRIzol reagent (Invitrogen, Carlsbad, CA), after which 1-μg samples were reverse transcribed into cDNA by incubation with TaqMan RT reagents (Applied Biosystems, Foster City, CA), first for 120 min at 37 °C and then for 10 min at 25 °C. Real-time PCR was carried out using an ABI Prism® 7900 sequence detection system (Applied Biosystems). The PCR mixture included 40 ng of cDNA, which was amplified using AmpliTaq Gold DNA polymerase (Applied Biosystems) as described previously. 16 Appropriate Assays-on-Demand™ primers and probes (Applied Biosystems) were used for detection of CXCL10, CX3CL1 and ribosomal RNA (rRNA) expression. For quantification, the number of target mRNA copies per rRNA copy was calculated, and values were expressed as fold increases over control (culture medium alone).
Statistical Analysis
Data are expressed as means ± SEM. Groups were compared using analysis of variance, after which the means of groups whose variances were determined to significantly differ were compared using Student's
Results
Effects of SS on Cell Viability
We, first, carried out trypan blue dye exclusion assays and measured LDH activity in culture supernatants to determine the degree to which cytotoxicity contributed to the inhibitory effects of higher concentrations of SS. After stimulating OBs with IFN-γ (1000 IU/ml) + TNF-α (20 ng/ml), treatment with SS at concentrations ranging from 0.1–10 μM elicited no significant changes in the numbers of viable (trypan blue negative) cells or LDH activity, though small increases and decreases were noted (Table 1). At a concentration of 100 μM, however, SS significantly reduced the numbers of viable cells and increased release of LDH.
Effect of SS on cell viability of OBs.

OBs (1 x 105 cells/ml) were stimulated with TNF-α (20 ng/ml) plus IFN-γ (1000 u/ml) for 24 h, with and without the indicated doses of SS, after which trypan blue exclusion and LDH release were assayed. Also examined were the effects of MEV, GGPP and FPP. To determine OB-derived total LDH, the cells were lysed by addition of the same volume of complete medium, followed by three freeze/thaw cycles. LDH concentrations were then determined to be 286.0 ± 24.5 IU/I. Three separate experiments were performed using three different OB samples. Data are expressed as the mean ± SEM.
Effects of Simvastatin on Expression and Secretion of OB-Derived CXCL10 and CX3CL1
We next determined whether secretion of CXCL10 and CX3CL1 from OBs could be induced by the inflammatory cytokines TNF-α and IFN-γ. We found that high-dose (1000 IU/ml) of IFN-γ induced secretion of small amounts of CXCL10 or CX3CL1 from OBs, as compared with medium alone, but TNF-α had no effect. On the other hand, when applied together, the two cytokines appeared to act synergistically to induce significant secretion of both CXCL10 and CX3CL1 from OBs (Figs. 1 and 2), which is consistent with earlier findings by us and others.7,19
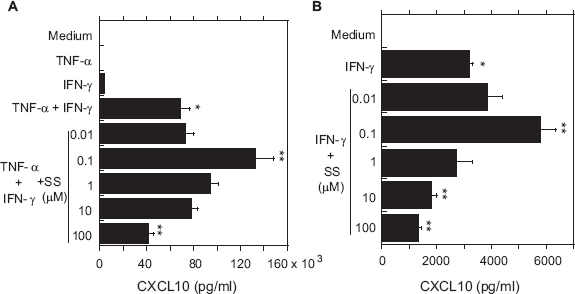
Effects of SS on secretion of CXCL10 from OBs stimulated with TNF-α and/or IFN-γ.
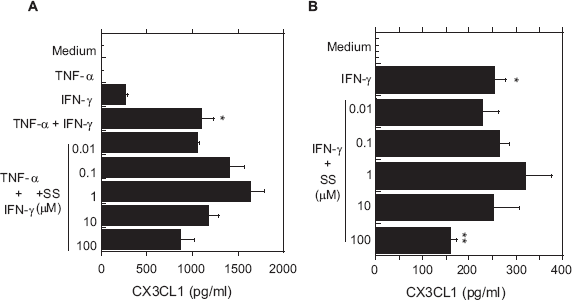
Effects of SS on secretion of CX3CL1 from OBs stimulated with TNF-α and/or IFN-γ.
Next, OBs stimulated with IFN-γ+TNF-α were incubated with various concentrations of SS, and CXCL10 and CX3CL1 in the culture supernatants were assayed. As shown in Figure 1A, a low concentration of SS (0.1 μM) significantly enhanced secretion of CXCL10 from OBs stimulated with IFN-γ+ TNF-α. At a higher SS concentration (100 μM), however, the cytokine-induced secretion of CXCL10 was significantly inhibited. In addition, similar induction (0.1μM SS) and inhibition (10 and 100 μM SS) of CXCL10 were seen in IFN-γ-stimulated OBs (Fig. 1B). SS also exerted this biphasic effect on CX3CL1 secretion from OBs stimulated by IFN-γ + TNF-α (Fig. 2A) or IFN-γ alone (Fig. 2B), though only the inhibitory effect of 100 μM IFN-γ was statistically significant.
To test whether the inhibitory effects of SS on secreted CXCL10 and CX3CL1 antigen levels were accompanied by corresponding changes in the levels of CXCL10 and CX3CL1 mRNA in OBs, we used real-time PCR to assess levels of chemokine mRNA. As shown in Figure 3, the expression of both CXCL10 and CX3CL1 mRNA was enhanced in OBs by lower concentrations of SS (0.1–1 μM), though only the effect on CXCL10 was significant. Higher concentrations of SS (10–100 μM) significantly inhibited expression of CXCL10 and CX3CL1 mRNA.
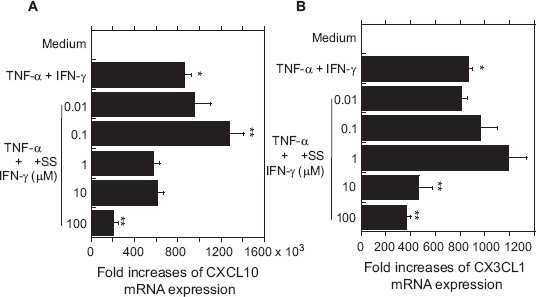
Effects of SS on cytokine-induced expression of CXCL10 and CX3CL1 mRNA in OBs.
Effects of MEV, GGPP and FPP on the Regulation of Chemokine Expression
To assess the extent to which intermediates created during cholesterol biosynthesis were involved in the observed effects of SS, chemokine expression and secretion were also examined in the presence of MEV (100 μM), GGPP (10 μM) or FPP (10 μM), three inhibitors of different steps in the cholesterol biosynthetic pathway. MEV abolished the inhibitory effect of high-dose (100 μM) SS and the stimulatory effect of low-dose (0.1 μM) SS on cytokine (TNF-α + IFN-γ)-evoked expression and secretion of CXCL10 (Figs. 4A and B). MEV had similar effects on the expression and secretion of CX3CL1, though some of the changes were not statistically significant (Figs. 5A and B). Notably, GGPP had no significant effect on the stimulation of CXCL10 expression secretion by low-dose SS; however, it abolished the inhibitory effect of high-dose SS (Figs. 4A and B). In contrast to MEV or GGPP, FPP had no effects on chemokine expression or secretion at any dose. Finally, the reduction in cell viability induced by high-dose SS was abolished by MEV and GGPP, but not by FPP (Table 1).
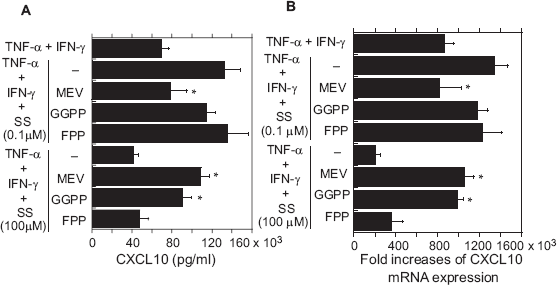
Effects of MEV, GGPP and FPP on CXCL10 secretion (A) and mRNA expression (B) in cytokine-stimulated OBs treated with SS.
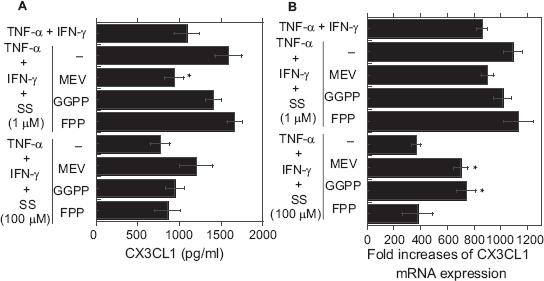
Effects of MEV, GGPP and FPP on CX3CL1 secretion (A) and mRNA expression (B) ftom cytokine-stimulated OBs treated with SS.
Discussion
The results of the present study suggest that OBs are prominent cellular sources of both CXCL10 and CX3CL1, which are secreted in response to stimulation by TNF-αand/or IFN-γ. Furthermore, SS exerted a biphasic effect on OB-derived CXCL10 and CX3CL1 synthesis: at higher concentrations, SS inhibited the expression and secretion of the chemokines and exerting a cytotoxic effect.
In particular, the negative effect of 100 μM SS on the mRNA expression of both chemokines seemed to be related to the inhibition of chemokine gene specific regulation, although the decreased secretion of both chemokines was, in part, dependent upon the drug's cytotoxicity at this high-dose of SS (100 μM). While at lower concentrations it stimulated both the expression and secretion of the chemokines.
Consistent with our present results, it was recently reported that statins exert a dose-dependent, biphasic effect on cultured ECs and angiogenesis.20,21 In addition, 10 μM SS significantly increased the secretion of IL-1β from macrophages, 22 while 0.1–10 μM SS increased expression of vascular endothelial growth factor (VEGF) by osteoblastic cells (MC3T3-E1 cells) and induced osteoblastic differentiation of untransformed MC3T3-E1 osteoblasts.14,23 On the other hand, 1 μM SS inhibited secretion of IL-6 and IL-8 from IL-1α-stimulated epithelial cells, 24 5–50 μM SS inhibited secretion of matrix metalloproteinases from IL-1β-stimulated human chondrocytes, 11 and 0.1–10 μM SS inhibited secretion of IL-6 or IL-8 from TNF-α-stimulated fibroblast-like synoviocytes. 25 These inhibitory effects of statins appear to be dependent on cytotoxic effects of high-dose SS, 26 which is consistent with our finding that high-dose SS reduces synthesis of OB-derived chemokines by exerting a cytotoxic effect.
The OB cytotoxity, and thus the inhibitory effects, of high-dose SS was reversed by MEV or GGPP, but not FPP, while the stimulatory effects of low-dose SS were reversed by MEV, but not by GGPP or FPP. That MEV reversed the biphasic effect of SS on chemokine expression in OBs confirms the involvement of the cholesterol synthesis pathway in the modulation of chemokine expression. Moreover, our finding that GGPP reversed the inhibitory effect of high-dose SS on chemokine synthesis, but not the stimulatory effect of low-dose SS, suggests the latter is not dependent on the membrane anchorage of small G proteins (e.g. Rho) or the Gγ subunit of heterotrimeric G proteins, as both are regulated by GGPP prenylation. 27 In addition, our finding that FPP had no effects indicates the biphasic response to SS is not mediated via farnesylation. It has been shown elsewhere that geranylgeranylated and farnesylated Rho family GTPases are involved in regulating cellular functions, including the synthesis of some cytokines and chemokines.28,29 It would be interesting to explore the signal transduction pathway via which SS modulates the induction of inflammatory proteins in OBs.
There is also a discrepancy between our findings and those of Wu et al, who reported that alkaline phosphatase activity and calcific nodule formation were stimulated by 0.1–0.4 μM SS in an osteoblast cell line (M2–10B4 cells), although these concentrations of SS inhibited both calcific nodule formation and alkaline phosphatase activity in aortic valve myofibroblasts.
30
The lowest concentration of SS (0.1 μM) used in the present study is similar to the serum levels measured in patients' receiving regular doses of SS for therapy (usually less than 90 nM).31,32 Given our present results and the earlier findings of others, one might expect a clinical dose of SS to increase chemokine secretion from OBs at sites of bone inflammation, though it remains unclear whether higher doses of SS are cytotoxic to bone tissues
In conclusion, several recent investigations, as well as our present study, have noted that statins such as SS modulate the inflammatory process in bone tissues, thereby inhibiting bone resorption, stimulating bone formation, and biphasically modulating chemokine synthesis. A more complete understanding of the mechanisms by which statins affect OB-derived chemokine (e.g. CXCL10 and CX3CL1) synthesis under pathological conditions such as rheumatoid arthritis, as well as under normal conditions, should provide new strategies for clinical intervention.
Conflict of Interest Statement
T. Kasama received research grants from Banyu Pharmaceuticals Inc.