
Abstract
Ferrochelatase (FECH) activity is decreased in erythropoietic protoporphyria (EPP), causing increased production and excretion of protoporphyrin. This study examined whether the level of expression of the nonmutant FECH allele is a determinant of phenotype in a mouse model of EPP that carries a heterozygous deletion of exon 10 in FECH. Two mice strains that had a two-fold difference in FECH mRNA levels in bone marrow and liver (low expressing C3H/HeJ and high expressing CBA/J) were used to establish congenic strains containing the mutation. Erythrocyte protoporphyrin levels in C3H/HeJ heterozygous mice were significantly higher than in their wildtype littermates, whereas levels in CBA/J heterozygous mice did not differ significantly from their wildtype littermates. Biliary excretion of protoporphyrin was also significantly higher in C3H/HeJ heterozygous mice. The levels of normal FECH mRNA in bone marrow measured by real time PCR were 138 +/– 30 copies per ug total RNA in C3H/HeJ +/– mice, 320 +/– 59 in C3H/HeJ +/+ mice and 634 +/– 38 in CBA/J +/+ mice. Levels in liver tissue of the mice differed significantly in the same pattern. Thus, the level of expression of the nonmutant FECH allele is a determinant of phenotype in a mouse model of EPP as has been demonstrated in human EPP.
Introduction
Erythropoietic protoporphyria (EPP) is a genetic disorder of porphyrin metabolism in humans that was first described in 1961 by Magness and co-workers when they reported a man with lifelong sensitivity to sunlight and increased protoporphyrin levels in erythrocytes and feces (Magness et al. 1961). Photosensitivity, which is the major clinical feature, results from the photoactive damage of protoporphyrin to skin (Timonen et al. 2000). Some patients also develop hepatobiliary disease due to protoporphyrin induced damage to liver structure and function (Doss and Frank, 1989). This may progress to liver failure and necessitate liver transplantation (McGuire et al. 2005).
A deficiency of ferrochelatase (FECH, EC4.99.1.1) activity underlies the excessive production and excretion of protoporphyrin in EPP (Bonkovsky et al. 1975). FECH, which is the last enzyme in the heme biosynthesis pathway, is located on the matrix side of the inner mitochondrial membrane and catalyzes the insertion of ferrous iron into protoporphyrin to form heme (Dailey et al. 2000). All heme forming tissues may potentially contribute to the excess formation of protoporphyrin in EPP, with the bone marrow being the major source (Poh-Fitzpatrick, 1985). The functional human FECH enzyme is a homodimer that contains two NO sensitive and coordinated 2Fe-2S clusters (Wu et al. 2001). The FECH gene contains 11 exons, and FECH mRNA has an open reading frame of 1269 bases that encodes a protein of 423 amino acid residues (Nakahashi et al. 1990). There is a single transcript with 2 polyadenylation sites in erythroid and non-erythroid cells, indicating there is only one FECH gene in heme forming tissues (Taketani et al. 1992).
Patients with EPP are usually heterozygous for a FECH mutation, even those with severe disease, and extensive genetic heterogeneity has been noted (Wang et al. 1994; Rufenacht et al. 1998). The gene mutations by themselves do not account for disease expression, as they cause only minimally increased protoporphyrin overproduction (Chen et al. 2002). Thus most individuals with symptomatic disease also have a polymorphism in intron 3 of the nonmutant FECH allele (IVS3-48c) that lowers gene expression (Gouya et al. 2002; Risheg et al. 2003).
The purpose of this study was to examine whether the level of expression of the nonmutant FECH allele is also a determinant of phenotype in a mouse model of EPP, using mice in which an exon 10 deletion of FECH had been introduced into the genome of mouse embryonic stem cells by homologous recombination (Magness and Brenner, 1999; Magness et al. 2002).
Methods
Selection and breeding of Mice
Magness and Brenner provided mice carrying the exon 10 deletion, along with a neomycin resistant cassette in the same allele (Magness et al. 2002). Primers were designed to hybridize with this cassette, and PCR amplification of genomic DNA from animals with the exon 10 deletion produced a product that was identified by ethidium bromide stain on agarose gel, whereas genomic DNA from wild-type animals did not produce the product. Wild-type strains were selected on the basis of FECH gene expression as assessed by the level of FECH mRNA in liver and bone marrow, using relative quantitative RT-PCR as previously described (Risheg et al. 2003). Mice from five different strains were evaluated (Table 1). Based on the results, CBA/J mice were selected as a high expressing strain and C3H/HeJ mice as a low expressing strain. FECH cDNA and 1000 bases of the FECH gene promoter and 5’ untranslated region of the two mice strains were sequenced, and no differences were found. Intron 3 was also sequenced, and both strains had base t at position -48. Thus it is likely that the difference in FECH expression in the two strains is caused by transacting factors and/or modifier genes, not by a difference in FECH gene sequence.
FECH mRNA in different mice strains.
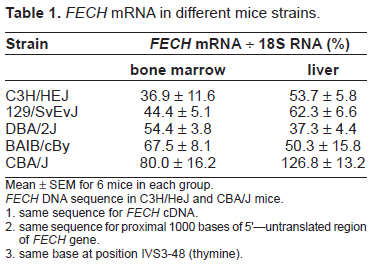
Mean ± SEM for 6 mice in each group.
FECH DNA sequence in C3H/HeJ and CBA/J mice.
same sequence for FECH cDNA. same sequence for proximal 1000 bases of 5'–-untranslated region of FECH gene. same base at position IVS3-48 (thymine).
In order to produce congenic strains, male offspring carrying the exon 10 deletion were back-crossed with wild-type female C3H/HeJ and CBA/J mice through 7–9 generations. Breeding of the mice was carried out in the University of Alabama at Birmingham Genetic Engineered Mouse Breeding Facility. The biochemical phenotype of heterozygous adult mice (3–6 months old) and their wild-type littermates was assessed by measurement of 1) erythrocyte protoporphyrin level; 2) total porphyrin level and fluorescence peak in bile; 3) FECH enzyme activity in liver. The normal FECH mRNA level in bone marrow and liver was measured by quantitative real time PCR. Statistical analysis was done by the students T test and Wilcoxin/Kruskal-Wallis test.
Biochemical measurements
Protoporphyrin levels in erythrocytes were measured by spectroflourometry after solvent partitioning (Morton et al. 1988; Bloomer et al. 1998). The level and fluorescence peak of porphyrin in bile was assessed by spectroflourometry after dissolving the bile in perchloric acid methanol (Morton et al. 1988; Bloomer et al. 1998). Bile was obtained from gallbladders of the mice at the time of sacrifice.
The level of FECH activity in whole liver homogenates was measured by Zn-deuteroporphyrin formation in pmol per second per mg protein (Bloomer et al. 1998).
Quantitative measurement of FECH mRNA by real-time PCR
The level of FECH mRNA was measured in total RNA extracted from whole liver tissue and from bone marrow of leg bones of the adult mice. Only the level of the wild-type (normal) species of FECH mRNA was measured in wild-type mice and mice heterozygous for the exon 10 deletion in FECH gene. The mutant species of FECH mRNA was not measured. The method used the Quantitect™ custom assay developed by Qiagen (Valencia, CA).
In order to develop the standard curve for the assay, RNA was extracted from mouse liver, and a 484 bp FECH amplicon (from exon 7 to exon 11) was generated by RT-PCR. The primer sequences were sense, TCATCCAGTGCTTTGCAGAC; antisense, AGCTTGTTGGACTGGATGTG. The amplicon was purified on silica column (QIA quick PCR purification, Qiagen) and cloned into pCRS.1 vector (Invitrogen). Ligated fragments were placed in DH5a competent cells (Life Technologies). Plasmid DNA was prepared and the cloned amplicons were sequenced; cDNA plasmid concentrations were measured by optical density spectrophotometry (Spectronic Genesys 5). Copy number determination was calculated using the following formula: Copies/ml = 6.023 × 10 23 × [C] × OD260/molecular weight Where [C] = 5 × 10–5 g/ml for DNA; Molecular weight of the PCR product = number base pairs × 6.58 × 10 2 . Serial dilutions from the cDNA plasmid were used as standard curves.
Primers and probes were designed by Qiagen Quantiprobe Design Software (Qiagen). The probe was designed to locate in exon 10 for specific detection of wild-type FECH mRNA. The probe sequence was ATCAGAAGAGCGGAGT with fluorophone at the 3’ end and nonfluorescent quencher and minor grove binder (MGB) at the 5’ end. The PCR primers yield a 93-nt amplicon from wild-type FECH with one primer specific to exon 9/10 fusion region and one primer specific to exon 11. Real-time PCR was performed on an ABI prism 7700 sequence detection system (Applied Biosystems). PCR conditions were 95 °C for 15 min, 40 cycles at 94° for 15s, 56 °C for 30s and 76 °C for 30s. Each 50 µl reaction contained 25 µl 2x QuantiTect probe PCR Master Mix, 2.5 µl 20x primer mix, 2.5 µl 20 × Quantiprobe Solution and template cDNA. Each sample was run in triplicate. Standard wells contained 4000, 2000, 1000, 500, 200, 100, 50 and 10 copies of plasmid FECH cDNA.
DNA microarray analysis of bone marrow gene expression
Microarray analysis of bone marrow gene expression in C3H/HeJ mice and CBA/J mice was done using the GeneChip® Mouse Expression Set 430 (Affymetrix, Santa Clara, CA). Bone marrow was obtained from leg bones of 3 adult male mice and pooled as a single sample. Three samples were prepared for the two mice strains, total RNA was isolated by TRIzol reagent (Life Technologies, Rockville, MD) from each of the 6 samples, and 2 ug of total RNA from each sample was submitted to the Microarray Shared Facility for Affymetrix Gene Chip Analysis, University of Alabama at Birmingham Comprehensive Cancer Center.
The details of the procedures used in the analysis are presented in the manufacturer's technical manual (Affymetrix). In brief, the quality of the RNA was determined using the RNA Nano Chip on the Agilent BioAnalyzer before synthesis of double-strand cDNA. Double-strand cDNA was generated by linear amplification using an oligo dT-T7 primer and reverse transcriptase (RT). Biotin labeled cRNA was then synthesized by in vitro transcription (IVT) using the 3‘-Amplification Reagents for IVT labeling (Affymetrix). The quality of the cRNA was determined on the Agilent BioAnalyzer before it was fragmented into 50 to 200 base fragments. Prior to hybridization to the expression arrays, the quality of the hybridization target was determined by hybridization to a Test3 array. The result indicated the efficacy of the RT/IVT reaction by the ratios of expression level of 5’ to 3’ of the housekeeping genes β-actin and GAPDH. If the quality of the hybridization target passed the quality parameters from the Test3 array, the expression arrays were hybridized overnight at 45 °C and then washed, stained, and scanned on the following day. Gene expression levels were extracted using the Gene Chip Operating Software. Comparisons were made of bone marrow gene expression for each of the 3 CBA/J samples with each of the 3 C3H/HeJ samples to determine the fold change for each transcript (9 total comparisons).
For microarray data analysis and annotation, the software packages GeneTraffic (Iobio/Stratage/Agilent, Inc. Santa Clara, CA) and ArrayAssist Enterprise together with PathwayAssist (Stratagene/Agilent, Santa Clara, CA) were used. Briefly, the raw Genechip files from GCOS were uploaded, background-subtracted, and normalized with GC-RMA method (Gentleman et al. 2004). The control group was used as a baseline to calculate the intensity ratio/fold changes of the treated group versus the control group. The ratio was log2-transformed before further statistical analysis. The p-values were obtained by an unpaired t-test assuming unequal variance.
Results
Levels of erythrocyte protoporphyrin in adult (3–6 months old) heterozygous C3H/HeJ mice were significantly higher than in C3H/HeJ wild-type mice, CBA/J heterozygous mice and CBA/J wildtype mice (Table 2). There was no significant difference noted between male and female mice in the same strain. Bile porphyrin levels in the heterozygous C3H/HeJ mice were also significantly increased compared to C3H/HeJ wild-type mice (79 ± 0.1 versus 28 ± 0.1 ug/dl, p = 0.006), and the peak emission fluorescence was at 604 nm, characteristic of protoporphyrin. Thus, there was increased production and excretion of protoporphyrin in the heterozygous C3H/HeJ mice.
Normal FECH mRNA and FECH activity in mice.
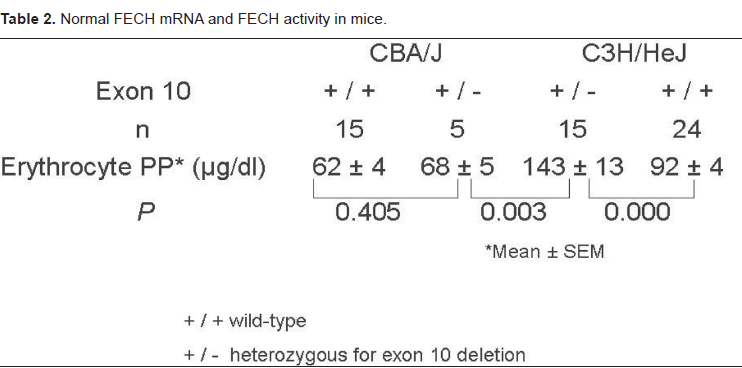
The levels of normal FECH mRNA in bone marrow and liver of wildtype C3H/HeJ mice were approximately 50% of those in CBA/J wildtype mice (Table 3), which agreed well with the relative levels estimated by relative quantitative PCR (Table 1). The lowest levels of normal FECH mRNA were found in C3H/HeJ heterozygous mice, in which bone marrow and liver levels were reduced by 57% and 60% respectively compared to levels in wildtype C3H/HeJ mice.
Normal FECH mRNA and FECH activity in mice.
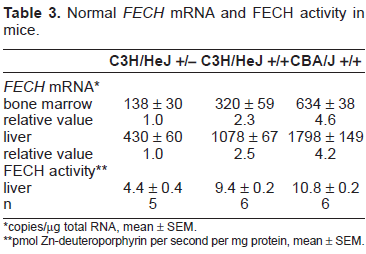
copies/µg total RNA, mean ± SEM.
pmol Zn-deuteroporphyrin per second per mg protein, mean ± SEM.
FECH enzyme activity in livers of heterozygous C3H/HeJ mice was reduced by 53% compared to the activity in wildtype C3H/HeJ mice, in parallel with the reduction in normal liver FECH mRNA. This argues against a significant dominant/negative effect of the mutant FECH protein on the normal protein in mice with an exon 10 deletion. If that were the case, the reduction in FECH enzyme activity should have been approximately 75%.
In order to compare bone marrow expression of other genes in these two mouse strains, and potentially to identify candidate modifier genes of FECH, DNAmicroarray analysis was done in adult male CBA/J and C3H/HeJ mice. This identified 103 genes that were upregulated in CBA/J mice compared to C3H/HeJ mice as defined by at least a 1.5 fold increase in mRNA level (p < 0.05) (Table 4). There were 31 genes which had more than a two-fold change in the level of expression. In contrast there were only 48 bone marrow genes that were upregulated in C3H/HeJ mice compared to the CBA/J mice, six of which had a more than two-fold change in level of expression (Table 5). Thus more bone marrow genes in CBA/J mice are upregulated than in the C3H/HeJ mice. The pathways that were most impacted by these changes were SAPK-JNK signaling, mitrochondrial apoptosis control, integrin signaling, death receptor signaling, caspase signaling, apoptosis, PDGF signaling. The genes encoding other enzymes of the heme biosynthesis pathway showed no significant difference in level of expression between C3H/HeJ mice and CBA/J mice.
Bone marrow genes upregulated in CBA/J mice compared to C3H/HeJ mice (mean fold change >1.5, P < 0.05).

Bone marrow genes upregulated in C3H/HeJ mice compared to CBA/J mice (mean fold change >1.5, P < 0.05).

Discussion
This study demonstrates that mice strains have different levels of FECH mRNA in the major tissues of heme formation, bone marrow and liver, as CBA/J mice had levels approximately two times those found in C3H/HeJ mice (Tables 1 and 3). As a consequence, C3H/HeJ mice heterozygous for an exon 10 deletion in FECH had a significantly higher level of protoporphyrin in erythrocytes and bile than wildtype C3H/HeJ mice (Table 2), with an insignificant increase in heterozygous CBA/J mice compared to wild-type CBA/J mice. Thus, the level of expression of the nonmutant FECH allele is a determinant of biochemical phenotype in this mouse model of EPP.
However, the biochemical abnormality in C3H/HeJ heterozygous mice is mild compared to that in humans with clinically manifest EPP, where erythrocyte protoporphyrin levels are usually several hundred ug/dL, and in patients with severe disease several thousand ug/dL. In most humans with clinically manifest EPP, the level of expression of the nonmutant FECH allele is lowered by a polymorphism in intron 3 (IVS3-48c) that causes increased formation of aberrantly spliced FECH mRNA due to activation of a cryptic acceptor splice site. This causes incorporation of 63 bases of intron 3 into FECH mRNA, and the aberrantly spliced transcript contains a stop codon that causes it to be rapidly degraded by nonsense mediated decay. Thus the polymorphism exerts a more pronounced effect on the expression of the nonmutant FECH allele than occurs due to baseline expression in the mouse model.
Nevertheless, among patients who carry the same mutation and the polymorphism there can be considerable variation in the severity of the phenotype, thus indicating that there are probably additional factors outside the FECH locus that impact phenotype in EPP, in particular transcription factors and modifier genes that might alter the level of expression of FECH. There are several other bone marrow genes in CBA/J mice that are differently regulated than those in C3H/HeJ mice (Tables 4 and 5). Which, if any, of these might be causing a difference in FECH expression cannot be determined at this time, however.
Several other investigators reported that the phenotype of a single mutation in mice was modulated by the genetic background of the strain, which was attributed to modifier genes (Montagutelli, 2000). This includes the ethylnitro-sourea-induced point mutation in FECH that was originally described as causing recessive inheritance of FECH activity in the house mouse (FECHml/Pas/FECH ml/Pas mouse) (Boulechfar et al. 1993). Abitbol et al. examined three congenic strains into which the point mutation had been introduced (Abitol et al. 2005). Compared with the BALB/cByJCrl genetic background in which recessive transmission of the mutation caused severe skin lesions, anemia, jaundice and hepatic dysfunction with massive protoporphyrin deposits, C57BL/6JCrl mice developed anemia and intense liver accumulation of protoporphyrin with hepatocyte damage, but bile excretory function was not affected, and the serum bilirubin remained low. In SJL/JOrlCrl homozygous mice there was a very high protoporphyrin concentration in erythrocytes, but anemia was mild, and there were few hepatic deposits. Navarro et al. used these same three congenic strains to show a difference in the level of mitochondrial respiratory chain enzyme activities and suggested that an increase of these activities provided protection against liver disease in the EPP mice (Navarro et al. 2005). Thus, the study of different congenic mice strains with EPP may provide a means by which to identify modifier genes of phenotype in this disorder, which could help in understanding the reason for different phenotypes in human EPP.