
Abstract
Living eukaryotic systems evolve delicate cellular mechanisms for responding to various environmental signals. Among them, epigenetic machinery (DNA methylation, histone modifications, microRNAs, etc.) is the hub in transducing external stimuli into transcriptional response. Emerging evidence reveals the concept that epigenetic signatures are essential for the proper maintenance of cellular metabolism. On the other hand, the metabolite, a main environmental input, can also influence the processing of epigenetic memory. Here, we summarize the recent research progress in the epigenetic regulation of cellular metabolism and discuss how the dysfunction of epigenetic machineries influences the development of metabolic disorders such as diabetes and obesity; then, we focus on discussing the notion that manipulating metabolites, the fuel of cell metabolism, can function as a strategy for interfering epigenetic machinery and its related disease progression as well.
Epigenetics
In biology, epigenetics is characterized by regulations of gene expression without alteration of the underlying DNA sequence.1,2 In epigenetic manifestations, external or environmental factors (including nutrition, stress, toxins, and medical history) play an important role in regulating the expression of certain genes. 3 Hence, epigenetic investigations aim to understand how external factors regulate gene expression, and even phenotypic traits. Actually, the term epigenetics, with the prefix “epi” taken from the Greek language (meaning outside of, or around), emerged in the 1990s. Until 2008, the formulated definition of epigenetic traits was present as a stably heritable phenotype without changes in DNA sequences at a Cold Spring Harbor meeting. 4 During the past 15 years, plenty of cases and mechanisms are discovered and studied in the field of epigenetics. Among them, three major mechanisms are categorized by biologists to instruct epigenetic regulations (Fig. 1): DNA methylation, histone posttranslational modifications (PTMs), and noncoding RNAs. 2

Mechanisms of epigenetic modifications. Cell transduces the environmental changes (eg, nutrients, toxins, etc.) into epigenetic modifications including DNA methylation, histone PTMs, and noncoding RNAs (especially the microRNAs), which eventually turn certain genes on or off and regulate the transcription process without changing DNA sequence.
DNA methylation
DNA methylation is among the best studied and characterized epigenetic modifications. A methyl group is added to the C-5 position of a cytosine adjacent to a guanine residue (CpG dinucleotides), which normally leads to gene suppression. 2 In mammals, the CpGs are predominately methylated. However, the CpG islands, which locate in the promoter region of housekeeping and developmental regulator genes with dense CG distribution, are largely resistant to DNA methylation. During the progression of diseases, CpGs of some key genes are reported to be abnormally hyper- or hypomethylated, which further result in transcriptional misregulation. 5 It is well demonstrated that in mammalian cells, DNA methylation is performed by DNA methyltransferases DNMT1, DNMT3A, and DNMT3B. Among them, DNMT1 propagates DNA methylation patterns during DNA replication, while 3A and 3B are involved in establishing de novo patterns of DNA methylation during development and cell fate determination.6,7 DNA can also be demethylated through several enzymatic reactions. For instance, the 10 to 11 translocation (TET) proteins can mediate the oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), finally leading to DNA demethylation. 8
Histone PTMs
Chromatin is a macromolecular structure that consists of DNA, protein, and RNA. DNA wraps around histone octamer to form nucleosomes, the fundamental unit of chromatin. Each histone octamer involved in the organization of nucleosomes contains two copies of four histones (H2A, H2B, H3, and H4). The precise structure of a nucleosome has been illustrated, and the functions of these protein–DNA or protein–protein interactions are suggested to be important for many cellular processes.9,10 Since chromatin is usually packed with nucleosomes, which is believed to be a challenge for transcription accessibility, chromatin should be remodeled to allow the access of transcription machinery to DNA. 11 In the past few decades, compelling evidences suggest that histone modifications are able to change chromatin structure and are associated with both transcriptional activation and repression. 12 Therefore, the chromatin features are essential for the molecular biologist to analyze the transcriptional regulations and cellular status.13–17
In general, histone modifications mainly include acetylation, methylation, phosphorylation, and ubiquitination. Recently, some novel modifications are identified by tandem mass spectrometry (MS/MS), such as histone lysine GlcNAcylation, butyrylation, malonylation, and crotonylation.18,19 Among them, acetylation is one of the most widely studied epigenetic modifications that commonly occurs on the basic amino acids (lysine and arginine). Basically, acetylation can neutralize the positive charge of basic residues and reduce the affinity between histones and DNA for gene activation. 19 This process is catalyzed by histone acetyltransferases (HATs). HATs are traditionally divided into two different classes (type A and B) based on their subcellular localization. Type A HATs locate in the nucleus and are involved in the regulation of gene expression through acetylation of nucleosomal histones. Type B HATs locate in the cytoplasm and are responsible for acetylating newly synthesized histones prior to their assembly into nucleosomes.20–22 The acetylation of histones can also be removed by histone deacetyltransferases (HDACs). HDACs are classified into four classes (I, II, III, and IV) according to their functions and DNA sequence similarity. Classes I, II, and IV are considered as “classical” HDACs whose activities are zinc dependent and inhibited by trichostatin A (TSA). Whereas class III enzymes, known as sirtuins, are NAD+ dependent and not affected by TSA.23–25 Histones can also be modified by other acyl moieties, eg, crotonyl, but the mechanisms and the functional consequences of acetylation versus crotonylation are still unclear. Sabari et al 26 indicated that p300, which belongs to the type A HATs, processes both crotonyltransferase and acetyltransferase activities, and the induced transcription by crotonylation is greater than acetylation. These results provide a new clue to clarify the specificities of different histone acylations in regulating metabolic-related gene expressions. Methylation is another widely studied histone modification, which is catalyzed by histone methyltransferases. 27 Methylation can influence gene expression in both positive and negative ways. For example, trimethylation of lysine 4 on H3 (H3K4me3) is strongly associated with gene activation, while trimethylation of lysine 27 on H3 (H3K27me3) is reported to be associated with gene silencing.28,29 Histone demethylation is performed by two classes of histone demethylases: lysine-specific demethylase (LSD) family proteins (LSD1 and LSD2) and JmjC domain-containing histone demethylase (JHDM). 27 The histone PTMs are not independent of each other, some crosstalks exist among them. Recent studies of Li et al indicated that a Serine-responsive SAM-containing Metabolic Enzyme complex, which consists of pyruvate kinase, serine metabolic enzymes, and SAM synthetases, can interact with a H3K4 methyltransferase Set1 and mediate the crosstalk between H3T11 phosphorylation (H3Tp11) and H3K4me3. 30
MicroRNAs
MicroRNAs (miRNAs), the short noncoding RNAs (approximately 22 nucleotides in length), are able to regulate target gene expression via multiple molecular pathways. Normally, miRNAs can interfere with gene expression through their complementary binding to the target mRNAs, which either inhibit translation or cause mRNA degradation.31,32 Currently, miRNAs are identified in almost all metazoan genomes and even in some viruses.
33
Bioinformatic analysis of known miRNAs suggests that they can regulate the large majority of mRNAs and that a single miRNA can also target multiple mRNAs. Recent findings implicate that miRNAs participate in either regulating expression of certain genes or RNA degradation in important cellular processes, such as apoptosis, cell proliferation, differentiation, and metabolism.32,34 Furthermore, enzymes involved in epigenetic modifications can also be governed by miRNAs. It is reported that some miRNAs can directly inhibit the activities of enzymes, which are related to histone acetylation, thereby considered as key regulators during epigenetic cascade. For example,
Epigenetic Regulation on Metabolism
The prevalence of type 2 diabetes (T2D), obesity (OB), and other metabolic diseases has caused wide public attention during past two decades. Increasing studies also reveal that the epigenetic regulation can modulate metabolic dysfunctions in many aspects, which further broadens our understandings about metabolic disease progressions. Here, we discuss the recent research progress about how epigenetic machineries: DNA methylation, histone PTMs, and miRNA regulate the metabolisms.
DNA methylation and metabolic disorders
DNA contains the genetic information of a cell, and methylation of DNA bases can be highly influential on regulating gene activity, including some important genes that are involved in metabolic pathways.37–39 Over the past decade, evidence has been accumulated to define the regulatory role of DNA methylation in metabolic disorders such as diabetes and OB by taking the advantage of molecular genetic dissection and genome-wide association study (GWAS) analysis. 40
The balance of blood glucose is very important for human health, the pancreatic β cells in islets can secrete insulin (INS) in response to the increased blood glucose level to prevent hyperglycemia and inhibit insulin secretion under fasting conditions to avoid hypoglycemia.
41
Studies by Kuroda et al
42
showed that the CpG sites in mouse
The GWAS studies indicate that metabolic diseases (eg, diabetes and OB) are also associated with the alterations of DNA methylation at specific genomic loci such as peroxisome proliferator-activated receptor γ
PTMs and T2D
Histone modifications can regulate gene expression by modifying the structure of chromatins as described. Recently, an increasing number of evidence is accumulated to elucidate the roles of histone PTMs in metabolic diseases, especially in T2D. 49 , 50
For diabetic patients, metabolic memory means inflammation and vascular complications after controlling the blood glucose level, which presents major challenges to treatment.
51
Recent findings suggest that epigenetic mechanisms may be responsible for metabolic memory. Compared with normal cells, trimethylation on H3 lysine 9 (H3K9me3) was significantly reduced in vascular smooth muscle cells (VSMCs) derived from the
MiRNAs and cellular metabolism
MiRNAs are found to be responsible for regulating various biological processes and viewed as an emerging and exciting topic of biomedical research. In recent years, it has been increasingly reported that miRNAs are associated with disease development.56,57 However, the underlining molecular mechanisms are still ambiguous. Here, in order to get a comprehensive understanding of the associations between miRNAs and metabolic disease progression, we collect and summarize the latest progress in miRNA studies (see Table 1).
Involvement of miRNAs in cellular metabolic pathways.

Rottiers and Naar
58
showed that miRNAs are essential for glucose homeostasis. For example,
Metabolites Modulate Epigenetics
Chromatin-dependent gene modifications interact with the cellular metabolism reciprocally. Metabolic-related genes can be regulated by epigenetic modifications, and many metabolites of cell metabolism can serve as essential cofactors for chromatin-modifying enzymes and control the transcription or translation processes. Therefore, the fluctuation of local metabolites' concentrations is considered as novel signaling cues for regulating gene expressions. 61 Here, we take several metabolites from glycolysis and tricarboxylic acid (TCA) cycle as examples to discuss the regulatory role on epigenetics. Moreover, the supplement or depletion of nutrients (through fasting or calorie restriction [CR]), which can also influence epigenetics via manipulating the concentration of metabolites, is also detailed in this part.
Metabolic products and epigenetics
Once glucose enters into cells, the major portion can be converted to acetyl-CoA through glycolytic pathways, along with the decreased NAD+/NADH ratio.
61
NAD+ is a key cofactor in a variety of enzymatic reactions, such as sirtuin-mediated histone deacetylations. Unlike the other HDACs, sirtuins consume NAD+ and release nicotinamide,
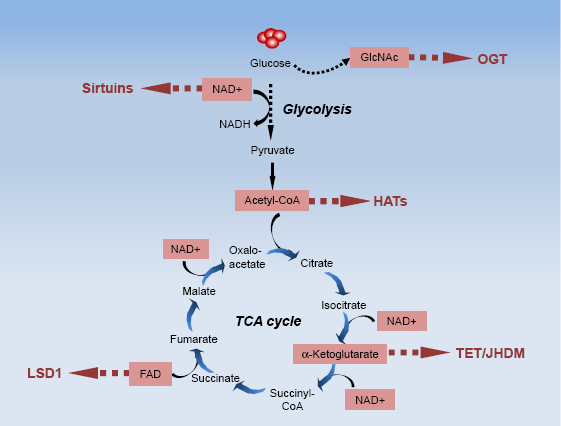
Crosstalk between cell metabolism and epigenetics. As glucose comes into glycolytic reactions, a minor portion involves in hexosamine biosynthetic pathway to produce GlcNAc, which can act as the substrate for histone GlcNAcylation by
Nutrients and epigenetics
Nutrients are essential components in foods that are used by organisms to survive and grow. Among them, most of the carbohydrates, proteins, and fats are the energy supplier for supporting daily metabolism and body activities. While for other nutrient components like vitamin, docosahexoenoic acid (DHA) and dietary fibers are also indispensible for human health. Here, we discuss how these nutrients affect health and disease progression via the epigenetic regulation.65–67
Folate (also called folic acid), a water-soluble vitamin B, is extensively studied for its important function in DNA synthesis, DNA repair, and DNA methylation.
68
Moreover, folate is widely used as food supplementation during pregnancy and infancy to decrease the risk of neural tube defects. Because humans cannot synthesize folates in a de novo manner, folate has to be provided and absorbed from diet to meet demands. Structurally, folate carries a methyl group and can feed into the one-carbon metabolism, in which folate goes through several reactions to generate

Involvement of dietary nutrients in epigenetics. Dietary intake of folate, vitamin B (2, 6, and 12), choline, and methionine (green color) regulate epigenetic modifications through involving one-carbon metabolism where the intermediate SAM is produced (red color) and subsequently be provided as the universal methyl group donor for DNA and histone methylations. One-carbon metabolism is described briefly as follows: folate is first converted to dihydrofolate, then to tetrahydrofolate (THF), which enters the cycle. Vitamin B6 is a cofactor in the conversion of THF to 5,10-methylene THF. The process of 5,10-methylene THF to 5-methyl THF is vitamin B2 dependent. 5-methyl THF serves as a methyl donor in a reaction converting homocysteine to methionine, in which vitamin B12 serves as a precursor to methionine synthase. In turn, methionine generated SAM.
Components of vitamin B family (eg, 2, 6, and 12) and choline can also function as essential cofactors in one-carbon metabolism, which provides the methyl donor for chromatin methylation (Fig. 3).66,67 Severe deficiency of vitamin B12 induces promoter hypomethylation of the cystathionine betasynthase gene and represses its transcription in rats, which cannot be recovered by dietary supplementation of methionine to the B12 deficient rats. The human studies also show a link between vitamin B12 deficiency and the increased risk of various cancers (eg, colon and breast cancers).
73
Choline is essential for fetal neurogenesis such as hippocampal development and memory function throughout life. During mouse embryonic development, choline deprivation caused hypermethylation of CpG3 within the promoter region of
DHA, an omega-3 factty acid, is a primary structural component for human brain, cerebral cortex, sperm, and retina, which has a wide range of effects to human health, including against Alzheimer's disease and cancer.75,76 Sadli et al 77 reported that the addition of DHA to M17 neuroblastoma culture medium showed elevated levels of H3K9ac and reduced levels of HDAC1, 2, and 3, suggesting that DHA promotes gene expression.
Dietary fibers are the indigestible component of food derived from plants, which are helpful for the digestion and absorption of nutrients.78,79 It has been demonstrated that dietary fibers facilitate the generation of short-chain fatty acids (butyrate), and there is a positive linear correlation between butyrate level and histone acetylation in colon epithelial cell. 80 In the last decade, several effects of butyrate are clarified, including anti-inflammatory effects, effects on OB, IR, and inherited disorders.81–84 One of the major functions of butyrate that is involved in the epigenetic regulation of gene expression is inhibiting HDACs.85,86 Specifically, butyrate treatment of cells results in histone hyperacetylation, and butyrate itself inhibits class I HDAC activity in mammalian cells. These findings suggest that dietary fibers are an effective food component that is able to regulate epigenetic modifications.
CR and epigenetics
CR simulates human dieting, which is considered as the most effective environmental manipulation for extending life span so far. This phenomenon has also been experimentally proved in multiple species that range from worm to rodent.
87
CR can induce wide metabolic changes (gluconeogenesis, fat mobilization, etc.) and undergoes weight loss in human subjects. Huang et al found that weight loss is associated with DNA methylation.
88
The DNA methylation pattern in human individuals with normal weight, OB, and successful weight loss maintainers is different, and the pattern in successful weight loss maintainer is more closely resembled than those in normal weight. These findings suggest a relationship between OB, weight loss, and DNA methylation. Moreover, extensive studies reported the crucial role of CR on regulating SIRT1 activities, which is an NAD+-dependent deacetylase. Mechanistically, CR increases NAD+/NADH ratio and induces SIRT1 expression. The upregulated SIRT1 further suppresses the downstream PPARγ, then down-regulates its target gene
Conclusions
Epigenetics influence gene expression with no alterations of DNA sequence and might be inherited by next generation. The major molecular machinery of epigenetics includes DNA methylation, histone posttranslational modifications, and miRNAs. Modifications of chromatin structure ultimately regulate transcription. Increasing evidence demonstrates that metabolic diseases are highly associated with epigenetic alterations. It is well established that metabolic products from cellular metabolic processes like TCA cycle and glycolysis are involved in the regulation of chromatin methylation and acetylation. Moreover, dietary nutrients (such as folate, vitamin B family, DHA, and dietary fibers) play important roles in DNA methylation and histone modifications by directly inhibiting related enzymes or by changing the availability of substrates for those enzymatic reactions.
In the past two decades, scientists have generated a variety of valuable data and knowledge about the basis of epigenetics and cellular metabolism. Here, we discussed the reciprocal regulation between these two phenomena, which eventually leads us to a better understanding of metabolic-related disease progression and pathogenesis. Nevertheless, due to our incomplete knowledge from current studies, we need to make investigations on two key issues. (1) How nutrients and environmental factors are sensed and signaled to the epigenetic machinery? Here, we discussed sparse cases and steps of how nutrients are able to direct epigenetic modifications. However, the detailed molecular mechanisms are still missing for several important signaling pathways. (2) How does the cell response to the metabolic changes at a systematic level? Since the whole cellular network is highly orchestrated and complicated, it is basically impossible to understand the connection between metabolite and epigenetics by focusing on a single pathway. In order to achieve a deeper understanding of these puzzles, there is no doubt that the conventional genetic and cellular analysis should be upgraded and combined with recently developed high-throughput sequencing strategies. In future, we expect more exciting discoveries and methods to thrive and help us understand the molecular regulation of epigenetics and cellular metabolism, which may provide a better way to maintain our health through nutritional modulation besides pharmacotherapies.
Author Contributions
Wrote the first draft of the manuscript: WX. Contributed to the writing of the manuscript: ZY and FX. Jointly developed the structure and arguments for the paper: FW. Made critical revisions and approved final version: ZY and FX. All authors reviewed and approved of the final manuscript.