
Abstract
Babesia microti is an emerging zoonotic protozoan organism that causes “malaria-like” symptoms that can be fatal in immunocompromised people. Owing to lack of specific therapeutic regiment against the disease, we cloned and characterized B. microti lactate dehydrogenase (BmLDH) as a potential molecular drug receptor. The in vitro kinetic properties of BmLDH enzyme was evaluated using nicotinamide adenine dinucleotide (NAD+) as a co-factor and lactate as a substrate. Inhibitory assay was also done using gossypol as BmLDH inhibitor to determine the inhibitory concentration 50 (IC50). The result showed that the 0.99 kbp BmLDH gene codes for a barely soluble 36 kDa protein (332 amino acids) localized in both the cytoplasm and nucleus of the parasite. In vitro enzyme kinetic studies further revealed that BmLDH is an active enzyme with a high catalytic efficiency at optimal pH of 10.2. The Km values of NAD+ and lactate were 8.7 ± 0.57 mM and 99.9 ± 22.33 mM, respectively. The IC50 value for gossypol was 0.345 &mUM, while at 2.5 &mUM, gossypol caused 100% inhibition of BmLDH catalytic activity. These findings, therefore, provide initial evidence that BmLDH could be a potential drug target, although further in vivo studies are needed to validate the practical application of lactate dehydrogenase inhibitors against B. microti infection.
Introduction
Babesia microti is a protozoan organism (Piroplasmida: Apicomplexa) that infects mainly rodents and humans. 1 Through phylogenetic investigations, 2 B. microti has been classified into four subtypes, namely US (R1 and Gray strains), Munich, Kobe, and Hobetsu. Recent studies suggest that B. microti has a wide distribution of wild reservoirs in the United States of America 3 and northeastern Eurasia. 4 The emerging zoonotic importance of B. microti has been widely reported.5–7 The organism has been implicated as a major concern for the safety of blood transfusion supply in USA.8,9 Human co-infection of patients with both B. microti and other zoonotic tick-borne hemoparasites has been reported. 10 Moreover, the parasite causes “malaria-like” symptoms in immunocompetent individuals and can potentially lead to fatal relapsing illness in immunocompromised patients. 11
The importance of B. microti as an emerging zoonotic disease has created an urgent need for innovation of effective chemotherapeutic agents for its treatment. A combination of atovaquone and azithromycin as well as clindamycin and quinine were reported as effective against B. microti. 2 However, recrudescence and resistance against atovaquone and lack of enzymes involved in heme metabolism limits the use of atovaquone and quinoline-derived drugs such as quinine and chloroquine in the treatment of B. microti infections.13,14
Lactate dehydrogenase (LDH) is one of the apicomplexan glycolytic enzymes that catalyze interconversion of pyruvate to lactate. 15 The enzyme plays indispensable role in apicomplexan energy metabolism using NAD+ as a co-factor under anaerobic conditions. 16 The resultant energy generated is used by parasites for their biochemical processes and survival. Available reports indicate that the enzyme is a novel drug target in Plasmodium spp, Toxoplasma gondii, Theileria annulata, and Babesia bovis.15,17–19 For this reason, there is a growing momentum in the synthesis of new LDH inhibitors as potential drugs against the above organisms. 20 However, to the best of our knowledge, there is still paucity of scientific evidence on the suitability of B. microti LDH as a potential drug target. Therefore, the aim of the current study was to clone B. microti lactate dehydrogenase (BmLDH), characterize the protein, and evaluate its in vitro kinetic properties with the view of assessing its potential as a novel drug target against B. microti infection.
Materials and Method
Ethics
Ethical clearance (approval No. 250035) was sought and obtained in accordance with the provision of Article 32–1 of the Regulation of Animal Experiment of Obihiro University of Agriculture and Veterinary Medicine, Japan.
Polymerase chain reaction (PCR), cloning and sequencing
The purified genomic DNA of human isolate of B. microti Gray strain (US type, culture collection catalog No. 30221) was used as template. 21 Oligo nucleotide primers with BamHI restriction site at the forward (5′-CCTCG-GATCCCATTCGTTAAAAGAAGA ATTTC-3′) and Xhol site at the reverse (5′-GGGGCTCGAGTTATAGTTG-GATATCTTTCTGTG-3′) were designed from B. microti strain R1 LDH gene sequence obtained from GenBank (Accession No. CCF72479). 14 Thermocycling was done in a 50 μL reaction volume containing 10 ng genomic DNA, 20 pg primers, 0.2 μM dNTP (Takara, Japan), and 2.5 U ExTaq polymerase (Takara, Japan). The PCR condition included two minutes denaturation followed by 30 cycles of 98°C for 10 seconds, 55°C for 30 seconds, 68°C for 50 seconds, and further extension at 72°C for five minutes. The resultant polymerase chain reaction (PCR) product was electrophoresed, stained with ethidium, and viewed under UV transilluminator. The BmLDH amplicon was extracted and purified from the gel using QIAquick Gel extraction kit (Qiagen, Germany) according to the manufacturer's instruction. The purified BmLDH was cloned into pGEM-T Easy Vector (Promega, USA) and transformed into competent Escherichia coli DH5α (Invitrogen). The resultant pGEM-T Easy-BmLDH was purified using NucleoSpin® Plasmid kit according to the manufacturer's (Macherey-Nagel, Germany) instruction. Sequencing was carried out using M13 forward 5′-TGTA-AAACGACGGCCAGT-3′) and reverse (5′-CAGGAAA-CAGCTATGACC-3′) primers with automated sequencer (ABI Prism 3100 Genetic Analyzer, USA).
Expression and purification of recombinant BmLDH in E. coli BL21
Both BmLDH PCR product and pGEX-6P-1 expression vector were double digested with BamHI (Roche, Germany) and Xhol (Roche) restriction enzymes. The digested BmLDH and pGEX-6P-1 were ligated and transformed into competent E. coli BL21 (Invitrogen, Japan). The nucleotide sequence of the cloned BmLDH was confirmed using pGEX-6P-1 forward and reverse sequencing primers with automated sequencer (ABI Prism 3100 Genetic Analyzer, USA).
The recombinant BmLDH was expressed in E. coli BL21 as glutathione S-transferase fusion protein. The soluble fraction of the recombinant protein was purified as previously described 21 with minor modifications. PreScission™ Protease (GE Healthcare, Sweden) was used to cleave BmLDH from Glutathione-Sepharose™ 4B beads to yield GST-free BmLH for enzyme kinetic assay. The purity and concentration of the purified protein were determined using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Lowry protein assay kit (Thermo Scientific, USA).
Production of anti-BmLDH sera
One milliliter of the recombinant BmLDH (220 μg/mL) was mixed with 1 mL of complete Freund's adjuvant (CFA) in sterile syringe as previously described. 22 Five BALB/c mice (10 weeks old) were immunized with 33 ng of BmLDH through the intra-peritoneal route. Three booster doses of 15 days apart were administered, and sera were collected 10 days after the last dose.
Infection of hamsters with B. microti Gray strain, parasite lysate, and Western blotting
Four female (8 weeks old) specific pathogen-free (SPF) Syrian hamsters (SLC, Japan) were used in this experiment. Prior to parasite inoculation, SPF sera were obtained from all the hamsters. Two hamsters were inoculated with previously stocked B. microti-infected hamster RBC intra-peritoneally as previously described. 23 Observation of parasitemia was done every 2 days by microscopy. At peak (51%) parasitemia (day 13 and 14), the hamsters were sacrificed, and blood was obtained for collection of infected sera and preparation of parasite lysate. Parasite and non-infected RBC lysates were prepared as previously described. 24 Transfer of the protein from the gel to Western blot membrane was carried out as previously described. 21 However, the BmLDH polyclonal sera was diluted at 1:250, and the antibody solution was diluted at 1:2,000. The native BmLDH was detected with color development solution containing 3,3′-diaminobenzidine tetrahydrochloride (DAB) and hydrogen peroxide.
Native protein localization by indirect florescent antibody test (IFAT) and confocal laser microscopy
This was carried out using B. microti Gray strain-infected hamster blood smears (day 8 post-infection) as previously described. 21 The secondary antibody was anti-mouse Alexa-488 secondary diluted 200 times with 4% fetal bovine serum (FBS), and the parasite nucleus was stained with propidium iodide. Parasites were observed using confocal laser scanning microscope (Leicia, Japan).
Enzyme kinetic assay
Optimization of pH and determination of Vmax and Km for lactate and NAD+
The method by Kavanagh and others was used with minor modifications. 16 The oxidation of lactate to pyruvate and subsequent formation of NADH from NAD+ was assayed in 7 mM KCl and 70 mM Tris-HCl buffer. Optimal pH, Km, and Vmax were all determined using 0.5 μg of BmLDH. The amount of NADH produced was measured using DU® 800 spectrophotometer (Beckman Coulter Inc., Fullerton, CA, USA) at 340 nm (ε = 6.2 mM–1 cm–1) and 25°C. Optimal pH for BmLDH catalytic activity was determined using 3 mM NAD+ and 120 mM lactate in varying pH (2–12.2) of the buffer. Initial velocities for the rate of formation of NADH in the first 60 seconds were calculated, and relative percentage activity was determined. The Km and Vmax for NAD+ (co-factor) were determined in a 500 μL reaction volume containing the buffer, 100 mM lactate and 0.05–6.8 mM NAD+. Similarly, Km and Vmax for lactate were assayed in a solution containing the buffer, 2.4 mM NAD+ and 20–500 mM lactate.
BmLDH inhibitory assay
This was carried out according to the methods reported by Bork and colleagues, 19 with gossypol as an inhibitor. Briefly, a 400 mM gossypol stock solution was prepared by dissolving gossypol in dimethyl sulfoxide (DMSO). The percentage inhibition of BmLDH was determined at various concentration of gossypol (9.5–5,000 nM) at 1.2 and 30 mM of NAD+ and lactate, respectively. The final concentration of DMSO in each assay did not exceed 0.2%. The negative control contained 0.2% of DMSO. All the experiments were repeated three times, and the concentration of gossypol that inhibited NADH production by 50% (IC50) was calculated using non-linear regression with GraphPad prism (GraphPad Prism version 5.00 for Windows, GraphPad Software, USA).
Bioinformatic analysis
The BmLDH nucleotide sequences of both pGEM-T Easy-BmLDH and pGEX-6P-1-BmLDH clones were edited using CodonCode Aligner version 1.5.2 (CodonCode Corporation, USA) and consensus sequence derived using Genetyx software (Genetyx Corporation, Japan). The consensus nucleotide sequences for full-length BmLDH were aligned using BLAST (http://blast.ncbi.nlm.nih.gov/) to determine the homology. Prediction of importin a-dependent nuclear localization signals was done using cytoplasmic-nucleus localization signal predictor (cNLS) mapper so as to account for localization of the native protein based on its amino acid sequence (http://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi).
Results
Characterization of BmLDH gene
The amplified BmLDH gene was approximately 0.99 kbp. Nucleotide sequence analysis revealed that BmLDH gene has an open reading frame of 996 nucleotides that codes for 332 amino acids. There were no introns in the BmLDH DNA. Alignment of the translated BmLDH amino acid sequence with that of related apicomplexan parasites and selected mammalian LDH gave contrasting result. The amino acid sequence for BmLDH was less than 28% identical to that of Plasmodium falciparum (Accession No. AAK12097), T. gondii (Accession No. XP_002368488), B. bovis (Accession No. EDO07479), and T. annulata (Accession No. ADG45564). However, there was high frequency of identical amino acid sequence between BmLDH and LDH mammals like hamster (70.3%, Accession No. NP_001230979), mouse (69.1%, Accession No. CAA26360), and humans (61.9%, Accession No. CAE11711). Approximately 43 amino acid residues were highly conserved in all the eight LDH sequences aligned (Fig. 1).

Multiple amino acid sequence alignment and identity score between BmLDH and LDH for P. falciparum (PfLDH), B. bovis (BbLDH), T. gondii (TgLDH), T. annulata (TaLDH), Mus musculus (mouse LDh), Cricetulus griseus (hamster LDH), and human (H. sapiens LDH).
Characterization of expressed recombinant BmLDH protein
The recombinant GST-fussed BmLDH (GST-BmLDH) had a molecular weight of approximately 62 kDa (Fig. 2B, lane 4). Since the molecular weight of GST is 26 kDa, it is estimated that the molecular weight of BmLDH protein is 36 kDa. Over 70% of the recombinant protein was insoluble in water upon over expression in E. coli (Fig. 2A, lane 2). The average yield of the GST-BmLDH and GST-free BmLDH from the soluble fraction was approximately 200 μg/mL (Fig. 2B, lanes 4 and 5).
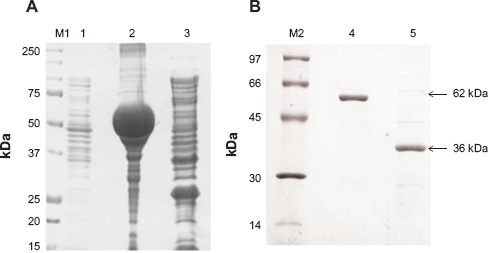
SDS-PAGE (
Characterization of native BmLDH protein
Analysis of parasite lysate by Western blot method revealed a specific 36 kDa native BmLDH (Fig. 3, lane 1). No band appeared when mice SPF sera was used as control as shown in Figure 3, lane 2.
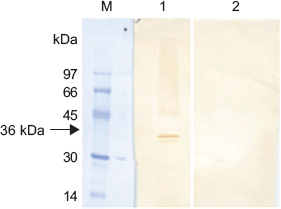
Western blot lane M: molecular marker stained with amide black; lane 1 is 36 kDa native BmLDH in parasite lysate after probe with polyclonal sera raised against BmLDH; lane 2 is parasite lysate probed with SPF sera (negative control).
The polyclonal antibody raised against recombinant BmLDH was able to detect the native BmLDH in infected RBCs by indirect IFAT. This was shown by specific green fluorescence observed in the cytoplasm and nucleus of B. microti merozoites (Fig. 4). The location of the green fluorescence in both the merozoite cytoplasm and nucleus is consistent with the finding from the cNLS mapper, which showed that BmLDH has both cytoplasmic and nucleus localization signal sequence from amino acid number 241–269 (EIIKLKGYTSWAIGLSVGDLSCSLIKNLR).
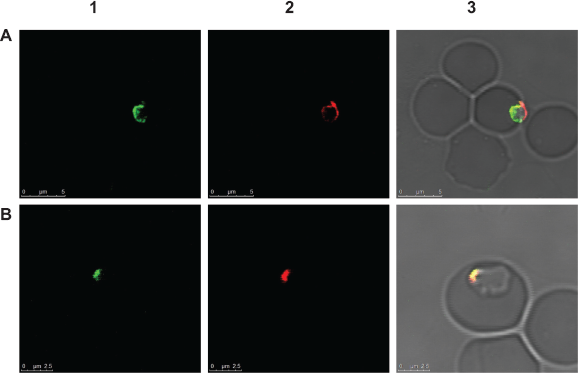
(
BmLDH enzyme activity
Optimal pH, Vmax, and Km
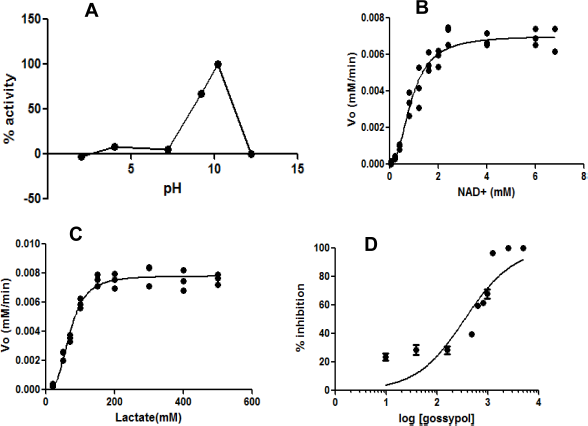
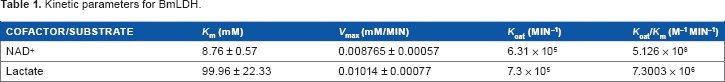
The recombinant BmLDH protein is an active enzyme with a high catalytic efficiency at optimal pH of 10.2 (Fig. 5A). The Km values for NAD+ and lactate were 8.7 ± 0.57 mM and 99.9 ± 22.33 mM, respectively (Fig. 5B and C). BmLDH had very high catalytic activity for both NAD+ (6.3 × 10 5 minute–1) and lactate (7.3 × 10 5 minute–1). Similarly, a catalytic efficiency of 5.126 × 10 8 M–1 minute–1 and 7.3003 × 10 6 M–1 minute–1 was recorded for NAD+ and lactate, respectively (Table 1).
(
Kinetic parameters for BmLDH.
Inhibition of BmLDH by gossypol
Gossypol (NAD+ analog) caused over 20% inhibition of BmLDH catalytic activity at very low concentration of 9.5 nM. Moreover, 100% inhibition of BmLDH enzyme activity was recorded when 2.5 μM of the inhibitor was added to the assay. The IC50 value for gossypol was 345.0 ± 1.11 nM (Fig. 5D).
Discussion
This study revealed that both B. microti Gray strain and R1 strain have identical LDH and share high frequency of identical amino acid with their mammalian counterparts. This suggests that B. microti LDH was probably acquired through lateral gene transfer. This is possible because the chromosome on which the LDH gene is located is known to be susceptible to double-stranded breaks, hence suitable for insertion of foreign DNA. 14 The acquired genetic sequence can often be modified by the parasite to suit its survival mechanism in the host. 25 However, it is worth noting that despite the above similarity in amino acid sequence, BmLDH has different amino acid residues in its antigenic domains (Supplementary Table 1). This suggests that the host immune system can detect the antigenic epitopes of BmLDH and produce antibodies against them. Thus, explaining why anti-rBmLDH polyclonal sera raised in mice was able to detect native BmLDH by Western blot and IFAT. Confocal laser microscopy revealed that native BmLDH is dominantly localized in the cytoplasma, although cases of nuclear localization were also observed. The above finding agrees with previous reports that while LDH is dominantly expressed in the cytoplasm, it can be found in the nucleus.26,27 The role of LDH in the nucleus remains unclear but it is understood that their association with DNA may play a role in transcription and replication of DNA through general stabilization of the nuclear matrix or chromatin structure. 27
The enzyme kinetic studies revealed that BmLDH has higher catalytic activity and efficiency. This finding is consistent with earlier report that B. microti depends extensively on glycolysis for its energy requirement, and BmLDH plays a key role. 14 The high catalytic activity (high Kcat value) of BmLDH for lactate compared to B. bovis LDH, P. falciparum LDH, T. gondii, and human LDH further suggests that it could be a novel candidate for drug target.19,28–30 Indeed, BmLDH inhibitory assay showed that the IC50 value for gossypol (a competitive antagonist for NAD+) was 0.345 μM. At 2.5 μM, catalytic activity of BmLDH was completely inhibited. This finding is consistent with previous reports that LDH is a novel drug target against some protozoan parasites.15,17–19 Since B. microti lacks well-developed tricarboxylic acid cycle, we hypothesize that inhibition of glycolysis via BmLDH would lead to energy starvation and death of the parasite. However, there is need for further research on in vivo efficacy of BmLDH inhibitors in B. microti-infected hamsters. Such a study would also generate important information on the safety of LDH inhibitor at therapeutic dose. While LDH plays a key role in glycolysis in mammalian red blood cells and muscles, suppression or deficiency of LDH causes benign conditions with no or minimum adverse effects.31,32 Merkle and others 33 have also reported that up to 50% reduction of LDH-A activity in mice produced no obvious harm. This probably explains why there is a growing momentum in the synthesis of new LDH inhibitors as potential drug against some protozoan organisms 20 and their potential use as anti-tumor drugs.34,35
Conclusion
This study revealed that BmLDH is a 36 kDa protein that has nucleoplasmin localization. The kinetic parameters of BmLD also suggest that the enzyme has a very high catalytic activity compared to other apicomplexan LDH; moreover, its activity can be inhibited at low concentration of NAD+ analog, gossypol. We therefore propose further studies to explore the in vivo efficacy of BmLH inhibitor in hamster model.
Author Contributions
PV, TM, SC, MAT, AAM, KK, YN, and XX conceived and designed the experiments. PV, SC, TM, PFAM, and MAT analyzed the data. PV, SC, and XX wrote the first draft of the manuscript. PV, TM, SC, AAM, PFAM, KK, and XX contributed to the writing of the manuscript. PV, TM, SC, MAT, AAM, KK, PFAM, YN, and XX agree with manuscript results and conclusions. PV, SC, TM, and XX jointly developed the structure and arguments for the paper. MAT and XX made critical revisions and approved final version. All authors reviewed and approved the final manuscript.
Footnotes
Supplementary Data
Unique amino acid residues at antigenic domain of BmLDH compared to other apicomplexan parasites and mammalian LDH.
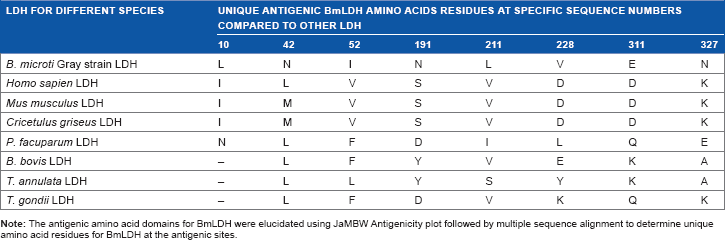
| LDH FOR DIFFERENT SPECIES | UNIQUE ANTIGENIC BmLDH AMINO ACIDS RESIDUES AT SPECIFIC SEQUENCE NUMBERS COMPARED TO OTHER LDH |
|||||||
|---|---|---|---|---|---|---|---|---|
| 10 | 42 | 52 | 191 | 211 | 228 | 311 | 327 | |
| B. microti Gray strain LDH | L | N | I | N | L | V | E | N |
| Homo sapien LDH | I | L | V | S | V | D | D | K |
| Mus musculus LDH | I | M | V | S | V | D | D | K |
| Cricetulus griseus LDH | I | M | V | S | V | D | D | K |
| P. facuparum LDH | N | L | F | D | I | L | Q | E |
| B. bovis LDH | – | L | F | Y | V | E | K | A |
| T. annulata LDH | – | L | L | Y | S | Y | K | A |
| T. gondii LDH | – | L | F | D | V | K | Q | K |
Acknowledgment
We acknowledge the technical contributions made by members of the Host defense, Babesia and Vaccine laboratories, National Research Center for Protozoan diseases (NRCPD), Obihiro University of Agriculture and Veterinary Medicine. The contributions offered by Dennis Muhanguzi, Maki Ishiwata, Miki-Araki, and Ken Saito are acknowledged.