
Abstract
Tissue factor (TF) is an integral membrane protein widely expressed in normal human cells. Blood coagulation factor VII (fVII) is a key enzyme in the extrinsic coagulation cascade that is predominantly secreted by hepatocytes and released into the bloodstream. The TF–fVII complex is aberrantly expressed on the surface of cancer cells, including ovarian cancer cells. This procoagulant complex can initiate intracellular signaling mechanisms, resulting in malignant phenotypes. Cancer tissues are chronically exposed to hypoxia. TF and fVII can be induced in response to hypoxia in ovarian cancer cells at the gene expression level, leading to the autonomous production of the TF–fVII complex. Here, we discuss the roles of the TF–fVII complex in the induction of malignant phenotypes in ovarian cancer cells. The hypoxic nature of ovarian cancer tissues and the roles of TF expression in endometriosis are discussed. Arguments will be extended to potential strategies to treat ovarian cancers based on our current knowledge of TF–fVII function.
Introduction
Epithelial ovarian cancer (EOC) is the most fatal gynecologic malignancy worldwide. Most cases are diagnosed at advanced stages, and the overall five-year survival rate of patients with EOC is 45%. 1 EOC can be classified based on multiple histological subtypes. Generally, five histological subtypes (low-grade serous carcinoma, high-grade serous (HGS) carcinoma, clear cell carcinoma (CCC), mucinous carcinoma, and endometrioid (EnM) carcinoma with different characteristics of EOC are known to exist. 2 HGS ovarian cancer is the most prevalent type, accounting for approximately 70% of ovarian cancers. 3 HGS tumors are relatively sensitive to chemotherapy, including those using platinum- and taxane-based agents. Clinical trials have revealed that drugs targeting signaling cascades activated in ovarian cancer cells, such as the PI3K- and RAS-signaling pathways, are promising candidates for the treatment of EOC in the future. 4 However, disease diagnosed at late stages or relapsed disease is difficult to treat. Moreover, certain histological subtypes of EOC, such as low-grade serous carcinoma and CCC, are resistant to chemotherapy. 2 Improving our understanding of the biology of ovarian cancer cells may lead to the development of new therapeutic strategies for the treatment of aggressive EOCs associated with poor prognosis.
Blood coagulation factor VII (fVII) is an approximately 50 kDa precursor of a serine protease involved in the extrinsic coagulation cascade. 5 This unprocessed protease precursor is produced predominantly by hepatocytes and released into the bloodstream. Tissue factor (TF) is an ~50-kDa transmembrane lipoprotein that functions as the cellular receptor for fVII.5,6 The concentration of fVII in the human blood is the second lowest among plasma coagulation factors; 5 however, upon injury, fVII interacts with TF and accumulates on the exposed cell surface, leading to its conversion to the active form factor VIIa (fVIIa).5,6 The serine protease activity of the TF–fVIIa complex initiates the blood coagulation cascade, thereby leading to fibrin deposition5,6 (Fig. 1). In addition to its procoagulant activity, the cytoplasmic domain of TF plays an important role in the biology of cells by transmitting signals. This binary complex also contributes to cellular angiogenic and inflammatory responses via phosphorylation of the cytoplasmic domain of TF, followed by activation of cellular signaling cascades.6,7
A number of studies have revealed that TF is overexpressed in various normal
8
and cancer tissues, including those of pancreatic, ovarian, and breast cancers.
9
In addition to the physiological functions described above, the protease activity of the TF–fVIIa complex, which occasionally forms a ternary complex by association with activated coagulation factor X,
6
enhances key pathogenic events in cancer progression, such as cell motility, invasion, angiogenesis, and cell survival. This function occurs via activation of protease-activated receptors (PARs)
6
as exemplified for protease-activated receptor-2 (PAR2) activation (Fig. 1). Phosphorylation of the cytoplasmic domain of TF leads to its association with PAR2 to activate cellular signaling pathways that promote breast cancer development
10
(Fig. 1). Moreover, the cytoplasmic domain of TF is responsible for the experimental metastasis of melanoma cells in vivo,
11
suggesting that a coagulation-independent pathway of TF–fVIIa-driven phenotypes is possible. Furthermore, TF can be secreted from cancer cells,12–14 and this aberrant TF expression is associated with venous thromboembolism (VTE) and pulmonary embolism, which are major complications that occur in cancer patients and are known as Trousseau's syndrome. TF levels are high in preoperative ovarian cancer patients and can be an independent prognostic factor.
15
TF–fVlla complex formation on the surface of EOC cells results in the activation of the extrinsic coagulation cascade and/or PARs. The serine protease activity of the TF–fVlla binary complex associated with the plasma membrane initiates activation of downstream coagulation cascades associated with coagulation factors (IX, X, prothrombin, and fibrinogen). Otherwise, this protein complex cleaves the N-terminal end of PARs. PARs are then activated via intramolecular binding between the newly created N-terminus and an extracellular loop region of the receptors. Activation of G-protein-coupled receptors subsequently activates downstream signaling cascades. Phosphorylation of the C-terminal end of TF could also lead to association with PAR2 in a coagulation-independent manner to augment the signaling cascade.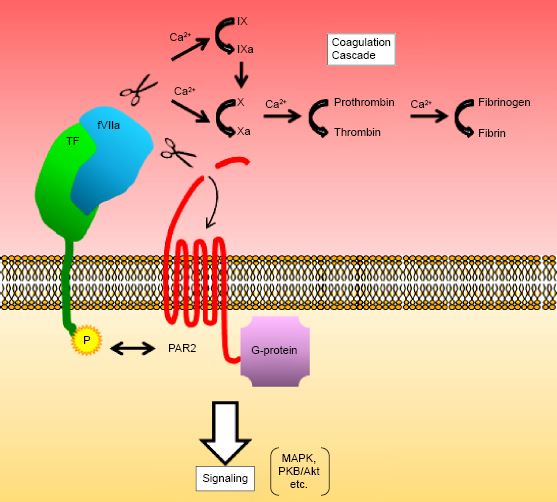
Within tumor tissues, the overexpression of angiogenic factors, such as vascular endothelial growth factor (VEGF), can lead to the hyperpermeabilization of blood vessels. 16 Therefore, the function of TF on the surface of cancer cells within tumor tissues can be enhanced via fVII derived from blood plasma in the absence of vascular injury. Accordingly, fVII may be more abundant within tumor tissues than in non-cancerous tissues.
In addition to aberrant TF expression in EOC cells and tissues, fVII is synthesized in various EOC cells and tissues.17,18 Ectopic fVII expression is relatively rare, but it is frequent in CCC tissues. 18 Ectopic fVII expression followed by TF–fVIIa complex formation on the surface of CCC cells activates cell motility and invasiveness. 17 Notably, fVII expression is induced at the transcriptional level in CCC cells in response to hypoxia,17–20 leading to shedding of extracellular vesicles 21 enriched in the TF–fVIIa complex. 18 This experimental evidence suggests that TF–fVIIa is a key regulator of ovarian cancer phenotypes. Here, we summarize the potential roles of the TF–fVIIa complex in EOC phenotypes and analyze the mechanisms regulating TF–fVIIa activity associated with EOC cells. Current knowledge of the transcriptional regulation of TF and fVII in CCC cells in response to hypoxia, a general characteristic of the tumor microenvironment, is discussed to help in designing the strategies for the inhibition of their aberrant gene expression. Arguments will be extended to the potential roles of this procoagulant protein complex in the development of endometriosis, a gynecological disorder considered as a precursor of some histological subtypes of EOC. We finally discuss how current knowledge of TF–fVIIa signaling mechanisms can be translated into the development of promising therapeutic strategies for the treatment of aggressive gynecological disorders.
TF and fVII Expressions in Ovarian Cancer Tissues: Relationship with Histological Subtypes
Ovarian cancer is a heterogeneous disease that can affect tumor tissues not derived from the ovary, such as uterine and fallopian tube tissues.2,3,22 Thus, ovarian cancer should be regarded as a general term to define cancers in the pelvic and peritoneal cavities. The serous types are the most common worldwide,3,23 and HGS carcinoma is relatively chemosensitive. CCC is relatively frequent in some Asian and European countries, 23 and its known resistance to chemotherapy 24 makes it difficult to treat and is associated with its poor prognosis.
Since the definition of Trousseau's syndrome, TF has been suspected as a major factor linking various disorders, including cancer, and accompanying thromboembolic diseases, such as VTE and pulmonary embolism. The expression of TF in normal and cancer tissues was investigated at the protein level by immunohistochemistry (IHC).8,18,25 TF is a vital component of the extrinsic coagulation cascade, and it is therefore expressed at different levels in various normal tissues, including normal human ovarian surface epithelium (OSE).9,18 TF is overexpressed in many cancer tissues; however, some cancers such as renal cancer and lymphoma lack TF expression. 9 TF overexpression may contribute to hypercoagulation observed in cancer patients. The incidence of VTE is highest among pancreatic, liver, and ovarian cancer patients,12,13 and TF expression has therefore been analyzed in clinical ovarian cancer tissues in relation to the frequency of VTE. Earlier studies showed that TF is expressed in the human OSE in a manner similar to other normal tissues. TF expression in EOC tissues varies according to the individual patient and tends to be high in certain histological subtypes. 25 However, its expression level is not necessarily higher in cancer tissues than in normal tissues. 9 IHC experiments showed that the expression level of TF is significantly higher in CCC tissues than in other histological types of EOC.18,25 A recent study using IHC and enzyme-linked immunosorbent assay (ELISA) showed that TF is abundant in CCC and EnM tissues. 26
The expression of fVII was also examined by IHC using clinical ovarian cancer specimens, which showed that fVII is expressed at higher levels in CCC tissues than in EOC tissues with other histological origins and normal OSE. 18 These results are consistent with the fact that hypercoagulation is more frequent in CCC patients than in cancer patients with other histological subtypes.25,27 Recently, genome-wide mutation profiling was performed in multiple ovarian cancer cell lines for the genetic characterization of CCC cells. 2 The general features of CCC cells were no mutations in the TP53 gene, overexpression of HNF1B, and mutations in the ARID1A gene. 2 These data question the reliability of the histological identification of CCC.
Ovarian Cancer Cell Lines Used for TF and fVII Expression Analysis
Cancer phenotypes regulated by TF–fVIIa signaling have been widely studied using different cell lines, including ovarian cancer cell lines. Particularly, TF–fVIIa signaling mechanisms associated with breast cancer biology have been widely investigated and reviewed extensively.6,28,29 Similar mechanisms may be applicable to TF–driven phenotypes of ovarian cancer cells. Ovarian cancer is a heterogeneous disease, and the expression of TF and fVII is frequent in CCC. Given that CCC is associated with a relatively poor prognosis among EOCs with other histological subtypes; CCC cells with high TF levels are ideal model cell lines to explore the TF–fVIIa-driven functions of EOC.
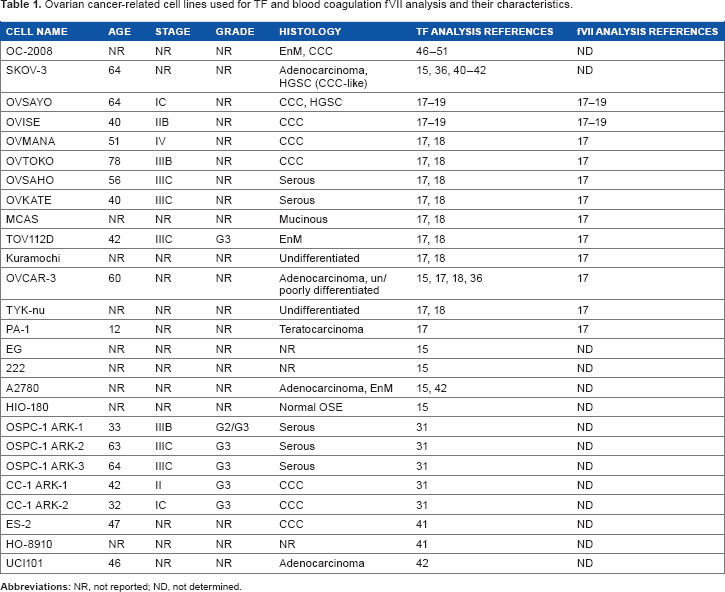
To date, multiple ovarian cancer cell lines have been used for TF analysis (Table 1). Initial analysis of histological subtypes identified seven cell lines30,31 as CCC-type ovarian cancer cell lines. Five cell lines30,31 are serous type, one is mucinous type, 2 and two are EnM cell lines. 2 Four adenocarcinoma cell lines without reported histotypes have been described.2,32–34 Genome-wide mutation analysis revealed that A2780 can be potentially classified as EnM-type cancer cells. 2 Two lines are undifferentiated or poorly differentiated types of cell lines. 2 OVCAR-3 cells can be categorized in this histotype because this cell line was established from papillary adenocarcinoma with a poorly differentiated histological subtype. 33 HIO-180 has been used as a normal OSE cell line.15,35 Recent genome-wide sequencing and IHC analyses have raised concerns regarding previously reported histological subtypes. For example, it was suggested that the OVSAYO cell line should be regarded as HGS carcinoma rather than CCC 2 and that the SKOV-3 cell line, while histologically similar to HGS carcinoma, genetically resembles CCC 2 . OC-2008 cells, which were initially identified as EnM-type cancer, are potentially CCC cells. 2
Ovarian cancer-related cell lines used for TF and blood coagulation FVII analysis and their characteristics.
The expression of the TF (F3) gene has been examined at the mRNA and protein levels in multiple cell lines (Table 1) by reverse-transcription polymerase chain reaction15,17,18,31 and western blotting. 18 Four CCC lines (OVSAYO, OVISE, OVTOKO, and OVMANA), MCAS, and Kuramochi cells express TF mRNA at considerable levels when cells are exposed to hypoxia or hypoxia-mimetic stimuli.17,18 A more recent study showed that CCC cell lines (CC-1 ARK-1 and CC-2 ARK-2) have high levels of TF transcripts compared with those in serous-type cell lines. 31 Studies using SKOV-3 and OVCAR-3 cells showed that the expression level of the F3 gene is regulated via multiple signaling cascades. 36
Subsequent to TF analysis, fVII expression has been examined because this procoagulant is ectopically synthesized in various cancer cells. 17 Constitutive expression of fVII is generally low in ovarian cancer cells. 17 However, constitutive fVII mRNA levels are relatively high in the Kuramochi cell line. 17 Moreover, as discussed later, this transcript is increased in many EOC cell lines when cells are cultured under conditions of hypoxia or hypoxia-mimetic CoCl2 stimulation.17–19
General Biology of TF–fVII Signaling Associated with Ovarian Cancer Cells
Ovarian cancer phenotypes regulated by the TF–fVII complex
The amino acid sequence of TF is similar to that of the cytokine/growth factor receptor family, suggesting that the TF–fVIIa complex can transmit signals independently of the coagulation cascade.37,38 TF–fVIIa triggers intracellular signaling cascades through multiple molecular pathways, predominantly those associated with PAR2 (Fig. 1). TF–fVIIa signaling associated with cancer progression, such as that involved in increased cell motility, invasiveness, angiogenesis, and cell survival, has been widely studied using breast cancer cell lines. 6 Few studies have analyzed TF–fVIIa signaling mechanisms directly associated with ovarian cancer cell phenotypes, and such mechanisms associated with breast cancer biology may also be applied to ovarian cancer cells with high TF expression. Briefly, the binary complex of TF–fVIIa (occasionally a ternary complex with factor X to produce a TF–fVIIa-factor Xa (fXa) complex), could activate downstream cellular signaling cascades involving MAPK, protein kinase B (PKB/Akt), and mTOR via PARs, leading to various malignant phenotypes of breast cancer cells. 6
Currently, there are multiple reports concerning TF–fVIIa complex-driven phenotypes of EOC cells. The motility and invasiveness of the ovarian cancer cell line OVSAYO can be increased by forced expression of fVII. 17 These phenotypes were found to be mediated via formation of the TF–FVIIA complex and PAR1 on the cell surface. PAR1 is cleaved and activated by thrombin. However, hirudin, a potent thrombin inhibitor, failed to inhibit these phenotypes, whereas NAP-5, a specific fXa inhibitor, suppressed them. 17 This suggests that the TF–fVIIa-fXa complex, which is presumably formed on the surface of OVSAYO cells, can directly activate PAR1, as evidenced by experiments using HeLa cells, 39 thereby facilitating cell motility and invasion.
Relatively high concentrations of low-molecular-weight heparin block TF–dependent motility and invasiveness in various cancer cells, including SKOV-3 cells in collagen-coated membranes, 40 suggesting that procoagulant activity contributes to the expression of these phenotypes. TF–fVIIa complex activity on the surface of immune cells is important to activate the invasiveness of EOC cells within tumor tissues. TF–fVIIa complex formation stimulates monocytes to highly produce IL-8 and transcription factors, such as hypoxia-inducible factors (HIFs) and Egr-1. 41 IL-8 released from monocytes attracts cancer cells and mediates EOC cell invasion. 41 A recent study showed that platelets isolated from EOC patients act as chemoattractants, thereby increasing motility. 42 Further, platelets enhance sphere formation activity and TF expression in several EOC cell lines, including SKOV-3, A2780, and UCI101. 42 These data are true for primary cultured ovarian cancer cells prepared from ascites of patients. 42 These results suggest that EOC cells in the peritoneal environment can be stimulated by platelets to increase TF expression levels, although the detailed molecular mechanism remains unclear.
In these studies, whether autonomously produced fVII contributes to the TF-dependent phenotypes of EOC cells remains unclear, as ectopic fVII expression was not examined.
Regulation of TF–fVIIa complex activity on the surface of EOC cells
One of the cell lines initially used to characterize the functions of the TF–fVIIa complex is the OC-2008 cell line (Table 1), which constitutively expresses TF. As described above, the derived histological subtype of this cell line was initially reported as EnM. 2 Recent genome-wide mutation and IHC analyses have suggested that this cell line is derived from CCC. 2
Treatment with heparin is a common method to protect cancer patients from thrombotic events, such as VTE. 43 This anticoagulant activates antithrombin III (ATIII) and inhibits procoagulants such as thrombin and fXa. 44 Tissue factor pathway inhibitor-1 (TFPI-1) is a Kunitz-type serine protease inhibitor predominantly synthesized in endothelial cells. This anticoagulant protein can also be synthesized in immune and fibroblast cells. 45 TFPI-1 directly inhibits the enzymatic activity of the TF–fVIIa complex, 45 thereby regulating normal hemostatic processes. The kinetics of the protease activity of the TF–fVIIa complex and its negative regulation by TFPI-1, ATIII, and heparin have been analyzed in detail using the OC-2008 cell line.46–49 These studies revealed that two distinct TF–fVIIa complexes can be formed on the monolayer of this ovarian cancer cell line. One is the active complex formed at the initial stage of the TF–fVII interaction. The other is an inactive complex that can be formed at relatively late stages of the TF–fVII interaction. This complex can be activated via exposure to anionic phospholipids in response to various stimuli, such as cell damage. Studies further demonstrated that the TF–fVIIa complex on the surface of OC-2008 cells can be inhibited by both TFPI-1 and ATIII/heparin treatment, albeit via different molecular mechanisms. These results have provided molecular insights into how extrinsic blood coagulation can be tuned under physiological conditions and why heparin, unlike warfarin, can block long-term intravascular hypercoagulation states, such as those associated with VTE and Trousseau's syndrome.
TF–fVIIa activity on the OC-2008 cell surface can also be influenced by cellular cofactors, as the enzymatic activity of TF varies depending on its source, namely, the cell membrane, cell lysate, or reconstituted purified protein.50,51 These studies raised the possibility that cofactors associated with the cell membrane, such as the anionic phospholipids described above, and Annexin V could affect TF–fVIIa activity. This regulatory mechanism by anionic phospholipids is currently known as encryption-decryption, 6 which describes a manner of regulation of TF activity mediated by its conformational change.
Relationship between VTE and TF–fVIIa Complex Activity in Ovarian Cancer Cells
Increased coagulation activity has been observed in cancer patients for many years. A number of risk factors are associated with VTE in gynecologic oncology patients. 52 An earlier IHC study with specimens of ovarian cancer patients demonstrated that coagulation mediators, including TF, are expressed within tumors, thereby contributing to fibrin generation. 53 Indeed, later studies showed that VTE is frequent in ovarian cancer patients12,13,25 and correlates with reduced survival rates.54,55 Accumulating experimental evidence suggests that the histological type of EOC with the highest incidence of VTE is CCC.24–27,56–58 It is conceivable that TF plays a major role in VTE in ovarian cancer patients, given that TF can be highly expressed in EOC tissues. Indeed, IHC analysis revealed that TF tends to be highly expressed in CCC tissues.25,26 This expression pattern also correlates with the concentration of a fibrin degradation product, D-dimer, and with the incidence of VTE, rather than disease stage and patient characteristics. 25 In this study, ectopically synthesized fVII was not examined, and thus, its contribution to VTE development is unclear.
fVII can be induced in ovarian cancer cells in response to hypoxia; therefore, the expression of TF and fVII in ovarian cancer tissues was estimated by IHC, which showed that in addition to TF, the level of fVII is higher in CCC tissues compared with cancer tissues with different histological types and an OSE tissue. 18 Additionally, CCC and EnM-type tissues from patients with VTE express high levels of TF and TFPI-1 transcripts. 26 ELISA assays confirmed that TF is increased in CCC and EnM tissues at the protein level. 26 TFPI-1 protein expression tends to be high in EOC tissues, although its upregulation is not associated with the incidence of VTE. 26 Given that TFPI-1 is a negative regulator of TF–fVIIa activity, the significance of high TFPI-1 expression in VTE-prone CCC patients is unclear. TFPI-1 might affect the mechanism of VTE via molecular mechanisms not correlated with its anti-coagulation function. Indeed, TFPI-1 plays a negative role in the adhesion of MDA-MB-231 cells to collagen I. 59 Further, the interaction of exogenous TFPI-1 with the TF–fVII complex on the surface of bladder cancer cells enhances cell adhesion to fibronectin, 60 suggesting that ectopic TFPI-1 plays multiple roles in the expression of cancer cell phenotypes.
Extracellular vesicles, such as exosomes and microvesicles (MVs), are secreted from cancer cells into the bloodstream.
21
MVs are derived from the plasma membrane and are associated with lipid rafts.
61
This relatively larger vesicle can be a vehicle for TF;
61
therefore, shedding of MVs into the bloodstream could be a cause of VTE in cancer patients (Fig. 2). Indeed, CCC cells can shed TF–fVIIa complex-positive MVs via autonomous production of fVII in vitro.
18
The released MVs have high procoagulant activity, and this activity can be blocked by treatment with anti-TF antibodies,
17
suggesting that most of the TF integrated into MVs is in decrypted (coagulation active) form.
6
These facts raise the possibility that ectopically synthesized fVII also contributes to the high VTE rate in ovarian cancer patients (Fig. 2), although thrombus formation by EOC-derived MVs has not been directly demonstrated in vivo.
TF-fVIIa-positive MVs released from CCC cells may cause VTE. CCC cells secrete plasma membrane-derived MVs associated with the TF–fVIIa complex. The expression of both TF and fVII is upregulated in CCC cells in response to hypoxia. Production of TF-fVIIa-rich MVs is expected under conditions of hypoxia. These procoagulant MVs released into the bloodstream may cause VTE.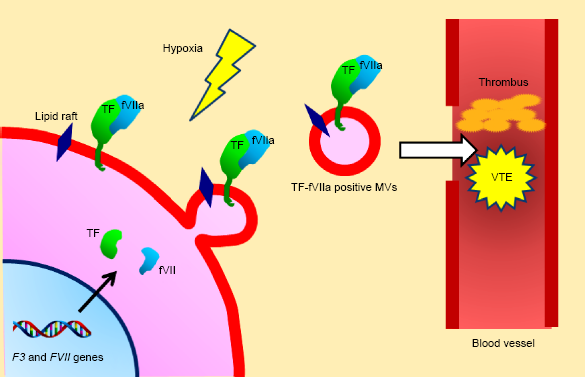
Additionally, the secretion of MV-free TF in its full-length or truncated form has been reported for some cancer cells.62,63 Thus, the release of these soluble TF forms may contribute to the incidence of VTE in CCC patients, although evidence supporting this hypothesis has not been provided to date.
Ectopic Expression of Tissue Factor Pathway Inhibitor-2 (TFPI-2) in Ovarian Cancer Cells
TFPI-2 is an inhibitor of various serine proteases with a similar amino acid sequence to that of TFPI-1.64,65 TFPI-2 is synthesized in fibroblasts and endothelial cells. 66 TFPI-2, which is particularly abundant in the placenta, is also referred to as placental protein 5. 65 As in the case of TFPI-1, TFPI-2 has three Kunitz domains,64,65 and inhibition of TF-fVIIa complex activity by TFPI-2 is weaker than that mediated by TFPI-1. 67 TFPI-1 is synthesized in ovarian cancer cells, including CCC. However, a recent proteome analysis showed that TFPI-2, but not TFPI-1, is highly expressed in CCC cell lines and clinical tissues compared with its expression in ovarian cancers with different histological types. 68 In addition to the expression of TFPI-2 in the cell body, this protease inhibitor can be highly secreted from CCC cells into the culture medium. 68 TFPI-2 levels are high in serum samples of CCC patients compared with those in other histotype ovarian cancers. 68 These results suggest that TFPI-2 is a new biomarker for CCC patients. According to this principle, OVSAYO is still categorized as a CCC cell line because it synthesizes high levels of TFPI-2. 68 The biological roles of TFPI-2 in CCC cells are currently unclear, although it has been suggested to play roles in vasculogenic mimicry 67 and suppression of invasiveness 69 of melanoma and choriocarcinoma cells, respectively.
Additionally, these results are inconsistent with cDNA microarray experiments showing that the mRNA level of TFPI-2 in clinical CCC samples is low compared with that in OSE tissues. 70 Although CCC is derived from the endometrium, 24 the differences in the relative levels of TFPI-2 mRNA between CCC tissues and endometrial tissues remain unclear. Elevated levels of TFPI-1 transcripts were not detected in this microarray analysis.
Transcriptional Regulation of TF and fVII in Ovarian Cancer Cells
Transcriptional regulation of genes is a vital mechanism that determines the expression levels of the corresponding proteins in cells. Recent studies suggested that microRNAs are involved in the regulation of TF levels at the translational level in breast cancer cells. 6 The function of TF can be modulated by its irreversible conformational change, which is referred to as encryption-decryption. 6 At present, the roles of the microRNA-driven regulation of TF function associated with ovarian cancer cell biology have not been reported. Here, we describe potential transcriptional regulation mechanisms of TF and fVII expression in ovarian cancer cells. A common feature of transcriptional regulation mechanisms of TF and fVII in ovarian cancer cells is that both transcriptions can be enhanced in response to hypoxia, 20 a general characteristic of tumor tissues.
Hypoxia and ovarian cancer tissues
Hypoxia, a stress condition, is a common feature of tumor tissues associated with their aberrant and sparse vasculature compared with that of normal tissues. 71 Cancer cells can adapt to such severe environments within tumor tissues by using multiple stress response mechanisms, such as transcriptional regulation by HIFs. 72 To date, the hypoxia status of ovarian cancer tissues has been tested by various experimental means. The oxygenation state of xenograft ovarian tumors was directly measured by electron paramagnetic resonance oximetry. 73 An adeno- or endometrioid-carcinoma cell line, A2780, 2 was used for this study, which showed that the tumor tissue is severely hypoxic.
Pimonidazole is a chemical agent that reacts with amino acid residues of proteins in cells exposed to severe reducing (hypoxic) conditions. IHC analysis of A2780 tumors orthotopically (intraperitoneally) implanted into nude mice 74 and OVISE tumors subcutaneously implanted into non-obese diabetic/severe combined immunodeficiency mice 75 using antibodies raised against pimonidazole amino acid adducts demonstrated that ovarian tumor tissues are severely hypoxic. Furthermore, a SKOV-3 tumor model revealed that ovarian cancer cells form spheroids and grow in the peritoneal cavity, and their core was severely hypoxic. 76 These results are consistent with the general concept that solid tumors contain areas of severe hypoxia. 75
Ovarian cancer patient samples were examined by IHC using antibodies against hypoxia-inducible factor-1 (HIF1). These studies showed that HIF1 is expressed in serous and CCC-type ovarian cancer tissues and its expression levels correlate with disease prognosis.77,78 HIF1 staining of OVISE xenograft tumor tissues showed that HIF1 exists in the nuclei of cancer cells within hypoxic tissue areas close to pimonidazole-adduct positive tumor regions. 75 These data also demonstrate the hypoxic nature of ovarian cancer tissues and their clinical importance. However, they are rather indirect evidences of tissue hypoxia, as HIF1 can also be upregulated in ovarian cancer cells via hypoxia-independent signal trans-duction mechanisms, such as activation of the IL6-STAT3-HIF signaling pathway. 78
Given that tumor tissues are supplied with variable levels of blood components in addition to O2, hypoxic tumor tissues that are distant from blood vessels should also be poorly supplied with blood components. Intriguingly, it was recently suggested that severe hypoxic regions within OVISE tumors contain few neutral lipids probably because of an insufficient supply of long chain fatty acid (LCFA)-albumin complex from the bloodstream. 75
Ovarian cancer cells disseminate in the peritoneal cavity by direct migration and invasion from the original tumor. 79 The majority of these migrated cancer cells float and survive in the peritoneal fluid, resulting in further progression of the disease. Although ovarian tumors are hypoxic, the oxygenation status of the peritoneal environment remains unknown; therefore, the hypoxia status of the ascites fluid of ovarian cancer patients was measured by gas analysis. 80 The results showed that ascites is hypoxic (~46 mmHg); however, the level of hypoxia is lower than that of tumor tissues (~2–8 mmHg). This study also performed IHC using a xenograft tumor model of the SKOV-3 cell line and showed that tumors were HIF1 positive. 80
Adipose tissues within the omentum are another candidate hypoxic environment within the peritoneal cavity that depends on physical conditions such as obesity. 81 In the peritoneal environment, ovarian cancer cells may predominantly use LCFA as an energy source rather than glucose. 82 In vivo experiments showed that orthotopically implanted ovarian cancer cells can be led to the omentum to receive LCFA from the adipose tissues. 82 Indeed, the growth of SKOV-3 xenograft tumors is dependent on LCFA supply, followed by the formation of cellular neutral lipids. 82 Overall, various peritoneal environments surrounding ovarian cancer tissues are hypoxic, and adaptive responses to this stress condition may play an important role in the progression of ovarian cancer.
Expression mechanism of TF in ovarian cancer cells
TF is encoded by the F3 gene, whose expression is regulated by the transcription factor Sp1, as previously reviewed in detail.6,20 This transcription is inducible in normal and cancer cells in response to various proinflammatory stimuli. F3 gene expression is induced by proinflammatory stimuli via binding of the transcription factors NF-kB, AP-1, Egr-1, and NFAT, which are activated by various stimuli within the tumor microenvironment. 20 This general mechanism of TF overexpression may be applied to ovarian cancer cells given that activation of NF-kB,75,83 AP-1,75,84 NFAT, 85 and Egr-118,86 play important roles in the biology of ovarian cancer cells. Additionally, the expression of the F3 gene in ovarian cancer cells may be regulated by sex hormones, such as progesterone, 87 according to their hormone receptor positivity. 88 Whole genome expression analysis showed that the transcript level of F3 may be increased in clinical specimens of CCC. 70 However, these data are not conclusive because the microarray data were based on a comparison with OSE, which is not necessarily considered as an appropriate normal counterpart to CCC tissues.
Under conditions of hypoxia, the ubiquitination and pro-teasomal degradation of HIFs are inhibited, resulting in their accumulation in cancer cells. 72 HIFs bind to aryl hydrocarbon receptor nuclear translocator (ARNT) to occupy regulatory regions of target genes referred to as hypoxia response elements (HREs), which are required for adaptive responses to hypoxic stress (Fig. 3). The F3 gene is also inducible in response to hypoxia. This mechanism differs from the above conventional HRE-dependent transcriptional induction mechanism. Hypoxic F3 induction is mediated by the transcription factor Egr-1, and there have been no reports on the involvement of HIFs in F3 transcriptional regulation. Indeed, the TF protein is upregulated in ovarian cancer cells (OVSAYO and OVISE) in response to hypoxia. 18 Furthermore, oncogenic events associated with the EGFR/RAS pathway upregulate F3 expression in glioma cells. 89 The involvement of the EGFR signaling pathway in F3 expression in OVCAR-3 and SKOV-3 cells was reported previously. 36
Cellular signaling mechanisms regulating F3 expression were analyzed in the ovarian cancer cell lines OVCAR-3 and SKOV-3. 36 The PI3K-Akt signaling pathway is aberrantly activated in ovarian cancer cells. This study reported that PI3K-Akt and MAPK-ERK signaling cascades oppositely regulate F3 expression levels. 36 These results raise the possibility that a steady-state transcript level of TF may be determined by the relative significance of these signaling pathways, depending on the environmental conditions of ovarian cancer tissues.
Expression mechanisms of the FVII gene in ovarian cancer cells
The biology of ovarian cancer is largely influenced by estrogen exposure.
90
FVII gene expression may be affected by this sex hormone in ovarian cancer cells, as this gene contains an estrogen receptor binding site within the promoter region.
91
FVII was shown to be negatively regulated by estrogen exposure in a hepatocytic cancer cell line, HepG2.
91
Conversely, it is possible that the supply of fVII from the bloodstream to cancer tissues is suppressed in hypoxic tissues because of poor vascularization. Therefore, cancer cells can autonomously produce fVII in response to hypoxia by activating FVII transcription. The basal transcript level of the FVII gene in ovarian cancer cells is generally low,
17
in contrast to the high constitutive FVII expression in breast cancer cells.
17
However, FVII is upregulated in different ovarian cancer cell lines (OVSAYO, OVISE, and OVSAHO) in response to hypoxia.17–20 The mechanisms of inducible FVII gene expression have been dissected using the ovarian cancer cell lines OVSAYO and OVISE, and most of them have already been discussed.
20
Briefly, hypoxia-driven FVII activation is mediated by the transcription factor Sp1.
19
Unlike ARNT-dependent transcriptional activation, such as that mediating the expression of VEGF (Fig. 3), this mechanism is dependent on HIFs but not on ARNT. The interaction between hypoxia-inducible factor-2 (HIF2) and Sp1 plays a role in the hypoxic activation of FVII gene expression (Fig. 3). HIF1 can also be involved in this process, although in a manner independent of ARNT.
19
It is likely that the Per-Arnt-Sim domains of HIF2 mediate the interaction between Sp1 and HIF2.
19
Differential activation mechanisms of the FVII and VEGF genes. Left: activation of conventional hypoxia response genes, such as VEGF. Sp1 regulates the basal promoter activity of the VFGF gene. Right: hypoxic activation of the FVII gene is mediated by the interaction of Sp1 with HiF2a. HDAC4 could function as a coactivator of this transcription. Under conditions of hypoxia, the ARNT-HiF1α complex becomes bound to the HRE region and enhances the basal activity of the distal gene promoter region.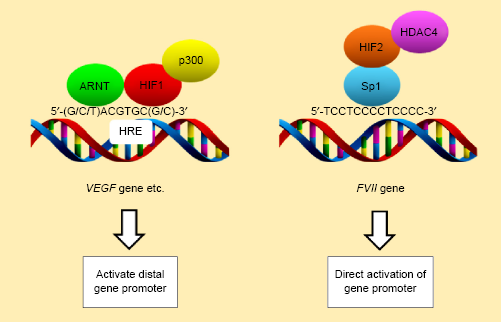
Sp1 is a ubiquitous transcription factor involved in the regulation of the majority of housekeeping genes. Although Sp1 preferentially binds GC-rich consensus gene regulatory sequences termed the GC-box, it binds a 5′-TCCTCC CCTCCCC-3′ sequence within the FVII promoter region in hepatocytes92–94 (Fig. 3). This is distinct from HRE sequences described as 5′-(G/C/T)ACGTGC(G/C)-3′ or 5′-RCGTGC-3′ 95 (Fig. 3). This CT-rich sequence is also vital for the ectopic expression of FVII. Both constitutive and inducible expressions of the FVII gene in cancer cells are dependent on Sp1 and associated with this sequence, 96 suggesting that this sequence acts as an alternative HRE with regard to the FVII gene (Fig. 3). Intriguingly, hypoxic FVII activation in ovarian cancer cells can involve deacetylation of histones within the FVII promoter region associated with the recruitment of the class II histone deacetylase HDAC419,20 (Fig. 3). This phenomenon is counterintuitive given that transcriptional activation normally involves hyperacetylation of histones within the chromatin of gene promoters.
Serum starvation synergistically enhances hypoxia-driven expression of the FVII gene in ovarian cancer cells
The levels of molecular oxygen and other serum components vary within tumor tissues depending on their tissue diffusibility. To adapt to this differential tissue hypoxia conditions associated with variable nutrient, growth factor, and hormone compositions, cancer cells utilize multiple molecular mechanisms in addition to HIF-mediated responses. The endoplasmic reticulum (ER) is an important cellular organ-elle responsible for the synthesis, subsequent modification, and folding of proteins. 97 Disruption of any of these events under stress conditions, such as hypoxia, causes ER stress. ER stress followed by the unfolded protein response (UPR) is an additional cellular response for adaptation to severe hypoxia. 98 The UPR can enhance the transcription of genes required for tumor survival and growth through the activation of ER-resident proteins, such as Perk, ATF6, and Ire-1, followed by upregulation of transcription factors. 98 Examination of the effect of serum starvation in conjunction with hypoxia on the ER stress condition in ovarian cancer cells showed that the UPR marker CHOP, a transcription factor, is expressed in OVSAYO cells cultured under hypoxia and serum starvation conditions.19,75 This suggested that this culture condition reflects the severe hypoxic conditions leading to ER stress. The transcript level of FVII is synergistically increased in response to serum starvation and hypoxia (SSH) in OVSAYO and OVISE cells. 19 However, FVII expression in ovarian cancer cells cultured under hypoxic conditions with serum stimulation was not enhanced by tunic-amycin (an ER stress-inducing agent) treatment. 19 Therefore, the synergistic expression of FVII under SSH conditions in ovarian cancer cells is not attributed to the UPR, although the detailed mechanisms of this synergism remain unclear. Recent studies showed that the ICAM1 gene is synergistically activated in ovarian cancer cells exposed to SSH; removal of an LCFA-albumin complex from the culture medium upon serum deprivation was shown to be responsible for the unusually robust upregulation of this gene.75,99
Relationship between TF–fVIIa Signaling and Endometriosis
Endometriosis is defined as a disorder associated with endometrial tissues existing outside of the uterine endometrium. This disease is the most common cause of morbidity among reproductive-aged women worldwide. 100 Endometrial tissues can be shed from the uterus by retrograde menstruation to the extrauterine spaces, such as the peritoneal cavity and ovary, and implanted in them. These ectopically implanted tissues are believed to cause this disease.
Endometriosis is also a precursor to ovarian cancers with EnM and CCC subtypes. 101 The relationship between endometriosis and ovarian cancers has been studied extensively. For example, these subtypes of cancers are currently classified as type-I and type-II subtypes according to their responsiveness to estrogen. 102 Type-II is considered as a more aggressive disease, and its risk factors have been explored. 102 Moreover, this classification has been reexamined by analyzing the expression and mutation patterns of specific marker proteins. 103 These studies raise questions regarding the classical classification of endometriosis-derived ovarian cancers. The detailed mechanism underlying the conversion of endometriosis into certain types of ovarian cancer remains obscure. However, accumulating experimental data suggest that mutations in kinases, such as PI3-kinase, activate cellular signaling pathways in CCC cells. 104 The chromatin regulator gene ARID1A is inactivated by gene alterations in CCC cells.105,106 Some of these genetic changes can be found in endometrial tissues,105,106 suggesting that these gene aberrations contribute to the transformation of endometrial cells to cancer cells.101,107 As in the case of ovarian cancer, endometrial tissues can be exposed to hypoxia during the menstrual cycle. This environment may be a key factor regulating the pathology of endometriosis by modulating the function of macro-phages, 108 proangiogenic factors, 107 hormone expression, 107 prostaglandins, 109 and microRNAs. 110
Generally, TF expression in glandular epithelial cells within the normal endometrium is low compared with that in decidualized stromal cells.
111
This TF expression can be enhanced, as F3 expression is increased in response to sex hormone exposure, and its expression level can therefore change during menstruation.
112
However, TF is highly expressed in glandular epithelial cells within endometriosis tissues, presumably in response to proinflammatory and hypoxic environments,109,113 suggesting that TF forms a complex with fVII and fX, thereby contributing to the malignant transformation of endometriosis cells (Fig. 4). PAR1 can be activated by thrombin within the endometriotic stroma in response to recurrent bleeding from highly vascu-larized lesions.114–116 Thrombin may stimulate endometriotic stromal cells to increase the transcript levels of TF.
114
Furthermore, PAR2 can be activated via TF-fVIIa activity and contribute to the progression of endometriosis.114–116 Given that TF is highly expressed in CCC cells and can enhance the malignant phenotype by promoting cell motility and invasiveness via PAR1,
17
it is feasible that the activity of the TF-PAR axis is enhanced as the disease progresses under repeated hypoxic and inflammatory peritoneal conditions, resulting in the transformation of endometriosis into CCC (Fig. 4). In contrast to CCC cells, ectopic synthesis of fVII in endometriosis cells has not been reported; therefore, the aberrant expression of fVII during the disease process and its contribution to endometriosis remain unclear.
Hypothetical model of the TF-fVIIa-dependent transformation of endometriosis into CCC. Repeated hypoxia and/or inflammation conditions during menstruation cycles could upregulate the expression of TF and fVii in epithelial cells. TF-fViia activity on the cell surface is expected to activate PAR2. fX may participate in TF-FVIIA-FXA ternary complex formation, leading to activation of PAR1 (potential fX involvement is designated as a dotted line). These events could facilitate the transformation of endometriosis into CCC. Activation of PAR1 by thrombin because of recurrent bleeding may contribute to this transformation process.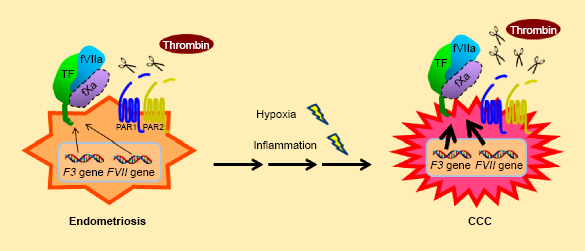
Therapeutic Potential
As previously reviewed, multiple therapeutic strategies to block TF-fVIIa activity have been tested in breast cancer models. 6 Briefly, the methodologies can be classified into three categories. First, the use of antibodies targeting TF to block procoagulant and cellular signaling activities. Second, suppression of the transcriptional activation of the F3 and FVII genes in cancer cells by targeting transcription factors aberrantly activated within the tumor microenvironment. 20 Conventional small molecule inhibitors targeting HIFs 72 may be applicable for the inhibition of inducible FVII activation, which is dependent on HIFs. However, this is not specific to FVII activation associated with the Sp1 transcription factor, as these drugs are expected to inhibit conventional ARNT-dependent genes. 20 Inhibitors that specifically block inducible FVII activation have not been reported to date. Third, TF can be used as a target of drug delivery. Cytotoxic drugs conjugated with inactive fVII accumulate in TF, expressing tumor tissues to a greater extent than their fVII-free counterparts, thereby increasing therapeutic efficacy. 6
Icon is an immunoconjugate molecule composed of enzy-matically inactive fVII and IgG1 Fc.31,117 Because TF can be overexpressed in ovarian cancer and endometriosis cells, icon is expected to accumulate on the surface of cancer cells overexpressing TF via fVII. Cell surface icon then kills these cancer cells by activating host immune response via the IgG1 Fc moiety without severe toxicity. Indeed, the icon strategy has been applied to the treatment of endometriosis in vivo 117 and ovarian cancer cells, in particular CCC cells, in vitro. 31 These studies indicate that this method is a promising therapeutic strategy for the treatment of aggressive gynecologic diseases.
Future Perspectives
The TF–fVII signaling pathways and associated phenotypes have been extensively studied using breast cancer cells, and a number of therapeutic strategies can be suggested. TF can be overexpressed in EOC cells. However, the understanding of ovarian cancer cell biology associated with aberrant expression of the cell surface TF–fVII complex has not kept up with that of breast cancer cells. Thus, future active investigation in this field may lead to the identification of signaling pathways associated with TF–fVII complex formation as promising targets for ovarian cancer treatment. The proportional relationship between the incidence of VTE and the morbidity and mortality of ovarian cancer patients has been reported; therefore, a detailed understanding of the molecular mechanisms connecting TF–fVII signaling, VTE, and disease prognosis may lead to the development of novel therapeutic strategies for ovarian cancers, particularly CCC. The contribution of the TF–fVII complex to the biology of endometriosis is also intriguing. Dissection of the TF–fVII pathway may improve our understanding of the molecular mechanism underlying the transformation of normal endometrial cells into CCC cells. Ovarian cancer cells can ectopically express fVII, particularly when cells are exposed to hypoxia. It would be interesting to investigate how the TF–fVII complex, composed of inducibly expressed fVII, functionally differs from that composed of exogenously supplied fVII. These studies may provide a rationale to explain why ectopically synthesized fVII is necessary for cancer progression. Moreover, the ectopic synthesis of fVII in endometriosis cells has not been reported. Thus, a more detailed dissection of the molecular mechanism of fVII expression and how it affects ovarian cancer and endometriosis is vital for translating our current knowledge into the development of therapeutic strategies for the treatment of ovarian cancers with a poor prognosis. Clinically, as in the case of breast cancer cells, the identification of relationships between ectopic fVII expression and various clinical parameters of EOC, such as chemosensitivity, relapse, and survival rate, could predict which patients may benefit from TF-fVII-targeted treatment. In any case, elucidating the biology of procoagulant activity and signaling pathways associated with the TF–fVII complex may lead to new therapeutic and diagnostic opportunities for problematic gynecological disorders, such as EOC and endometriosis.
Author Contributions
Conceived and wrote the manuscript: SK. Provided comments and wrote part of the Relationship between TF-fVIIa Signaling and Endometriosis section: YM. Approved the final version of the manuscript: SK and YM.