
Abstract
Many attempts have been made to identify objective molecular biomarkers to diagnose and prognosticate oral epithelial dysplasia (OED) because histopathological interpretation is subjective and lacks sensitivity. The majority of these efforts describe changes in gene expression at protein level in OED as determined by immunohistochemistry (IHC). However, the literature on these putative markers of oral cancer progression is vast and varied. The main purpose of this article is to review current knowledge on biomarkers of protein expression for OED by IHC approaches. We further discuss these findings in terms of the proposed essential hallmarks of cancer cells to better understand their role in oral oncogenesis.
Introduction
The concept of a step-wise transition from oral potentially malignant lesions (OPMLs) to oral squamous cell carcinoma (OSCC) is well-established, 1 but it can be difficult to predict if and when an OPML will undergo full transformation and result in a tumor. 2 The presence of oral epithelial dysplasia (OED) in OPMLs is generally accepted as one of the most reliable predictors of malignant development; 3 however, histopathologic diagnosis is subjective and lacks sensitivity. There is no agreement on which features of dysplasia are important in predicting progression. In addition, there is both inter- and intra-observer variation in interpreting the degree of epithelial dysplasia.4–6 Therefore, several studies have been conducted to identify objective molecular biomarkers to diagnose and prognosticate OED using different types of markers such as loss of heterozygosity, DNA ploidy, telomerase activity, methylation, and gene expression analysis. There are numerous reports describing changes in gene expression at the mRNA and protein levels in OED as putative markers of oral cancer progression. Most of these studies used immunohistochemistry (IHC) for protein detection. IHC examination has the potential to be a useful tool for diagnosing OED as it does not require specialised equipment, does not involve lengthy laboratory manipulation of tissue samples, permits evaluation of cell morphology during examination, and can be applied to archival specimens. Although the interpretation and quantification of immunohistochemistry results are governed by many factors, such as examiner experience, processing of tissue, antibody specificity, antibody dilution, and detection systems, 7 improvements in automated analysis with wider applicability could lead to more standardization. 8 IHC is currently being used for diagnosis of other tumors such as breast lesions 9 and bone tumorlike lesions. 10 Hence, if appropriate candidate markers can be applied, IHC can be used in routine diagnostic protocols of OED. Currently however, the literature is overwhelmed with IHC studies with no general agreement regarding the use of tissue markers in routine diagnosis of OED. The main purpose of this article was to review the current knowledge on biomarkers of protein expression for OED by IHC approaches to better understand their role in oral oncogenesis and to set these against the hallmarks of cancer as proposed by Hanahan and Weinberg in their seminal papers.11,12
OED biomarkers and hallmarks of cancer cells
Oral carcinogenesis is a highly complex, multistep process involving accumulation of genetic alterations that lead to the induction of proteins promoting cell growth (encoded by oncogenes), as well as the loss of proteins restraining cell proliferation (encoded by tumor suppressor genes). 1 The molecules involved in these processes may therefore provide markers for the early detection of malignant transformation. Proteins investigated in OED by IHC belong to different family groups, including: growth factors, growth factor receptors, cell-cycle proteins, proliferation markers, cell-cycle inhibitors, apoptotic factors, angiogenic signals, and cell adhesion molecules, among others. Figure 1 summarizes the pattern of protein expression and whether expression increases or decreases during oral carcinogenesis. Some proteins showed irregular expression patterns. Hanahan and Weinberg proposed six essential hallmarks of cancer cells that distinguish them from their normal counterparts.11,12 The hypothesized hallmarks include: self-sufficiency in growth signals, insensitivity to antigrowth signals, avoidance of apoptosis, resistance to cell senescence, development of new vascular supplies, and invasion and metastasis. Dysplastic epithelial cells are predisposed to develop these phenotypes as they progress toward cancer. Figure 2 summarizes how protein expression alterations identified in our review contribute to the acquisition of the essential hallmarks of oral cancer. The role of each marker in oral carcinogenesis is discussed below.
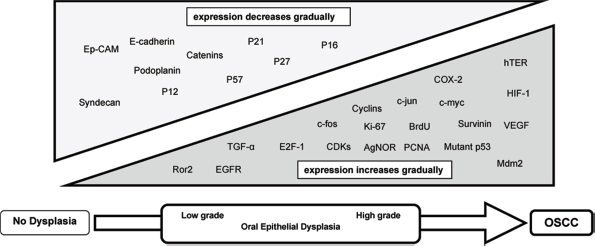
Pattern of protein expression during oral carcinogenesis.
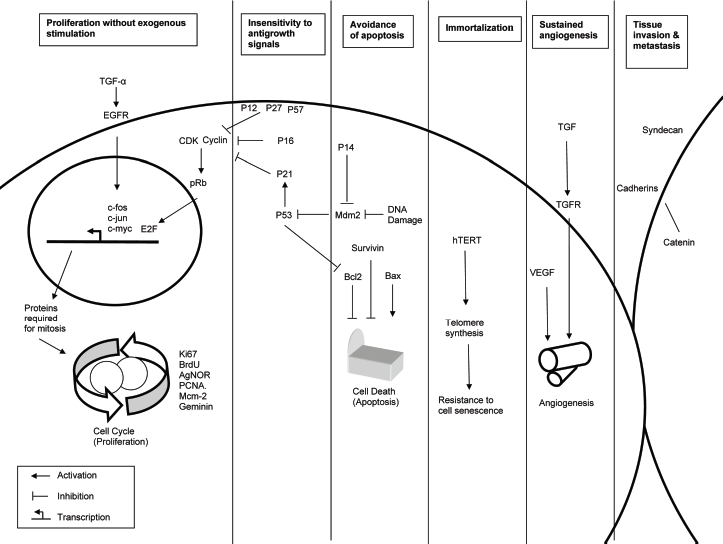
Schematic representation of contribution of protein alterations to the acquisition of the essential hallmarks of oral cancer.
Proliferation without exogenous stimulation
Normal cells require extracellular growth signals to proliferate, while cancer cells can grow without exogenous stimulation. 11 This can occur through one or more of the mechanisms described below.
Over-expression of extracellular growth signals
Growth factors are extracellular signals that play an important role in the regulation of cell growth, proliferation, and differentiation by binding to their receptors on the cell membrane. 11 Mitosis in a variety of mammalian epithelial cells is stimulated by epidermal growth factor. Transforming growth factor-alpha (TGF-α) is an epidermal growth factor family protein. 13 It has been found that the intensity of immunohistochemical expression of TGF-α increases progressively as dysplasia advances from low grade to high grade, reaching its highest level in oral carcinoma. 14 The level of expression of TGF-α oncoprotein in dysplastic oral leukoplakia was clear when compared with adjacent histologically normal mucosa. 15 Similar results were found at the mRNA level. 13 Additionally, Smad proteins that play a pivotal role in intracellular signaling of the TGF superfamily were observed, in an ascending order, in normal oral mucosa, mild dysplasia, moderate to severe oral epithelial dysplasia, and OSCC. 16 These data support a role of TGF-α expression in the progression to oral cancer.
Alteration of the receptors for growth signals
Exogenous growth signal action is transmitted into the cell by binding to a specific transmembrane tyrosine kinase receptor that subsequently activates multiple intracellular signaling pathways, leading to cell proliferation. 11 Many studies have shown early over-expression of epidermal growth factor receptors (EGFRs) in OED14,17–21 and a positive correlation with the severity of dysplasia.14,22 EGFR family members include c-erbB1 (Her1), c-erbB2 (Her2-neu), c-erbB3 (Her3), and c-erbB4 (Her4). These subclasses have been studied in OED.20,23 Several reports demonstrate that c-erbB-2 expression increased progressively from non-dysplasia to dysplasia and OSCC.24–27 In addition, a positive association between cytoplasmic c-erbB2 and the Ki-67 proliferation index was found in OED. 28 Although a non-significant difference in c-erbB-2 expression between normal, OED, and OSCC was observed in one study, 29 dysregulation of EGFR expression may be important in the development of oral cancer, and its evaluation in OED may assist in the diagnosis of these lesions.
Ror2 is a member of the Ror family of receptor tyrosine kinases which bind with a member of the Wnt family (Wnt5a), activating a planar cell polarity pathway (Wnt/JNK pathway) to regulate cell polarity and cell movement. Kobayashi et al 30 examined the expression of this receptor in OED and OSCC and found that the number of cells expressing Ror2 increased with the progression from dysplasia to malignancy.
Dysregulation of intracellular pathways that translate growth signals into mitosis
In addition to the dysregulation of growth factor receptors and their ligands, direct activation of the intracellular signalling pathways may also occur in carcinogenesis, allowing cells to proliferate without exogenous stimulation. 11
Cyclins and cyclin-dependant kinase. The cell cycle is divided into four phases: G1 (first gap), S (DNA synthesis), G2 (second gap), and M (mitosis). Near the end of G1, cells reach a key restriction point at which they either enter S phase and complete the cycle or exit and become quiescent. 31 Progression of mammalian cells from quiescence to mitosis is tightly controlled by regulatory proteins such as cyclin and cyclin-dependent kinase (CDK). 32 These two molecules form a complex which is responsible for the phosphorylation and inactivation of retinoblastoma protein (pRb). Following the inactivation of pRb, a transcription factor known as E2 promoter-binding factor (E2F) is released, which mediates the expression of significant cell-cycle control genes to allow progression into the remainder of the cell cycle. 33 Deregulation of expression of these proteins can result in loss of control of normal cell growth, leading to cancer development. Various types of cyclins and CDKs are synthesized during cell cycle events.
It has been shown that CDK2 was not detectable in normal tissue or mild or moderate OED, but it was clearly observed in severe dysplasia and OSCC.32,34 Similarly, CDK4 expression has been found to increase as dysplastic tissue progresses to OSCC with no expression in normal tissue. 32
Various studies have demonstrated that activation of cyclins can lead to loss of growth control underlying the development of various forms of human cancer. 31 IHC studies in OED and OSCC showed increased cyclin A protein expression with increasingly severe grades of dysplasia, with the highest levels in OSCC as compared with normal controls.34–36 Conversely, Thomson et al 37 showed that cyclin A protein expression in invasive OSCC was lower than those recorded for either severe dysplasia or carcinoma in situ. In OED, there was strong correlation between cyclin A and proliferation markers such as Ki-67 36 and bromodeoxyuridine (BrdU). 38
Several studies agree that cyclin D expression is seen late in the development of oral cancer because it is expressed predominantly in OSCC, and in few cases of moderate to severe dysplasia but not in normal epithelium and mild dysplasia.32,34,39,40 However, Rousseau et al 41 found a similar proportion of cyclin D expression in all grades of dysplasia and OSCC. Oliver and MacDonald found high levels of cyclin D expression in OED, which was positively correlated with Ki-67. 42 Another study sh owed that cyclin D1 expression was associated with loss of pRb expression, and its expression in OSCC was significantly higher compared with its expression in OPMLs and histologically normal oral tissue. 33
Cyclin E has also been examined in OED. It has been found that this subclass of cyclin was detectable in 57.1% of severe dysplasia and 62.8% of OSCC, but was not observed in normal epithelium or mild dysplasia.32,34
The progressive increase of CDKs and cyclins during oral carcinogenesis suggests that these markers can be used as predictive indicators of OED.
pRb and E2F. Quantitative immunohistochemical results for pRb in OED have shown that its frequency increased with dysplasia progression.40,43 In contrast, a significant loss of pRb was observed in the transition from hyperplasia to dysplasia. 33 Only one study investigated the expression of E2F-1 proteins in OED. The study showed that E2F-1 was significantly increased in both oral dysplasia and OSCC compared with normal epithelium and this increase was associated with increased cyclooxygenase 2 (COX-2) expression. 44 Additional studies are needed to confirm the usefulness of pRb and E2F in OED diagnosis.
Transcription factors. Another mechanism by which cells can proliferate without exogenous stimulation is through alterations in the expression of transcription factors. Transcription factors are proteins that bind to gene promoters and regulate transcription. Oncogenes that produce altered forms or excessive quantities of specific transcription factors have been detected in a broad range of human cancers. 45
The oncoproteins c-fos and c-jun have been implicated in the development of epithelial dysplasia as they are components of the transcription factor activating protein-1 (AP-1). 39 Sachdev et al 46 showed gradual increase of c-fos expression from normal mucosa to dysplastic lesions to OSCC, suggesting early activation of this protein in oral carcinogenesis. Similar results obtained from Turatti et al 39 showed gradually increased expression from mild dysplasia to moderate dysplasia to OSCC, but intense expression was found in normal tissue. Nuclear expression of c-jun protein has not been detected in non-dysplastic oral tissues, while the intensity of staining increased linearly with increasing grades of dysplasia showing very strong staining in carcinoma samples.39,47
Another transcriptional factor which has been implicated in human cancer, but is not well-studied in OED, is c-myc. Overexpression of c-myc has been observed in dysplastic epithelium and with lower intensity in non-dysplastic oral tissue. 17 Bmi-1, a c-myc co-operating oncogene in murine lymphomagenesis, was found to be expressed at a very early stage in oral carcinogenesis, including that of mild epithelial dysplasia. 48 Cytoplasmic expression of cancerous inhibitor of protein phosphatase 2A (CIP2A), which stabilizes the c-myc protein by inhibiting its degradation, was found to be highly expressed in severe dysplasia compared to mild dysplasia, while nuclear expression was found to be lower in severe dysplasia than in mild dysplasia. 49 The results of these studies support that over-expression of transcription factors may contribute to the multistep nature of oral carcinogenesis.
Proliferation markers. Multiple studies have examined several indicators of cellular proliferation in OED using IHC. Most of these studies demonstrated an increased rate of proliferation in OED by examining Ki-67,50–52 BrdU, 53 silver-binding nucleolar organizer region (AgNOR),54–56 or proliferating cell nuclear antigen (PCNA).32,57 Despite the non-significant prognostic value for Ki-67 in OED progression, 58 proliferation indices can provide further objective measurements of OED.
Minichromosome maintenance proteins (Mcm 2–7) are essential for eukaryotic DNA synthesis. Mcm-2 was used recently as a new marker for cell proliferation. Immunohistochemical examination of OED samples showed a greater frequency of Mcm-2 expression in surface layers of moderate/severe dysplasia and OSCC compared to benign keratosis/mild dysplasia. 59 Other studies found that expression of Mcm-2, geminin (another novel proliferation marker), and Ki-67 increased progressively from normal to OED and OSCC.50,58 Mcm-2 is seen to be a useful prognostic marker for OED, as its expression was markedly higher in oral dysplastic lesions with malignant progression compared to non-progressive lesions.50,58
Insensitivity to antigrowth signals
Progression of the cell cycle is typically restricted by cell-cycle inhibitory signals. Disruption of inhibitory molecules can lead to uncontrolled cell proliferation and tumor formation. 31 The multistep process of oral carcinogenesis likely involves the functional loss of these proteins through various mechanisms, including mutation, loss of heterozygosity, or hypermethylation. 60
Alteration of the tumor suppressor gene TP53 is one of the most common genetic abnormalities in human malignancy including oral cancer, 61 and it is considered one of the strongest predictors for cancer development. 62 Based on the fundamental assumption that mutant p53 protein has a prolonged half-life, it can be detected by IHC; the half-life of wild-type p53 protein is too short to permit detection.61,63,64 Using IHC, p53 protein expression has been detected in 10–80% of OED.64–70 Some investigations revealed that the TP53 mutation is an early event in oral carcinogenesis,65,66 while others have shown that it is a late event.61,64 Several studies linked p53 detection with greater risk for malignant progression,61,64,65 particularly when it was detected in basal or parabasal layers.62,71–74 Although most previous investigations revealed a direct correlation between p53 accumulation and histologic grade of dysplasia,66,68,73,75 no such relation was found in one study. 67 The limitation of using this molecular marker as the only evidence of risk assessment is that IHC can detect the overexpression of both mutant and wild-type p53 proteins.
p63 and p73 are members of the p53 family. It has been found that p63 and p73 expression was higher in oral cancer and dysplasia than in normal tissue.76–78 A gradual increase in the extent of p63 staining was observed as the lesions advanced from hyperplasia to dysplasia and ultimately to carcinoma. An association between tumor suppressor protein mammary serine protease inhibitor (maspin) was also observed. 79 However, Bortoluzzi et al 80 found that p63 and Ki-67 showed different patterns of staining in OED, suggesting that p63 may not be associated with proliferation. In a 5-year follow-up study, a subset of OED showing p63-positive staining underwent malignant transformation. 77
Another key regulator of p53 protein is mouse double minute 2 (MDM2). Increased expression of this protein has been observed as OED progressed to cancer.58,81,82
p16 proteins normally act to block cell cycle progression at the G1 to S transition; therefore, inactivation of the p16 gene enables unregulated cell growth. 60 Several studies have investigated hypermethylation of p16 in OED,83,84 but few studies have examined IHC expression of this protein. Shintani et al 32 showed that p16 immunostaining was detectable in normal epithelium, but its expression decreased in dysplasia and was almost absent in OSCC. However, the results of Bradley et al 85 suggest that p16 immunohistochemistry is not helpful for differentiating dysplastic from non-dysplastic oral mucosal biopsies, and thus is not a reliable biomarker for use in routine clinical practice.
The CDK inhibitor p27 plays an essential role in G1 arrest induced by TGF-β in normal cells. p27 controls cell proliferation by binding and inhibiting G1 cyclin-CDK complexes and negatively regulating progression through G1 and S phases of the cell cycle. 36 Various studies have shown that p27 is associated with loss of expression with disease progression from normal to OED and OSCC.32,36,40,58,86–88 Skp2 is a member of the F-box family and has been implicated in the degradation of several key regulators of mammalian G1 progression, including p27. Increased levels of Skp2 protein were associated with reduced p27 in a subset of oral epithelial dysplasias and carcinomas compared with normal epithelial controls. 87
Another CDK inhibitor which has been investigated in OED is p21. The expression of this tumor suppressor gene was detectable in most normal oral epithelia, while it was gradually lost in transition from dysplasia to OSCC.32,58 However, Schoelch et al 40 showed that expression of p21 and Ki-67 increased with disease progression. After an average follow-up period of 3.5 years of oral verrucous leukoplakia, a significant difference in the frequency of OSCC progression/recurrence was noted in lesions bearing aberrant immunoreactivity of either p53 or p21 compared with lesions with negative immunoreactivity. 89
Parallel results were seen for p12 and p57. Positive staining of p12 was observed in 100% of cases of normal mucosal epithelium, mild dysplasia, and moderate dysplasia compared to 85.7% and 28.2% of severe dysplasia and OSCC respectively. 32 p57 expression was found to be decreased in oral leukoplakia with moderate or severe dysplasia, and further decreased in OSCC. 90
The results obtained from the immunohistochemical studies discussed above indicate a possible role for cell-cycle inhibitor loss in malignant transformation of oral mucosa.
Avoidance of apoptosis
Normal cells have the ability to destroy themselves through the process of apoptosis if irreparable DNA damage occurs during cell division. This is a natural defence mechanism to prevent the propagation of genetically damaged cells. 11 Apoptosis is controlled by a cascade of signaling pathways, including pro-apoptotic molecules such as p53 and bax, and anti-apoptotic proteins such as bcl-2, mdm2, and survivin. Cancer cells must find ways to evade apoptosis if they are to survive. They do this by shifting the balance between apoptotic factors.
The bcl-2 family is divided into two groups: anti-apoptotic proteins, such as bcl-2 and bcl-xL, and pro-apoptotic proteins, such as bax and bak. 91 Apoptosis-associated proteins have been shown to be altered in variable patterns in both dysplastic and malignant oral lesions. 92 It has been found that bcl-2 oncoprotein is up-regulated in dysplastic epithelium 93 and its expression is directly proportional to the degree of epithelial dysplasia. 94 In contrast, Loro et al 91 showed that bcl-2 protein were markedly decreased in the basal layer of moderate and severe OED compared with the basal layer of normal oral epithelium, while Sousa et al 95 found over-expression of Bax in OED.
Survivin plays an important role in apoptosis inhibition. Lin et al 96 reported a high incidence of survivin protein expression in OED and OSCC samples, but not in adjacent normal oral mucosal tissues. A similar pattern was found at the mRNA level. 97 In an important follow-up study (13-292 months), Lo Muzio et al 98 showed that survivin expression was seen in 33% of OPMLs that did not progress to cancer, compared to 94% of OPMLs that eventually progressed to OSCC, suggesting the use of this marker for the identification of OPMLs at higher risk of progression. 98
Cyclooxygenases are enzymes that mediate the production of prostaglandins from arachidonic acid, which have been shown to decrease apoptosis rates in oesophageal adenocarcinoma cells. 99 Protein expression by IHC in normal oral epithelium and in different stages of oral cancer progression showed a gradient increase of COX-2 staining.44,100,101 However, one study showed that COX-2 expression was not a reliable marker to select OPMLs at risk for development of OSCC after 6-225 months follow-up. 102 Recent evidence suggests that the coordinated function of cytosolic PLA-2 (cPLA-2) and COX-2 in the arachidonic acid pathway may contribute to the process of carcinogenesis in various tissue types. 101 Expression of cPLA-2 in normal oral epithelia was significantly lower than in oral dysplasia and OSCC. 101
Heat shock proteins (HSP) are synthesized by cells in response to a variety of stress conditions, including carcinogenesis. 90 Experimental evidence suggests that HSPs may be associated with tumor progression by inhibiting apoptosis. 103 A study conducted on oral leukoplakia revealed that HSP60 expression increased in moderate dysplasia compared tomild dysplasia. However, no significant difference was found between leukoplakia with severe dysplasia and leukoplakia with moderate dysplasia, or between leukoplakia with severe dysplasia and OSCC. 90 Another study found a non-significant difference for HSP27 cytoplasmic expression between oral leukoplakia with and without epithelial dysplasia. 103 A significant correlation between HSP70 expression and dysplasia severity has been observed.103,104 A 5-year follow-up study showed that the median transition time (premalignancy to malignancy) was significantly shorter in cases showing overexpression of HSP70 protein. 104
Metallothionein belongs to a group of low-molecular weight proteins characterized by high levels of cystein, which are bound to metal ions. It plays a role in protecting against oxidative damage caused by free radicals, inhibition of cell apoptosis, and carcinogenesis. Johann et al 105 showed that the expression of this molecule was significantly higher in moderate dysplasia when compared to normal oral mucosa, hyperkeratosis, and mild dysplasia. P-Akt controls a variety of critical cellular pathways during the carcinogenic process, including those leading to apoptosis inhibition. A significant difference in the immunoexpression of metallothionein and p-Akt was found in OSCC samples compared with normal or dysplastic epithelial lesions. 106
Immortalization
Normal cells have a limited ability to proliferate as they cease growing after numerous cell replications, reaching a state known as ‘senescence’. Malignant cells acquire the ability to escape from cellular senescence and become immortalized by lengthening their telomeres. 11 Human telomeres consist of tandem arrays of repeated TTAGGG hexamers located at the end of chromosomes, which serve to prevent the end-to-end fusion of chromosomal DNA. 107 With each successive round of cell replication, telomeres become shorter, a phenomenon that eventually results in senescence and permanent growth arrest. Telomeres are synthesized by a ribonucleoprotein enzyme called telomerase, which maintains telomere length in human germline and stem cells. The activity of telomerase increases frequently in human cancer. 108 Telomerase consists of a catalytic protein subunit, human telomerase reverse transcriptase (hTERT), and an RNA template (hTR). Enzyme activity is very closely correlated with the level of hTERT expression. 109 Few studies have detected hTERT protein by IHC in dysplastic and neoplastic oral epithelia. Kumar et al 110 and Chen et al 111 found that the expression of hTERT increased significantly from normal through OED to OSCC. At both the mRNA and protein levels, the expression of hTERT in OED and in OSCC was markedly higher than in normal tissues.109,112 Thus overexpression of immortalization marker may occur early in oral carcinogenesis and may be used as a diagnostic indicator in OED; however, its prognostic value remains unknown.
Sustained angiogenesis
Angiogenesis is a crucial event for the development, progression, and metastasis of malignant tumours because tumor growth requires an adequate blood supply to provide nutrients and eliminate metabolic waste products. 113 Malignant tumours acquire this ability by increasing expression of pro-angiogenic factors such as vascular endothelial growth factors (VEGFs), fibroblast growth factors (FGFs), and heparin-binding protein 17 (HBp17).
VEGFs are a family of potent pro-angiogenic factors that stimulate endothelial cell proliferation. Recent studies have demonstrated up-regulation of VEGF in oral tissues during the transition from normal to dysplasia and then OSCC.114,115 In another study, significant correlation was found between the severity of OED and both nitric oxide synthase-2 (NOS2) and VEGF. 113 Lopez de Cicco et al 116 showed notably higher expression of VEGF-C and furin in most dysplastic and cancerous lesions than in normal tissue. A contradictory result indicated an inverse relationship between VEGF expression and oral cancer progression. 117
Begum et al 115 showed that strongest expression of FGF-2 and HBp17 was observed in severe dysplasia and OSCC, while another study concluded that down-regulation of FGF-1 is consistent with loss of differentiation in dysplasia. 24
Li et al 118 showed that micro-vessel density, VEGF, bFGF, and platelet-derived endothelial growth factor (PDGF) did not differ significantly between normal oral mucosa and epithelial dysplasia, but was significantly increased in tumour tissues.
Hypoxia-inducible factor 1 (HIF-1) is a key regulator of the cellular response to hypoxia. Recent studies have shown that HIF-1 a is associated with activation of more than 60 hypoxia-inducible genes such as genes encoding for VEGF. Lin et al 119 found that HIF-1a expression was found to be increased significantly from normal through mild, moderate and severe OED to OSCC. Similarly, Tilakaratne et al 120 showed that HIF-1 a was up-regulated at both the protein and mRNA levels in oral submucous fibrosis and the correlation with the severity of epithelial dysplasia was statistically significant.
Podoplanin is specifically expressed in lymphatic endothelial cells but not in blood endothelial cells, and has been utilized as a specific marker for recognizing lymphatic vessels. Kawaguchi et al 121 found that as the degree of dysplasia increased, there was an increase in podoplanin expression. They also showed that OPMLs with positive podoplanin expression were more likely to progress to oral cancer than those with negative podoplanin staining. 122 These data show that podoplanin merits further investigation as a promising biomarker that can discriminate between lesions at risk of developing OSCC, regardless of histologic evidence of dysplasia.
Tissue invasion and metastasis
Oral cancer is characterized by local invasion across the basement membrane and a tendency for dissemination and migration to other locations. These abilities are fundamental behaviors of cancer cells. 123 Invasion and metastasis involve complex multistep processes that lead to alterations in cell-cell and cell-matrix interactions. Typically, these changes are accompanied by loss of expression of cell adhesion molecules, even in the very early stages of cancer development.
Several immunohistochemical studies have shown that the cadherin/catenin cell adhesion system is often disrupted in OED. Cadherins are a large family of cell-surface glycoproteins that act as intercellular adhesion molecules and are named according to their tissue specificity (E-cadherins, epithelial; N-cadherins, neural; P-cadherins, placental). 124 Cadherins have the potential to be significant in the pathogenesis of OSCC. Recently, E-cadherin expression was found to decrease progressively with increased grade of dysplasia. 125 Loss of E-cadherin expression was observed in dysplastic oral mucosa adjacent to tumours.21,124 An irregular pattern was noticed for P-cadherin expression as it was found to be up-regulated in severe dysplasia and down-regulated in carcinoma insitu compared to normal epithelium. 123 Furthermore loss of E-cadherin expression could not select oral lichen planus lesions at risk of malignant transformation. 102
The adhesive function of E-cadherins depends on the association with cytoplasmic proteins, known ascatenins, that bind E-cadherins to the cytoskeleton. The family of catenins include alpha-, beta-, and gamma-catenins. 123 Beta- and gamma-catenin expression was found to be reduced in dysplasia and was generally more reduced with increased degree of dysplasia. 126 Nuclear expression of beta-catenin has been found to be more prominent in dysplastic than in nondysplastic leukoplakia. 127 No statistically significant difference was found in catenin expression between progressive and non-progressive lesions. 126
Syndecans are cell-surface molecules that belong to the heparansulfate proteoglycan family, which interact with extracellular matrix components, other cell surface molecules, and growth factors. 128 Early results demonstrated loss of syndecan-1 in OED 129 and a negative association with the extent of epithelial dysplasia; 130 however, another study showed no significant difference in syndecan-1 expression between mild, moderate, and severe dysplasia or between dysplasia and OSCC. 128
In addition to the disruption of cell-cell interactions, impaired distribution of basement membrane also plays an important role in invasion and metastasis. The most common basement membrane components include collagen IV and laminin. Cellular localization of type-IV collagen alpha chain distribution during oral carcinogenesis demonstrated specific alterations at the basement membrane, suggesting a crucial role for type-IV collagen alpha interruption in progression of dysplastic cells into the extracellular compartment. 131 Others have concluded that the loss of continuity in laminin and collagen-IV expression is associated with the evolution of dysplasia to malignancy.125,132
Parikka et al 133 observed down-regulation of collagen XVII (a hemidesmosome component) in basal cells in mild dysplasia and up-regulation in suprabasal keratinocytes in moderate and severe dysplasia as well as in the central cells of OSCC. Vaananen et al 134 demonstrated loss of collagen XVIII in the basement membrane of severe epithelial dysplasia and invasive areas of OSCC compared to continuous signal in mild epithelial dysplasia.
Integrins are heterodimeric cell surface glycoproteins that recognize a variety of extracellular ligands, such as laminins, fibronectin, collagen, and vitronectin. These molecules play a crucial role in the maintenance of tissue integrity and in the regulation of cell proliferation, growth, differentiation, and migration. Abnormal localization of integrins has been observed in OED.135,136
Moesin is a member of a three molecule family (moesin, ezrin, and radixin) which links actin filaments of the cell surface structure to the cell membrane. Kobayashi et al 137 showed variation in moesin immunolocalization in OED and OSCC but not in expression levels.
CD44 variant forms are a family of transmembrane glycoproteins encoded by a single gene. Bahar et al 138 found marked down-regulation of CD44v6 expression in most cases of severe dysplasia and OSCC. Expression of the variant exons v7, v8, and v10 was significantly down-regulated in cancer compared to hyperplastic areas and in dysplastic foci. 139 In contrast, Rautava et al 140 showed heterogenous staining patterns in oral dysplasia and carcinoma.
Keratins are proteins that constitute the intermediate filament cytoskeleton of epithelial cells, with approximately 20 known subtypes. 63 Changes in the type or distribution of keratins have been observed during oral carcinogenesis.52,141–146
The epithelial cell adhesion molecule (Ep-CAM) is a type I transmembrane glycoprotein involved in cell signalling, migration, proliferation and differentiation, cell-cycle regulation, and cancer metastasis. Hwang et al 147 revealed that Ep-CAM expression decreased significantly from normal and mild OED through moderate and severe OED to OSCC.
Perlecan is a major heparansulfate proteoglycan. Over-expression of perlecan in dysplastic epithelial cells has been observed with increased localizationin the inter-epithelial space. 148
Matrix metalloproteinases (MMPs) are a family of zinc metalloenzymes that mediate the destruction of the extracellular matrix, allowing for tumorinvasion and spread. Increased expression of MMP-7 and perlecan has been detected in OED, suggesting that MMP-7 immunohistochemistry may be valuable for recognition of cell proliferation in dysplasia. 149 A follow-up study showed that MMP-1 and MMP-9 mRNA levels were significantly higher in OED that progressed to oral cancer compared with those that did not. 150 Other studies found that IHC expression of MMP-2 and MMP-9 was high in OSCC compared to dysplasia or normal oral mucosa and that this gain of expression was associated with collagen IV α chain loss.151,152
Aberrant activation of the Wnt signalling pathway is thought to initiate the epithelial-mesenchymal transition in OSCC, promoting tumor invasion and metastasis, and may also play an early role in carcinogenesis by supporting malignant transformation. 153 Our recent IHC work showed downregulation of the epithelial marker E-cadherin together with upregulation of the mesenchymal marker Vimentin during progression from normal to dysplasia and OSCC. 153 These findings provide clearly demonstrated the epithelialmesenchymal transition of oral epithelial cells during oral carcinogenesis; these results have been supported by other studies. 154 – 155
In other studies, we also demonstrated a trend toward impaired differentiation accompanied by an increase in sternness properties during the transition from normal through dysplasia to OSCC.156,157 Cancer stem cells have been defined as a very small subset of cells within the tumor population that constitute a reservoir of self-sustaining cells with the exclusive ability to self-renew and to cause the heterogeneous lineages of cancer cells that comprise a tumour. 158 These two distinct biological phenotypes are what make the cancer stem cell a prime candidate for initiation of recurrence. Premalignant conditions may serve as a model for the cancer stem cell concept and a high proportion of putative cancer stem cells in OED may correlate with a high grade of dysplasia and a high progression rate. 159 A better understanding of the expression of cancer stem cell markers during the evolution from normal to premalignant epithelial dysplasia and OSCC may aid in the development of more effective therapeutics against oral cancer. 160 Terminal differentiation of oral epithelial cells involves a highly coordinated sequence of events that reflect the several distinct epithelial layers. Growth suppression is a hallmark of terminal differentiation, and impaired cellular differentiation due to genetic or epigenetic alterations may preserve a state of enhanced cell proliferation and predispose tissues to the accumulation of cancer gene mutations that elicit neoplastic transformation. 161 Loss of normal patterns of differentiation is common to most cancer types and dysregulation of cellular differentiation genes may indicate more fundamental abnormalities in cancer cells. The developmental process of epithelial-mesenchymal transition has a plausible link with cancer stem cell properties162,163 and may partially account for our observation of decreased epithelial differentiation marker expression and elevated expression of stem cell markers.
In their recent updated review, Hanahan and Weinberg 12 suggested that the acquisition of the hallmarks of cancer is made possible by two enabling characteristics. First is the development of genomic instability in cancer cells. Evidence of defective DNA repair has been found in OED. 164 Immunoexpression of human MutL protein homolog 1 (hMLHl), a protein of the mismatch repair system, was shown to be decreased in lesions with lower degrees of dysplasia compared to lesions with more severe dysplasia. 165 A second enabling characteristic includes the inflammatory state of premalignant and malignant lesions where immune cells infiltrate and engage in extensive and dynamic crosstalk with cancer cells. This may serve to promote cancer progression through various means. 12
Other distinct attributes of cancer cells have been proposed to be functionally important for the development of cancer and have been considered to be emerging hallmarks of cancer. 12 The first involves altered cancer cell-intrinsic metabolism, either as a consequence or as a cause. The best-characterized metabolic reprogramming that distinguishes cancer from normal cells is aerobic glycolysis. CA9, a hypoxiarelated protein which is associated with aerobic glycolysis, was found to be an independent risk factor of malignant transformation of OED. 166 The second involves the ability of tumor cells to evade the host immune system. Cancer cells can evade the host's defence by being poor stimulators of immune cells as well as being poor targets. 167
Conclusion
Numerous IHC studies have aimed to find objective molecular biomarkers that can be used to detect OED with an increased risk of developing oral cancer. We have discussed the role of each of these markers in relation to the acquisition of the six hallmarks of cancer as they pertain to oral carcinogenesis. Although the literature on putative markers is vast, none have been translated into routine clinical practice. Significant effort is required in this field before any IHC markers are established based on strong evidence as a basis for clinical practice. Well-designed multicenter studies inclusive of large datasets supplemented by long-term follow up data are required before IHC can routinely serve as an alternative to subjective histopathological diagnosis of oral epithelial dysplasia.
Author Contributions
Conceived the concept: CSF. Analyzed the data: AAA. Wrote the first draft of the manuscript: AAA. Contributed to the writing of the manuscript: AAA and CSF. Agree with manuscript results and conclusions: AAA and CSF. Jointly developed the structure and arguments for the paper: AAA and CSF. Made critical revisions and approved final version: CSF. All authors reviewed and approved of the final manuscript. AAA and CSF.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.