
Abstract
In the current study, we used immunoprecipitation and immunoblotting to examine the levels and phosphorylation status of the insulin receptor-beta subunit (IR-β), as well as the down stream target in PI3K pathway, total PKB/Akt as well as their phosphorylated forms. The assessment of FFAs treatment showed no direct and significant effect on the PI3K stimulation, specifically the IR-β in primary hepatic control cells treated with insulin. Cells treated with either oleate or palmitate (360 μM) showed no statistically significant values following insulin stimulation (P > 0.05). To further investigate the effect of both FFAs and high insulin (1 μg), we examined the effects of oleate and palmitate at 360 μM concentration on IR-β as well as PKB. There was no significant difference in the total protein levels and their phosphorylated forms in cells treated with or without oleate or plamitate. Interestingly, IR-β tyrosine phosphorylation showed a similar insignificant effect
Introduction
The close association between the ongoing worldwide epidemic of obesity, insulin resistance and type 2 diabetes has been well documented.1–5 Free fatty acids (FFA) have been implicated as an important causative factor in the development of type 2 diabetes. 6 The FFA-induced impairment of glucose metabolism in muscle is well documented,7–15 but little is known about the mechanisms of the FFA-induced impairment of glucose metabolism and the impairment of the insulin signaling cascade in the liver.12,16
In the liver, FFA increase gluconeogenesis both
The present study was performed 1) to investigate, whether chronic treatment of hamster hepatocytes with FFA induces changes in insulin signaling and development of insulin resistance and 2) whether, chronic exposure of hamster hepatocytes with high dose of insulin and FFA leads to desensitization of the PI3K pathway.
Experimental Methods
Materials
Fetal bovine serum (certified grade), liver perfusion medium, hepatocyte wash medium, liver digest medium, and hepatocyte attachment medium were obtained from Life Technologies (Grand Island, NY). Fatty acid free bovine serum albumin, protein A-Sepharose, oleic acid and palmitate were from Sigma Chemical Co. (St. Louis, USA). IR-β antibodies were from Santa Cruz Biotechnology (Santa Cruz, CA). PKB total antibodies and phosphospecific phospho–-Serine473 PKB and IκBα were from Upstate Biotechnology (Lake Placid, NY).
Animal protocols
Male Syrian golden hamsters (
Liver perfusion and isolation of primary hamster hepatocytes
Male Syrian golden hamsters (
Lipid preincubation
Lipid containing media were prepared by conjugation of FFAs with BSA (4%), using a method modified from that described by Svedberg et al. 28 Briefly, FFAs were dissolved in ethanol and diluted 1:25 at 45 °C, each solution contained 20% (w/v) fatty acid-free bovine serum albumin. Solutions were filter-sterilized and diluted 1:4 with serum or serum free (SF-William's E) as appropriate. Control media were prepared similarly containing ethanol and BSA in the absence of lipid. Primary hepatocytes were incubated for 16 hrs in 10 mm dishes at 3 ml/well in the absence or presence of FFA.
Chemiluminescent immunoblot analysis
Treated cells were collected in lysing buffer (150 mM NaCl, 10 mM trishdroxymethyl amino ethane, pH = 7.4, 1 mM EDTA, 1 mM EGTA, 1% Triton x 100, 1%NP-40, 2 mM PMSF, 10 μg/ml aprotinin, 10 μg/ml leupeptin, 100 mM sodium flouride, 10 mM sodium pyrophoshate and 2 mM sodium orthovanadate). The total cell lysates were either used directly or after immunoprecipitation against a target protein, 29 were subjected to chemiluminescent immunoblotting for IR-β (Santa Cruz), total PKB/ Akt (Upstate) as well as their phosphorylated forms at serine 473. Following SDS-PAGE the proteins were transferred electrophoretically overnight at 4 °C onto nitrocellulose membranes or PVDF. 30 The immunoblots were probed with the indicated antiserum. Bands were visualized using an enhanced chemiluminescence detection reagent (Amersham Pharmacia Biotech), and exposed to Kodak Hyperfilm. Films were developed and quantitative analysis was performed using an Imaging Densitometer (FluorChem and Alpha Innotech Corporation).
Statistical methods
All results are expressed as means ± SE, Statistical calculations were performed using Student's t test. A value is considered significant at p < 0.05.
Results
Effect of oleate or palmitate on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in control primary hamster hepatocytes
The monounsaturated FFA, oleate (18:1n9), and the saturated FFA, palmitate (16:0). Figure 1 shows the effect of oleate and palmitate on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in primary hepatocytes from chow-fed hamsters. We have examined the levels and phosphorylation status of the insulin receptor–-beta subunit (IR-β), the down stream target in PI3K pathway, total PKB/Akt as well as their phosphorylated forms. Interestingly, oleate and palmitate treatment showed a negative effect on the PI3K stimulation in primary hepatic control cells treated with insulin. Cells treated with either oleate or palmitate showed no statistically significant values following insulin stimulation (P > 0.05).
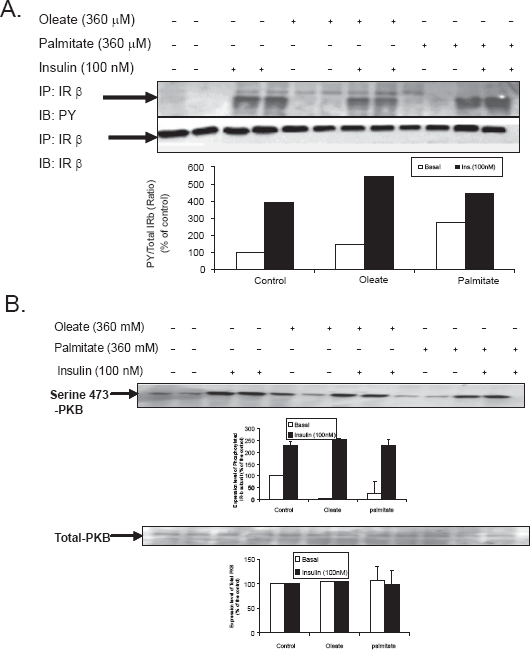
Effect of oleate or palmitate on IR-β subunit tyrosine phosphorylation phosphorylation in control primary hamster hepatocytes–-Primary hamster hepatocytes from chow-fed hamsters were treated for 10 min with (100 nM) insulin and 360 μM FFA for 16 hours. (A) Cells were lysed and immunoprecipitated using anti-IRβ mouse monoclonal anti body IgG followed by immunoblot with antiphosphotyrosine PY (99) mouse monoclonal antibody. (B) Membranes were reprobed using anti IR-β to verify mass. Each blot is a representation for one individual experiment (n = 3 experiments/group). T-test was conducted to compare FFAs treated versus appropriate basal or insulin stimulated, significant difference from STD (p < 0.05).
Effect of oleate or palmitate (360 μm) on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in control primary hamster hepatocytes in the presence of high insulin
To further investigate the effect of both FFA and high insulin levels (1 μg/ml), we examined the effects of oleate and palmitate (360 μM) on molecules of the insulin-signaling pathway, IR-β as well as PKB. Figures 2A, B show the effect of FFAs on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in control primary hamster hepatocytes. There was no significant difference in the total protein levels in cells treated with or without oleate or plamitate. IR-β tyrosine phosphorylation showed a similar negative effect in the cells treated with oleate and palmitate in comparison with control cells stimulated with insulin.
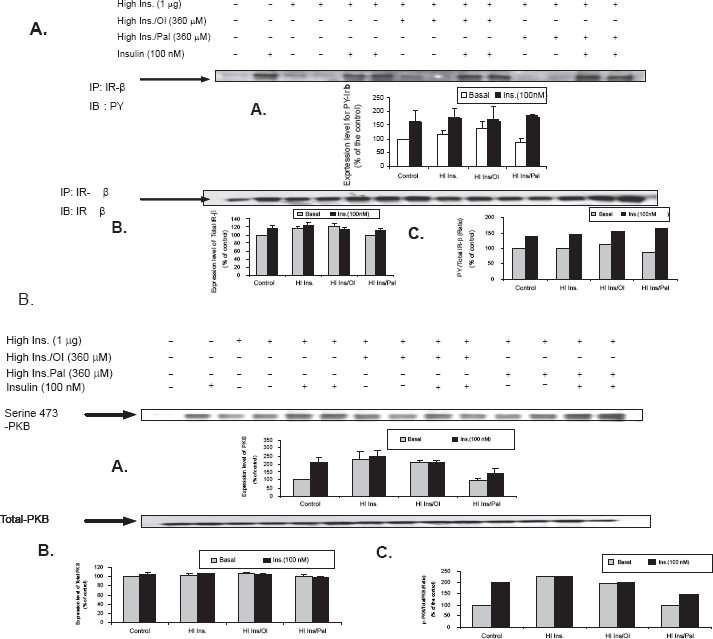
Effect of oleate or palmitate on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in control primary hamster hepatocytes in the presence of high insulin–-Figure 2A, B show a representative immunoblot and graphical representation of the effect of FFA on IR-β subunit tyrosine phosphorylation and PKB phosphorylation in control primary hamster hepatocytes. Each blot is a representation for one individual experiment (n = 3 experiments/group). T-test was conducted to compare FFA. To further investigate the effect of both FFAs and high insulin (1 μg/ml), we examined the effects of oleate and palmitate (360 uM) on molecules of the insulin-signaling pathway, IR-β as well as PKB. There was no significant difference in the total protein levels in cells treated with or without oleate or plamitate. IR-β tyrosine phosphorylation showed a similar negative effect in the cells treated with oleate and palmitate in comparison with control cells stimulated with insulin. T-test was conducted to compare FFAs treated versus appropriate basal or insulin stimulated, significance difference from STD (p < 0.05).
Effect of oleate or palmitate on IR-β subunit tyrosine phosphorylation in fructose fed primary hamster hepatocytes
Figure 3 shows the effect of oleate or palmitate on the PI3-K pathway and insulin signaling cascade in primary hepatocytes from fructose fed hamster. Phosphorylation of the IRβ subunit was assessed in cell lysates by first immunoprecipitating with the IRβ subunit followed by blotting for tyrosine phosphorylation. There was no additional effect above basal insulin signaling/phosphorylation upon treatment with either oleate or palmitate (Fig. 3A). The mass of IR for the same experiment was also verified (Fig. 3B) and this showed no significant change in IR mass suggesting that the lack of effect of FFAs on the PI3K pathway and insulin signaling cascade was not due to changes in the IR mass. Each blot is a representation for one individual experiment.
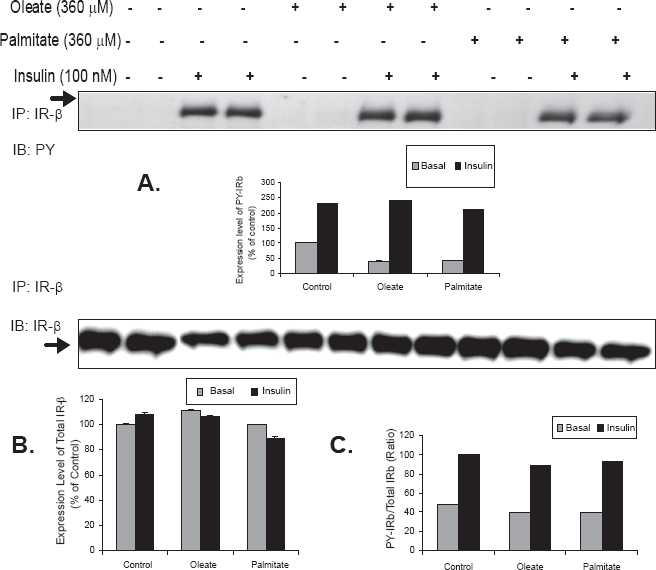
Effect of oleate or palmitate on IR-β subunit tyrosine phosphorylation in primary hamster hepatocytes from fructose-fed hamsters–-Primary hamster hepatocytes from fructose-fed hamsters were treated for 10 min/hours with insulin and 360 μM FFA for 16hours. (A) Cells were lysed and immunoprecipitated using anti-IRβ mouse monoclonal antibody IgG followed by immunoblot with antiphosphotyrosine PY (99) anti mouse monoclonal antibody. (B) Membranes were reprobed using anti IRβ to verify mass T-test was conducted to compare FFAs treated versus appropriate basal or insulin stimulated, significance difference from STD (p < 0.05), (n = 3 experiments/group).
Discussion
The majority of studies have focused on the acute effects of insulin, while the role of chronic hyperinsulinemia and free fatty acids in insulin resistance has been understudied. We employed a fructose-fed hamster as a model of insulin resistance to investigate the mechanisms involved in the state of insulin resistance. This model offers advantages over the more commonly used fructose-fed rat model, in that the metabolism of its apo-B-containing lipoproteins is more similar to that of humans. The rationale for the current study is testing the hypothesis that fructose-feeding results in FFA flux to the liver. Fatty acid estrification pathway rather than fatty oxidation could be implicated in hepatic insulin resistance. This will lead to accumulation of Diacylglycerol (DAG), Triacyl glycerol (TAG) and ceramides. As a result, FFA flux may desensitize hepatic cells to insulin action and reduce tyrosine phosphorylation of IR-β and other downstream signaling molecules involved in PI3K pathway.
In the present study, we intended to investigate, whether chronic treatment of hamster hepatocytes with FFA induces changes in insulin signaling and leads to the development of insulin resistance. It has been shown that liver is characterized as being one of the most insulin sensitive tissues in addition to muscle and adipose tissues. FFA-induced impairment in insulin clearance has been demonstrated in previous
Surprisingly, our data have shown insignificant effect seen for oleate and palmitate on total IR-β receptor mass as well as the IR-β tyrosine phosphorylation in comparison with controls upon stimulation with insulin. The results were confirmed by using a dose response (250 μM-2000 μM) that revealed similar results using either oleate or palmitate (data not shown). Down stream of the IR-β, we also investigated the effect of both oleate and palmitate on the phosphorylation status of PKB. There was no significant effect seen for oleate and palmitate on total PKB mass as well as the serine 473 PKB phosphorylation status that showed a similar results in comparison to their controls upon stimulation with insulin.
This led us to ask the second question: does chronic treatment of hamster hepatocytes with high insulin and FFA desensitize the PI3K pathway. Our data demonstrated that there was insignificant effect seen for oleate and palmitate on total IR-β receptor mass as well as the IR-β tyrosine phosphorylation that showed a similar result in comparison with their controls upon stimulation with insulin. Down stream of the IR-β, we also investigated the effect of both oleate and palmitate on the phosphorylation status of PKB. There was no significant effect seen for oleate and palmitate at on total PKB mass as well as the serine 473 PKB phosphorylation status that showed similar results in comparison with controls observed upon stimulation with insulin. Interestingly, primary hepatocytes extracted from fructose fed hamster have demonstrated a similar effect to the FFA treatment and showed insignificant effect response to the insulin stimulation.
Studies have demonstrated that the number of insulin receptors in liver cells is reduced in glucose-intolerant obese subjects. 35 High FFA levels in the portal blood are correlated with a decreased clearance of insulin in rat liver. 36 Previously, the effect of FFAs on insulin binding, degradation and action was examined in isolated rat hepatocytes. The effect of 0.4 mM oleic acid on insulin binding was reduced cell associated insulin binding by 45%. In agreement to our data, it was surprising to find, FFA does not affect the receptor affinity (comparing cells exposed to oleic acid and their controls). Lonnroth P et al demonstrated that partially purified insulin receptors from solubilized hepatocytes preincubated in the absence or presence of 0.2 mM oleic acid for 45 min with no difference in the binding. 37 It has been noted previously that the technique used for the hepatocytes preparation seems to be crucial for the kinetics of insulin binding and the insulin effect. It has also been demonstrated that the increase of oleic acid levels effectively reduced basal amino acid uptake and the maximal insulin effect, indicating the existence of post receptor perturbations. 38
Independent of the PI3K pathway, other mechanisms might be involved in the development of insulin resistance upon treatment with FFAs. Protein kinase C (PKC), a family of protein serine/threonine kinases has been implicated in FFA-induced insulin resistance.
14
It was postulated that free fatty acids activate classical PKCs isozymes in pretreated muscle cells that lead to insulin resistance development.14,39 Similar
The current findings substantiate our earlier published data which revealed that treatment of cells with FFAs–-induced hepatic insulin resistance through the PKC-NFκB activation pathway. Earlier we have reported that the phosphosite screen using the proteomics approach clearly showed a significant difference in the expression level of the key signaling components involved in the pathway. It manifested a significant increase in the phosphorylation of AMP-activated protein kinase alpha (α), I-kappa-B kinase beta, MKK6, PKC related kinases1 and 2, Raf as well as the mammalian target of rapamycin (mTOR). Consistent with these findings, and in support of our previous published data, a down regulation in the expression level of glycogen synthase kinase-3 alpha/beta (α/β), protein kinase B (PKB) and PKC zeta and lambda (ζ/λ) was observed. The data suggest that fructose feeding develops hepatic insulin resistance through a combination of mechanisms involving the attenuation of the PI3K pathway through FFA effect that occurs down stream of IR-β. 45
Footnotes
Acknowledgements
Authors are grateful to Professor Dr. K. Adeli for his great efforts as a consultant expert in the field of insulin resistance and metabolic dyslipidemia for guiding and revising the manuscript. We also wanted to thank Dr. R.K. Avramoglu, for her sincere feedback and her technical advices during the conductance of the experimental work (The Hospital for Sick Children, Toronto, Canada).
Conflict of Interest
Rafik Ragheb is a recipient of the Hospital for Sick Children Research Training Competition Scholarship. (Restra-comp)–-The University of Toronto, Canada.
Abbreviations
FFA, free fatty acid; Insulin Receptor-beta subunit (IR-β); PKB, protein kinase B.