
Abstract
The myosin superfamily is a versatile group of molecular motors involved in the transport of specific biomolecules, vesicles and organelles in eukaryotic cells. The processivity of myosins along an actin filament and transport of intracellular ‘cargo’ are achieved by generating physical force from chemical energy of ATP followed by appropriate conformational changes. The typical myosin has a head domain, which harbors an ATP binding site, an actin binding site, and a light-chain bound ‘lever arm’, followed often by a coiled coil domain and a cargo binding domain. Evolution of myosins started at the point of evolution of eukaryotes, S. cerevisiae being the simplest one known to contain these molecular motors. The coiled coil domain of the myosin classes II, V and VI in whole genomes of several model organisms display differences in the length and the strength of interactions at the coiled coil interface. Myosin II sequences have long-length coiled coil regions that are predicted to have a highly stable dimeric interface. These are interrupted, however, by regions that are predicted to be unstable, indicating possibilities of alternate conformations, associations to make thick filaments, and interactions with other molecules. Myosin V sequences retain intermittent regions of strong and weak interactions, whereas myosin VI sequences are relatively devoid of strong coiled coil motifs. Structural deviations at coiled coil regions could be important for carrying out normal biological function of these proteins.
Gene Organization and Evolutionary History
Myosin is a family of actin based molecular motors that hydrolyze ATP and generate physical force to move different cargoes inside the cell. This superfamily, divided into at least twenty four classes based on head domain sequence similarity and domain organization1,2 (Fig. 1), drive a large number of biological processes like cytokinesis,3,4 organelle transport,5,6 cell polarization and signal transduction7,8 in eukaryotes. They are typically 1000–2000 residues long and comprise of three domains: a conserved motor head with ATPase activity involved in actin binding and movement; a neck domain with bound regulatory proteins called myosin light chains; and a variable tail that often connects to various cargo associated proteins. The tail domain architectures vary considerably between the subtypes of myosin owing to their functional variation. 9 In conventional myosin (myosin II) the myosin tail region forms a long coiled coil that facilitates dimerization and oligomerization. 10 Coiled coils in many unconventional myosins terminate at a globular cargo binding domain, which associates with specific cargo to be localized in the cell (Fig. 2).

Different classes of myosin superfamily: Phylogenetic tree of 114 myosin sequences showing 24 classes of myosins (Foth et al; 65 Copyright (2006) National Academy of Sciences, USA.).
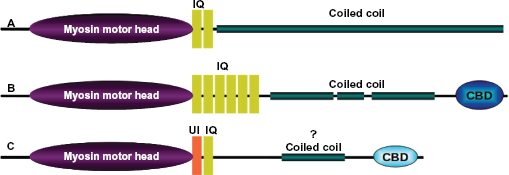
Domain architecture. The overall length and domain organization vary considerably between the subclass members of myosin superfamily. Domain architectures of human (
Myosin genes are highly interrupted with a large number of introns (Fig. 3). The number of exons in mammalian myosin can be as low as 12, as seen in myosins III and VII, whereas the exon number shoots up to 40 and higher in certain subclasses like myosin V. This variation is indicative of many isoforms with different cargo specificity and differences in localization. Most eukaryotes rely on myosins, except a few taxonomic groups, viz. red algae and diplomonad protists
1
which are not known to possess genes with myosin head domain. No classical myosins have been found in prokaryotes. The ATPase domain of myosins along with that of the kinesin family of microtubule based motors is believed to have evolved from the core GTPase domain of G-proteins through deletion of strands 6 and 7 and addition of two N terminal strands.11,12 The wide distributions of myosins I and II across taxonomic levels ranging from yeast to human imply the early evolution of these two classes. Myosins V and XI, which diverged to animal and plant lineages respectively, are thought to have evolved from a common myosin II-like ancestor.
1
The most recent hypothesis put forward the evolution of present day variability among myosins from three ancient myosin subfamilies (see Domain Architecture section for details): (i) the MSD subfamily standing for the general domain architecture of MYSc-SMC-DIL (MYSc-

Gene organization of myosin superfamily. Myosin genes are highly interrupted and contain large number of exons. (
Localization and Function
Myosins are localized to different organs or cell types based on their structural and functional peculiarities. Most of the non-muscle myosins are cytoplasmic. There are a number of myosins suggested to function during transcription within the nucleus by facilitating RNA polymerase movement, like myosin I, 13 myosin II 14 and myosin VI. 15 Possible roles of myosins in transcription and other possible nuclear functions is in need of additional research. Nuclear organization and dynamics of that organization, clearly an exciting and important area of study, is in its infancy. Within the cytoplasm, myosins are localized to various specialized zones, viz. myosin II in the rear of polarized cells, 16 myosin V in growth cones of neuronal cells, 17 myosin VI at the base of stereocilia, and myosin VIII in plasmodesmata of plant cells. 18
Myosin was first described in muscle cells, now known as muscle myosin II. Non-muscle myosin II is found in the cell division furrow of many unicellular and multicellular organisms, 19 where it forms oligomeric thick filaments and facilitates cell division. Non-muscle myosin II isoforms are also localized to the leading edge of growth cones of rat dorsal root ganglion cells (myosin IIB isoform) and in the perinuclear region (myosin-IIA isoform). 20 Myosin II association with Golgi-associated Rab6 resulting in fission of transport carriers was recently described. 21
Myosin V has three isoforms and they are primarily involved in transport of various cellular organelles, such as secretory vesicles (reviewed in Desnos et al., 2007). 22 Myosin V in melanocytes is involved in melanosome transport. 23 In neurons and glial cells, abundant punctate staining of myosin V was found in the peri-nuclear region, along the cellular processes and in the distal tips of processes.24,25 Myosin V is also localized in the centrosome, 26 where it is proposed to facilitate chromosomal segregation. 27 The myosin Va isoform is also involved in acrosome formation and nuclear morphogenesis during spermatogenesis. 28 Myosin VI and VII are present in the inner ear epithelial cells and hair cells. 29 Myosin VI is also localized to the cortical region of the cell, which is the point of endocytosis 30 and to specialized clathrin-coated invaginations at the base of the brush-border microvilli. 31 Myosin VI is involved in two distinct steps of endocytosis in higher eukaryotes: the formation of clathrin-coated vesicles and the movement of nascent uncoated vesicles from the actin-rich cell periphery to the early endosome with the help of various adapter proteins. 32 For a more extensive review of the roles of the myosin family of molecular motors in cells and tissues, see Hartman et al., 2011. 33
General Structural Features
Myosins function as monomers, 34 dimers 35 or oligomers 36 to bring about the diverse sub-cellular movements described above. In general, the myosin protomer consists of a long polypeptide known as the heavy chain comprising an ~80 kDa head domain followed by a variable length tail domain. The head domain is the most conserved and possesses the ATPase activity and actin binding ability.37,38 The C-terminal portion of the head is an α-helix with bound light chains that acts as a lever arm that amplifies smaller movements within the catalytic domain during the chemo-mechanical ATPase cycle. This light chain bound α-helix consists of a varying number of IQ motifs ([I,L,V]QxxxRGxxx[R,K]) where the light chains, calmodulin or calmodulin-like proteins, bind. A typical dimeric myosin consists of two heavy chains with a varying number of light chains, depending on the myosin type. In type II myosins, an essential light chain (ELC) and a regulatory light chain (RLC) bind at the IQ motifs. 36
The C-terminal tail of unconventional myosins often, but not always, have a coiled coil region and a terminal globular cargo-binding domain. Conventional myosins (myosins II) possess long coiled coil tails and lack the C-terminal globular domain typical of the unconventional myosins.
Specific Domain Architectures
To date, the most extensively studied myosin head is that of myosin II, with numerous crystal structures, the first one being the motor domain of chicken skeletal muscle myosin. 39 In the recent past, the myosin V motor domain was crystallized, 40 as well as that of myosin VI. 41 In all cases, the myosin head has an upper 50 kDa domain and a lower 50 kDa domain, connected by a loop. It has a converter domain, and a P-loop, switch I and switch II near the active site. Myosin II and a few other myosin classes possess an N-terminal SH3 domain. In myosin VI, there is a unique insert just after the converter domain that enables this motor to reverse its direction of movement toward the minus end of an actin filament. In addition, there are a number of domains, such as the PH domain, SH3 domain, and Kinase domain, which when combined with the myosin head make a plethora of domain architectures.
At the C-terminus of many non-muscle myosins is a globular cargo binding domain (CBD) that provides specificity for particular myosins binding to particular cargo. The CBD of myosins are relatively poorly conserved within the superfamily. Structures of only two CBDs, belonging to myosin V and VI, are known (Fig. 4). Myo2p is an essential myosin V in Saccharomyces cerevisiae involved in multiple functions through binding to multiple cargoes. Cargo binding domain of yeast M yosin V is an all-alpha fold containing 15 amphipathic α-helices connected by short and long loops organized into two five-helical bundles. These helical bundles correspond to sub-domains I and II originally defined by mild proteolysis and corresponds to two binding sites (one for vacuole binding and the other for secretory vesicle binding) that are oppositely placed at a 180° angle. 42 The myosin V CBD is also known as the DIL domain. The myosin VI CBD forms a new fold that contains four β strands (βA–βD) and six α helices (αA–αF). 43 The four β strands form an antiparallel β sheet, creating the core of the domain. One side of the β sheet is covered by three helices (αD–αF) that are oriented perpendicular to each other. The other side of the sheet is largely exposed to the solvent with a short αC helix capped at one of its edges. The mouse myosin VI CBD binds to a fragment of the clathrin-coated vesicle adaptor Dab2 with high affinity. 44
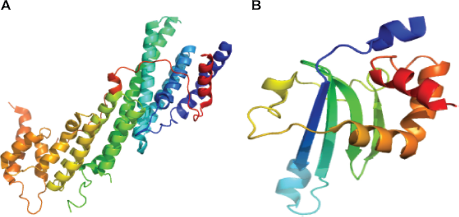
Crystal structures of Cargo Binding Domain (CBD). (
Between the head domain and the C-terminus of many myosins is a coiled coil structural motif that facilitates dimerization or oligomerization. The first myosin coiled coil crystal structure was from scallop myosin. 45 The length of the coiled coil region in myosin varies from class to class. Myosins II have a very long heptad repeat, whereas myosin VI has essentially no predicted coiled coil regions (see below) and myosin V possesses a coiled coil of intermediate length (Fig. 2).
Coiled Coil Interaction Strength
In spite of the prominence of the coiled coil as an important motif in the myosin superfamily, structural and functional details of these coiled coils are severely lacking, due to the general absence of crystal structures. Though most of the myosin coiled coils have typical heptad repeats, the strength of the interactions between the two α-helices forming the various segments of the coiled coil are likely to vary. Very recently, the participation of the coiled coil motif in thick filament assembly of yeast myosin II has been suggested to involve two pathways. These investigators have employed deletion mutants and in vivo assays to demonstrate the role of the coiled coil, and an evolutionarily conserved structural kink within, in forming cleavage furrow ingression at the division during cytokinesis. 46 In the literature, demarcation of coiled coil regions into weak and strong regions, protein-protein interaction sites and the recognition of structural kinks are just emerging.
There are many prediction programs that are available to predict the rough positions of coiled coils given the amino acid sequence of a protein. 47 There are also structural analysis programs that enable identification of the coiled coil boundaries following the ridges-grooves arrangement of amino acid side chains. 48 However, most of the predictive programs assume a uniform and ideal strength to the predicted regions, albeit being sensitive to sequence breaks like stutters and stammers. The sequence signatures at the coiled coils have evolved to such an extent that one can hardly find any conserved motifs across the subtypes. Yet many conserved (at the amino acid level) motifs can be found in the predicted coiled coil tail (please see a multiple sequence alignment of myosin VI in Supplementary Fig. 1).
We have calculated the strength of interactions between the coiled coil α-helices within dimeric myosins through the pseudo-energy function inscribed in COILCHECK. 49 For this we applied an algorithm (Fig. 5) that computationally ana lyzes the interaction strength of myosin at the coiled coil motif by three-dimensional modeling using MODELLER 50 followed by retrospective calculations of interaction energies at the coiled coil interface using COILCHECK. Unlike general protein-protein interfaces51,52 that are largely dominated by hydrophobic interactions, the coiled coil interfaces are special, elaborate and retain alternating hydrophobic interactions and salt bridges built around a simple system of a pair of helices. Van der Waals and electrostatic interactions are measured all along the coiled coils across the dimer (see Fig. 5 for Methods). Further, interactions such as hydrogen bonds and salt bridges also contribute to the stability of these domains. This method has been applied to 26 protein structural entries that contain coiled coil. The binding strength values correlate well with the structure and stability of coiled coils. Indeed, COILCHECK results are sensitive to demarcate DNA-binding proteins that structurally deviate from ideal coiled coils (Fig. 6A). The guanine nucleotide exchange factor (GEF) domain of sec2p protein is a 22-nm long coiled coil and has a weakly interacting N-terminal region and a strongly intertwined middle region as evident from the crystal structure. 53 Calculation of the interaction energies at different regions of the GEF domain using COILCHECK are consistent with the experimental results (Fig. 6B).
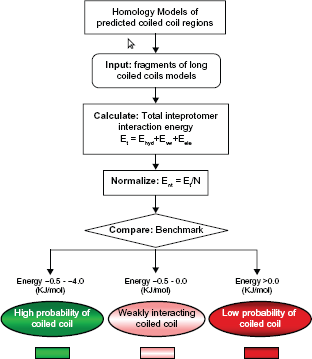
Coiled coils interaction strength analysis protocol. Molecular models of myosin coiled coils were made using tropomyosin as template. The models were generated using MODELLER program. 50 The long models were split into penta-heptad sized fragments and interaction energy values were calculated using COILCHECK program.

COILCHECK based inter-protomer interactions. (
When COILCHECK was applied to the tropomyosin structure (PDB code: 1C1G), actin-interacting zones were observed to acquire relatively poor energies for coiled coil formation (Fig. 6C). Furthermore, in order to probe the sensitivity of COILCHECK in detecting local structural perturbation due to the introduction of mis-sense mutations, we performed a series of virtual mutations (Fig. 6D). Interestingly, COILCHECK energy differences between the wild type and ‘single-site mutants’ were not very high when permitted amino acid exchanges, such as Leu to Val, were performed at either ‘a’ or ‘d’ positions. On the other hand, if drastic amino acid changes, such as Leu to Asp or Val to Lys, were introduced in the coiled coil, COILCHECK energy differences were quite high (Fig. 6D), suggesting that the method is sensitive to predict the effect of small sequence changes on the stability at the interface.
Using this approach, we found that the coiled coil regions of myosins V are generally interrupted by poorly interacting dimeric zones (Fig. 7B). Myosins VI have only a couple of short predicted strong interactions in its α-helical tail (Fig. 7C and D). These myosin VI results are consistent with experimental findings that the putative coiled coil region (ie region predicted as coiled coil by COILS or PAIRCOIL algorithms) of myosin VI, are actually not coiled coil but stable single α-helices. 54 Coiled coils of myosins II are predicted to have many segments of weak interactions that may have roles in oligomerization or protein-protein interactions (Fig. 7A).

Inter-protomer interaction strength. Non-uniform inter-protomer interaction strength in predicted coiled coil regions are color coded and represented as boxes. Each box corresponds to 35 amino acids. Strongly interacting regions are shown in green and non interacting regions are shown in red. An intermediate level of interaction is also possible and shown in lighter shade of red. (
Molecular Mechanisms
The myosin motor proteins convert chemical energy stored in ATP into mechanical movement by way of conformational changes within the head that are amplified by the neck domains. Myosins work by a swinging cross-bridge mechanism, 55 the cross-bridge being the head domain. 56 Following the first crystal structure determination of the myosin II head domain (see Rayment et al, 1993), 39 the swinging cross-bridge hypothesis was refined to the swinging lever arm hypothesis. In the swinging lever arm hypothesis, the light chain binding region acts as a lever arm and generates at least three conformational isomers of the myosin during the ATPase cycle.
Myosin II has a step size of 5–15 nm, as evident from single molecule measurements. 57 The ATPase site of the head domain has a nucleotide-binding site, which is a phosphate-binding loop (P-loop) that is closely associated with switch-I and switch-II helices. The actin-binding site and the P-loop communicate the presence or absence of Pi through switch-I and switch-II helix movements.
Myosin V and VI follow variants of the swinging lever arm mechanism, and achieve distinct step sizes, processivity and directionality.58,59 The unique long neck of myosin V with six calmodulin binding sites (IQ motifs) contribute to the long step size of 36 nm. Electron micrographs of myosin VI bound to actin revealed that the light chain binding region is oriented toward the pointed end of the actin unlike other myosins.41,60 Albeit having a short neck similar to myosin II, myosin VI step sizes are comparable to that of Myosin V. This large step size of ~36 nm owes to a mechanism of a 180° twist of its lever arm59,61 and an unusual tail domain that allows the 36-nm stretch.62,63 The reverse directionality of myosin VI comes from a unique insert just following the converter region.41,61 Diverse domain combinations of the myosin superfamily members result in imparting a wide variety of motility associated functions in the cell. The general myosin head-coiled coil-cargo binding domain architecture of many unconventional myosins is designed in such a way that vesicles or organelles are attached at the cargo binding domain of the dimer and the ATPase hydrolysis results in the walking motion of the dimer along the actin filament. Myosin VI is the only known retrograde myosin, which walks toward the minus end of the actin filament. Myosin VI dimerization appears to occur upon cargo binding. 64
Conclusion
On the basis of available literature and our own analysis, we have narrated the molecular features of the myosin superfamily. The subfamily types described here are derived based on sequence similarity relationships of the motor domain. Further investigations on sequence, structural and functional diversity of the tail domain will provide the basis for a deeper understanding about this superfamily and new classification schemes.
We wish to draw attention to the putative coiled coil regions of the myosins because these are relatively understudied. Indeed, as shown for myosin VI, the strongly predicted coiled coil region appears not to be a coiled coil at all. Interestingly, COILCHECK, unlike COILS or PAIRCOILS, suggests very weak coiled coil interactions in myosin VI. COILCHECK is now being applied to all the myosin coiled coils to better understand which regions are likely to be strong interacting domains and which are weak. These data together with experimental data on the functions of these different domains are likely to shed new light on the roles that these myosin tail domains play in myosin structure and function.
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.
Footnotes
Acknowledgements
DPS's stay at NCBS is supported by NCBS and Indian Council of Agricultural Research. This work is supported by Human Frontier Science Program. We also thank NCBS (TIFR) for infrastructural support.