
Abstract
Interphase nuclei are organized into structural and functional domains. The coiled body, a nuclear organelle of unknown function, exhibits cell type-specific changes in number and morphology. Its association with nucleoli and with small nuclear ribonucleo-proteins (snRNPs) indicates that it functions in RNA processing. In cycling cells, coiled bodies are round structures not associated with nucleoli. In contrast, in neurons, they frequently present as nucleolar “caps.” To test the hypothesis that neuronal differentiation is accompanied by changes in the spatial association of coiled bodies with nucleoli and in their morphology, PC12 cells were differentiated into a neuronal phenotype with nerve growth factor (NGF) and coiled bodies detected by immunocytochemical localization of p80-coilin and snRNPs. The fraction of cells that showed coiled bodies as nucleolar caps increased from 1.6 ± 0.9% (mean ± SEM) in controls to 16.5 ± 1.6% in NGF-differentiated cultures. The fraction of cells with ring-like coiled bodies increased from 17.2 ± 5.0% in controls to 57.8 ± 4.4% in differentiated cells. This was accompanied by a decrease, from 81.2 ± 5.7% to 25.7 ± 3.1%, in the fraction of cells with small, round coiled bodies. SnRNPs remained associated with typical coiled bodies and with ring-like coiled bodies during NGF-induced recruitment of snRNPs to the nuclear periphery. Together with the observation that coiled bodies are also present as nucleolar caps in sensory neurons, the results indicate that coiled bodies alter their morphology and increase their association with nucleoli during NGF-induced neuronal differentiation.
Keywords
T
Components of the interchromatin space are considered to contribute to this substructure, and include perichromatin fibrils, interchromatin granule clusters, and nuclear bodies. Among these, nuclear bodies have received considerable attention as dynamic structures (Brasch and Ochs 1992), exhibiting cell cycle- and cell state-dependent changes in number and in morphological complexity (Brasch et al. 1989; Andrade et al. 1991,1993; Malatesta et al. 1994; Ochs et al. 1995; Berciano et al. 1996; Rebelo et al. 1996). One such nuclear body in which such changes have been well documented is the coiled body (CB) (Andrade et al. 1993; Ochs et al. 1995; Rebelo et al. 1996), a conserved nuclear structure that occurs in both plant and animal cells. Although originally described as an “accessory body” because of its frequent association with the nucleolus in neurons (Ramon y Cajal 1903), its current designation as a coiled body is based on ultrastuctural studies in which it presents as a noncapsular, round to oval structure composed of meandering threads loosely arranged in a coiled fashion (Hardin et al. 1969; Monneron and Bernhard 1969; Malatesta et al. 1994).
Much of the recent interest in CBs has resulted from the identification of human autoimmune antibodies that recognize an 80-kD CB-specific protein named p80-coilin (Andrade et al. 1991; Raska et al. 1991). Studies by Andrade and co-workers (1991) have shown that CBs are enriched in p80-coilin, a protein that can be used as a specific marker for CBs. In addition to p80-coilin, several other proteins have been reported to be associated with CBs. Specifically, CBs have been reported to contain the snRNP-associated Sm proteins U1, U2, U4, and U6 (Fakan et al. 1984; Raska et al. 1991) and DNA topoisomerase (Raska et al. 1991). Although CBs also contain nucleolar proteins including proteins associated with nucleolus organizer regions (NORs) (Raska et al. 1990), Nopp140 (Meier and Blobel 1992), and fibrillarin (Raska et al. 1990; Carmo–Fonseca et al. 1993), they differ from nucleoli by the absence of the nucleolar proteins B23 and nucleolin (Raska et al. 1991).
The identification of a subset of nucleolar antigens in CBs suggests the existence of a structural similarity between CBs and nucleoli. In fact, early electron microscopic studies in neurons revealed that CBs occur in close association with the nucleolus, sometimes appearing as a budding structure (Hardin et al. 1969; Lafarga et al. 1983). Immunocytochemical studies have also shown that anti-p80-coilin antibodies labeled perinucleolar “caps” in nuclei of rat hippocampal (Carmo–Fonseca et al. 1993) and cerebellar (Raska et al. 1990) neurons. A similar pattern of capping of nucleoli with CBs was also observed in primary human fibroblasts, when grown at 29C, and in HeLa cells exposed to transcription inhibitors (Carmo–Fonseca et al. 1993). There exists, in addition, a considerable similarity between CBs and the dense fibrillar component of the nucleolus (Lafarga et al. 1983,1991; Thiry 1995). Both exhibit equal and specific argentophilic properties (Seite et al. 1982; Lafarga et al. 1983; Berciano et al. 1996). Further evidence comes from Bohmann and co-workers (1995), who reported that disorganization of both CBs and nucleoli may result from the overexpression of certain mutated forms of the p80-coilin gene. Together, these findings suggest a common structural framework or assembly pathway for CBs and nucleoli.
On the basis of their association with the nucleolus, CBs were believed to have a direct role in the processing of rRNA (Hardin et al. 1969; Raska et al. 1990; Lafarga et al. 1991). In fact, the presence of several Sm antigens in CBs (Raska et al. 1991), and of specific U-snRNAs (Thiry 1995), points to a potential role for CBs in pre-mRNA metabolism or in snRNP assembly or disassembly.
In actively cycling cells, CBs have been most often described as nucleoplasmic, uniformly staining round bodies, enriched in snRNPs and other splicing components (Andrade et al. 1993; Gall et al. 1995; Berciano et al. 1996). In contrast, in nuclei of neurons, a cell type considered to be terminally differentiated, CBs have been shown to occur in close association with nucleoli, appearing as nucleolar caps enriched in p80-coilin and snRNPs (Raska et al. 1990; Carmo–Fon-seca et al. 1993). In the work reported here, rat pheochromocytoma (PC12) cells were used as a model for neuronal differentiation. These cells undergo differentiation from actively cycling, chromaffin-like cells into sympathetic-like cholinergic neurons on treatment with nerve growth factor (NGF) (Greene and Tischler 1976). Such neuronal differentiation of PC12 cells has been previously shown to be associated with a redistribution of snRNPs and of DNase I-hypersensitive chromatin from a speckled distribution to a distibution characterized by a peripheral shell closely apposed to the inner nuclear envelope (Sahlas et al. 1993; Park and De Boni 1996). In the work reported here, PC12 cells were employed in a test of the hypothesis that neuronal differentiation is accompanied by an increase in the spatial association between CBs and nucleoli and by changes in the morphology of CBs. To test this hypothesis and to investigate whether snRNPs remain associated with CBs during redistribution of snRNPs to the nuclear periphery, CBs were double labeled by immunocytochemistry for p80-coilin and Sm antigens, and their intranuclear spatial distributions and numbers were analyzed by confocal laser scanning microscopy.
Materials and Methods
Cell Culture
Rat pheochromocytoma cells (Greene and Tischler 1976) were propagated at 37C in a humidified environment in tissue culture flasks (Becton Dickinson; Mountain View, CA). Culture medium consisted of 85% (v/v) minimal essential medium (MEM) with Hanks' salts and
Dorsal root ganglia (DRG) were excised aseptically from newborn CD-1 mice and suspended in a drop of cold Hanks' balanced salt solution (HBSS; GIBCO). Ganglia were sedimented (500 x g, 2 minutes) and the pellet incubated in trypsin (0.25%; 2 ml, 37C, 11 min; GIBCO) to allow dissociation of cells. Upon inactivation of trypsin with FBS (1 ml), the ganglia were mechanically disrupted by trituration with a Pasteur pipette and the dissociated cells sedimented (500 x g, 5 min). The pellet was resuspended in 0.2 ml of pre-warmed (37C) DRG culture medium (90% MEM with Hanks' salts and
Immunocytochemistry
Cultures were fixed (12 min) with formaldehyde freshly prepared from paraformaldehyde (4% w/v) in PBS (130 mM NaCl, 7 mM Na2PO4, 3 mM NaH2PO4, pH 7.2), washed in PBS (three times for 5 min), and permeabilized [Triton X-100, 0.1% (v/v), PBS, 30 min]. To reduce nonspecific binding of antibodies, cultures were blocked with bovine serum albumin (BSA) [4% (w/v), PBS, 30 min].
CBs were labeled using rabbit R288 antibody to p80-collin (Andrade et al. 1993) (1:150, 4% BSA, 2 hr; provided by E.K.L. Chan) and detected with FITC-conjugated goat anti-rabbit antibody (1:100, 4% BSA, 1 hr; Jackson ImmunoResearch, Hornby, Ontario, Canada).
snRNPs were labeled using human anti-Sm autoimmune serum (1:200, 4% BSA, 2 hr; ANA Human Reference Serum no. 5, Centers for Disease Control, Atlanta, GA) and detected with TRITC-conjugated goat anti-human IgG (H + L, 1:100, 4%BSA, 1 hr; Jackson ImmunoResearch). For double labeling of p80-coilin and snRNPs, sequential incubation in both anti-Sm and anti-p80-coilin with an intervening wash was carried out before sequential incubation with the appropriate secondary antibodies. For visualization of nuclear and nucleolar outlines, when required, cells were counterstained with ethidium bromide (1 μg/ml, PBS, 10 min; Sigma).
After labeling, coverslips were inverted on glass slides into a drop of freshly prepared p-phenylenediamine solution (1 mg/ml, 50% glycerol, PBS) to reduce photobleaching (Johnson and Nogueiro–Aguaro 1981). Cells were visualized with a Zeiss confocal laser scanning (LSM 410) microscope equipped with an argon/krypton laser. FITC and TRITC fluorochromes were excited using 488-nm and 568-nm wavelength light, respectively, passing through a x63/1.4 NA oil immersion lens. Digital images were stored on disk and representative images printed with a Kodak 8650 PS color printer.
For analysis, cells exposed to NGF for 10–14 days were categorized as neuronal or non-neuronal on the basis of cell morphology. Specifically, only those cells that exhibited large (diameter <20 μm) rounded cell bodies and extensive neurite outgrowth (minimal length <2 x cell body diameter) were designated as neuronal (Figure 1). Changes in the morphology and in the number of CBs per cell nucleus were compared among controls, NGF-treated cells of neuronal morphology, and NGF treated cells that failed to exhibit morphological evidence of differentiation. For quantification of the number of CBs per nucleus, nuclei were optically sectioned at 0.4-μm steps.
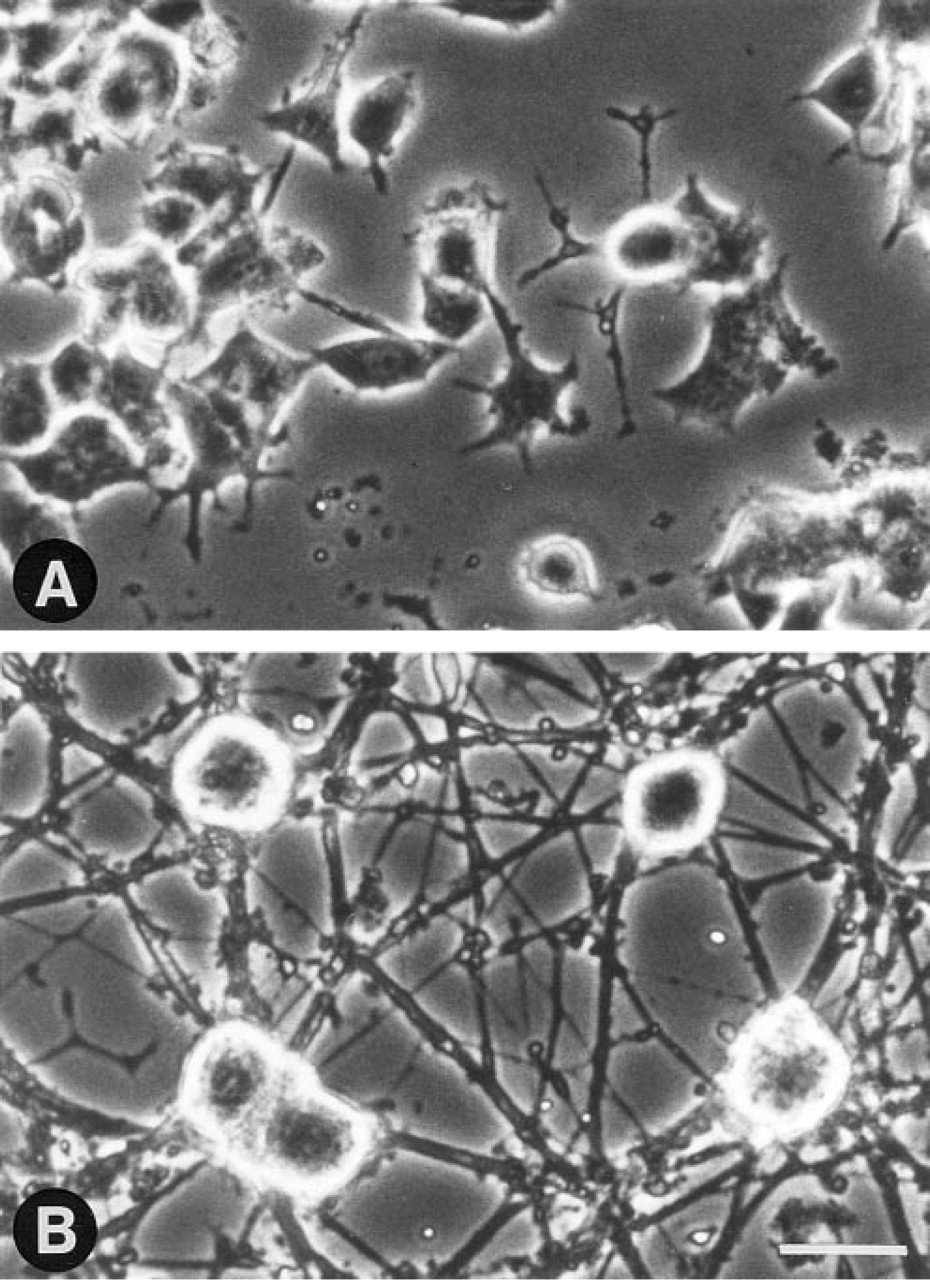
NGF-induced neuronal differentiation of PC12 cells. Phase-contrast micrographs of (
Results
NGF-induced Neuronal Differentiation of PC12 Cells Is Accompanied by a Change in CB Morphology Without a Change in CB Number
In PC12 cells, CBs, detected by immunocytochemistry of associated p80-coilin, varied in morphology and were classified into three categories. As described in previous studies (Andrade et al. 1993; Ochs et al. 1995), CBs appeared as small (0.5–1.0-μm diameter), round, uniformly staining structures within the nucleoplasm, occurring without a distinct association with the nucleolus (Figures 2A and 2A″). CBs with this morphology are herein referred to as typical coiled bodies (TCBs). In addition to this typical morphology, CBs also occurred as larger (1.5–2.5-μm wide) structures, exhibiting a ring of p80-coilin, herein referred to as ring-like CBs (RLCBs) (Figures 2B and 2B″). The absence of ethidium bromide-positive ligands in these structures indicates that RLCBs are unlikely to represent additional small nucleoli (Figure 3). Such structures exhibiting a peripheral shell of p80-coilin (Figures 2 and 3) had previously been described (Raska et al. 1990). In addition, and most commonly observed in NGF-responsive PC12 cells with a neuronal morphology, CBs also appeared as a hemisphere apposed to the nucleolus, with a peripheral distribution of p80-coilin (Figures 2C and 2C″). This subtype has been referred to as a nucleolar cap (Carmo–Fonseca et al. 1993).
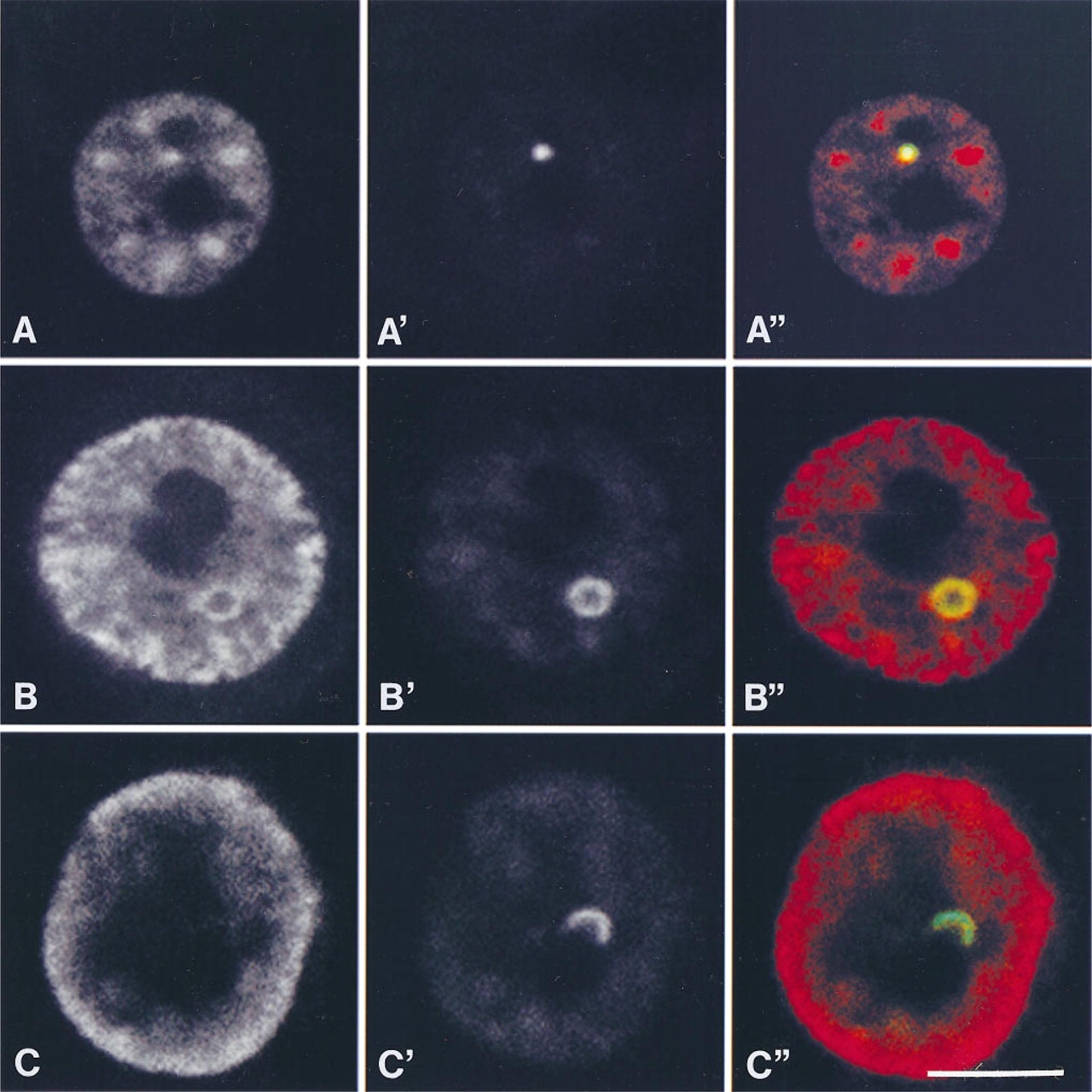
Laser confocal micrographs showing co-localization of snRNPs and p80-coilin in three characteristic subtypes of CBs in nuclei of PC12 cells. (
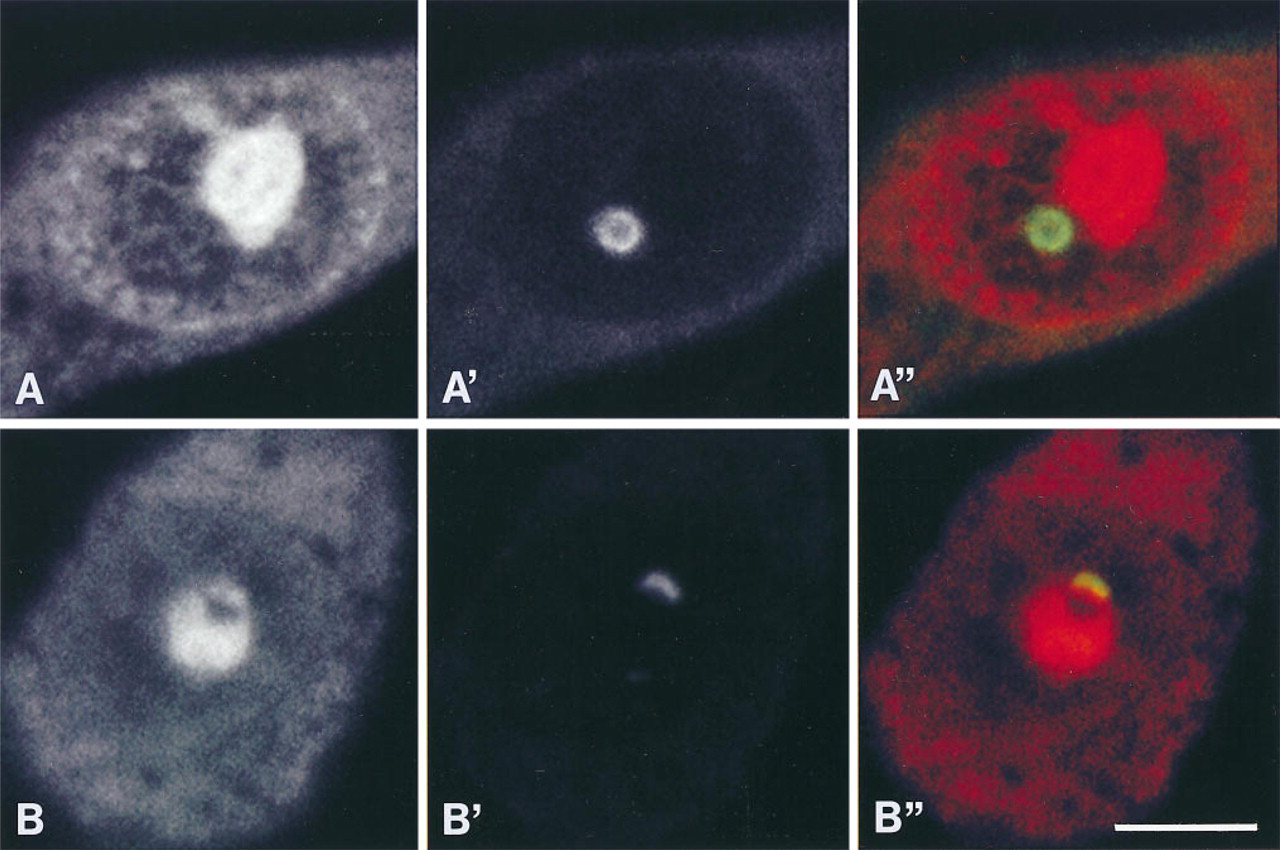
Representative laser confocal micrographs of NGF-responsive PC12 cells with RLCB (
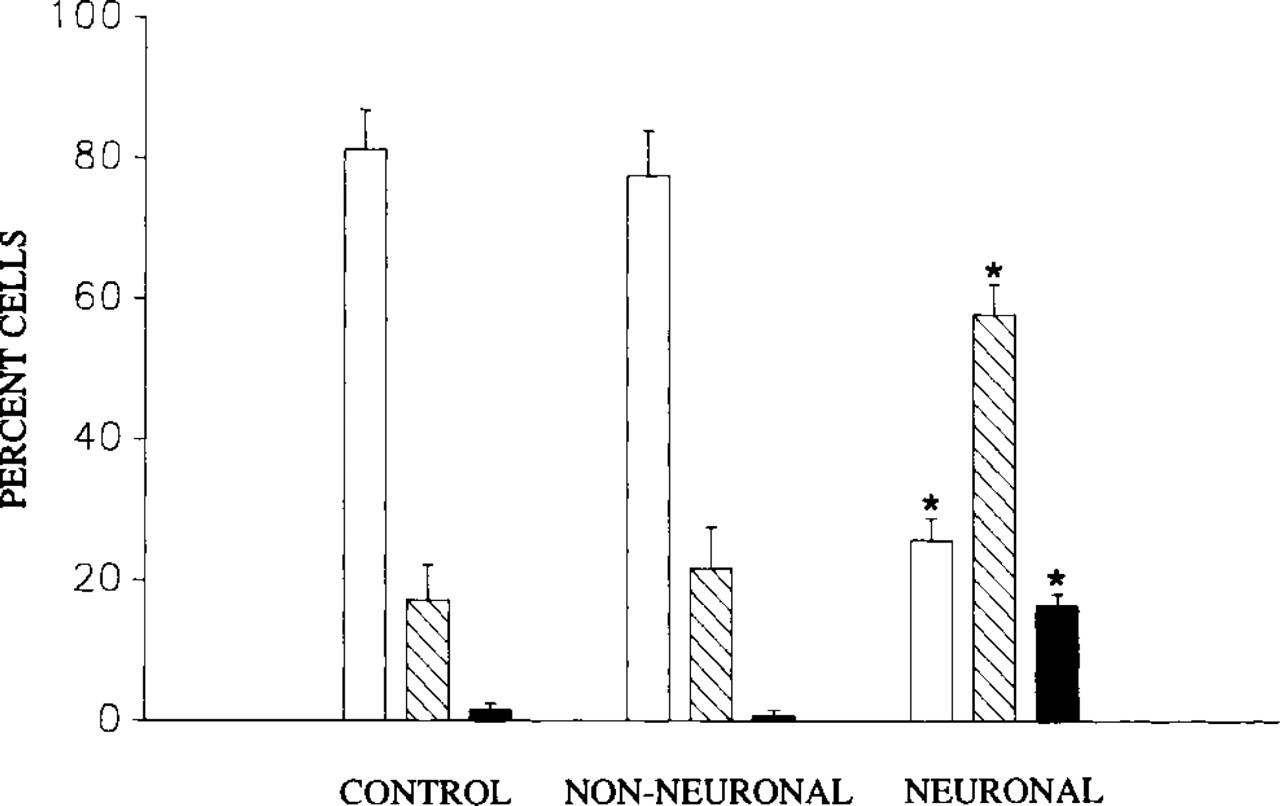
Bar graph showing association of NGF-induced neuronal morphology with significant (asterisks, ANOVA, p=0.05, multiple comparisons) changes in the fraction (% ± SEM) of PC12 cells that display TCBs (open bars), RLCBs (diagonal bars) and nucleolar caps (solid bars). Note decrease in fraction of cells with TCBs and increase in fraction of cells with RLCBs and nucleolar caps in cells with neuronal morphology compared to both controls and non-NGF-responsive “non-neuronal cells.” (Controls, n = 400 cells, six cultures. Non-neuronal, n = 300 cells, six cultures. Neuronal, n = 300 cells, six cultures.) Cells in NGF-treated cultures are divided into non-neuronal (non-NGF-responsive) and neuronal (NGF-responsive) categories by morphological criteria (see text).
Although all three of these CB subtypes were observed in both untreated and treated PC12 cells, the frequency with which they occurred differed between cells with neuronal and non-neuronal morphologies (Figure 4). Specifically, in PC12 cells that were not exposed to NGF, CBs most commonly appeared as nucleoplasmic TCBs and less commonly as RLCBs. Only in a small fraction of untreated cells were CBs observed in association with the nucleolus, as nucleolar caps. In cells treated with NGF but that failed to exhibit a neuronal morphology, the frequencies with which these CB subtypes occurred did not differ from those observed in untreated cells. In contrast, however, in cells that responded to NGF by differentiation into cells of neuronal morphology, the fraction of cells with TCBs was significantly lower than that observed in non-neuronal control cells and in non-neuronal NGF-treated cells within the same culture. This decrease was paralleled by a significant increase in the fraction of cells that exhibited RLCBs and nucleolar caps, compared to untreated and NGF-nonresponsive cells (Figure 4).
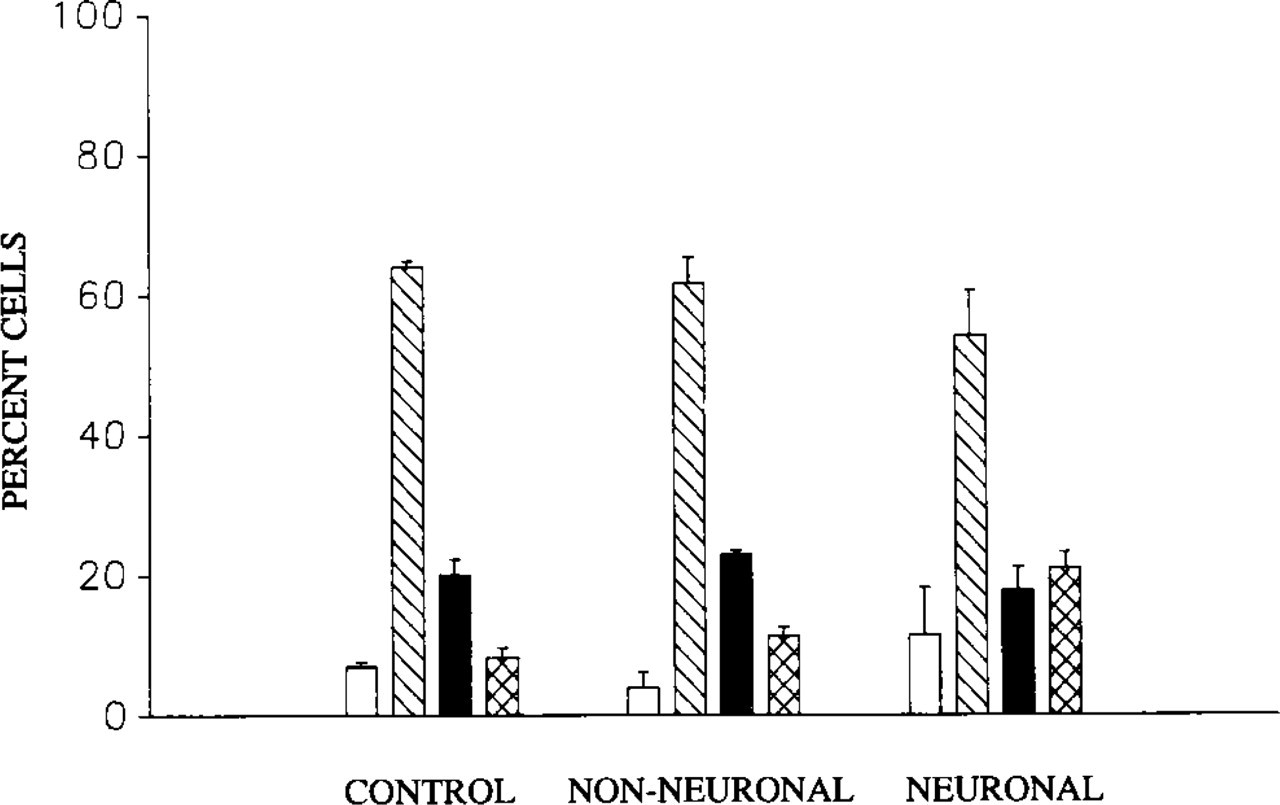
Bar graph showing that neuronal differentiation of PC12 cells is not associated with changes in the number of CBs per nucleus (ANOVA, p=0.05). Shown are fractions (% ± SEM) of cells with no CBs (open bars), 1 CB (diagonal bars), 2 CBs (solid bars), and 3 or more Cbs (range 3–7) (crosshatched bars). (Controls, n = 600 cells, seven cultures. Non-neuronal, n = 300 cells, five cultures. Neuronal, n = 500 cells, eight cultures.) Note lack of changes in number of CBs. See text for criteria for non-neuronal vs neuronal categories.
To determine whether increases in the number of cells exhibiting RLCBs and nucleolar caps were attributable to the formation of new coiled bodies, the number of CBs per nucleus were quantified in untreated and NGF-treated PC12 cells. In cells exposed to NGF, the mean number of CBs per nucleus in both those cells responding to NGF with a neuronal morphology and in NGF-nonresponsive cells did not differ from that observed in untreated (control) cultures (ANOVA; p=0.05). In addition, the fraction of cells that exhibited either none, one, two, or multiple (range 3–7) CBs did not differ (ANOVA; p=0.05) between controls, NGF-responsive, and NGF-unresponsive cells. In all these cases, nuclei most commonly contained one CB (Figure 5).
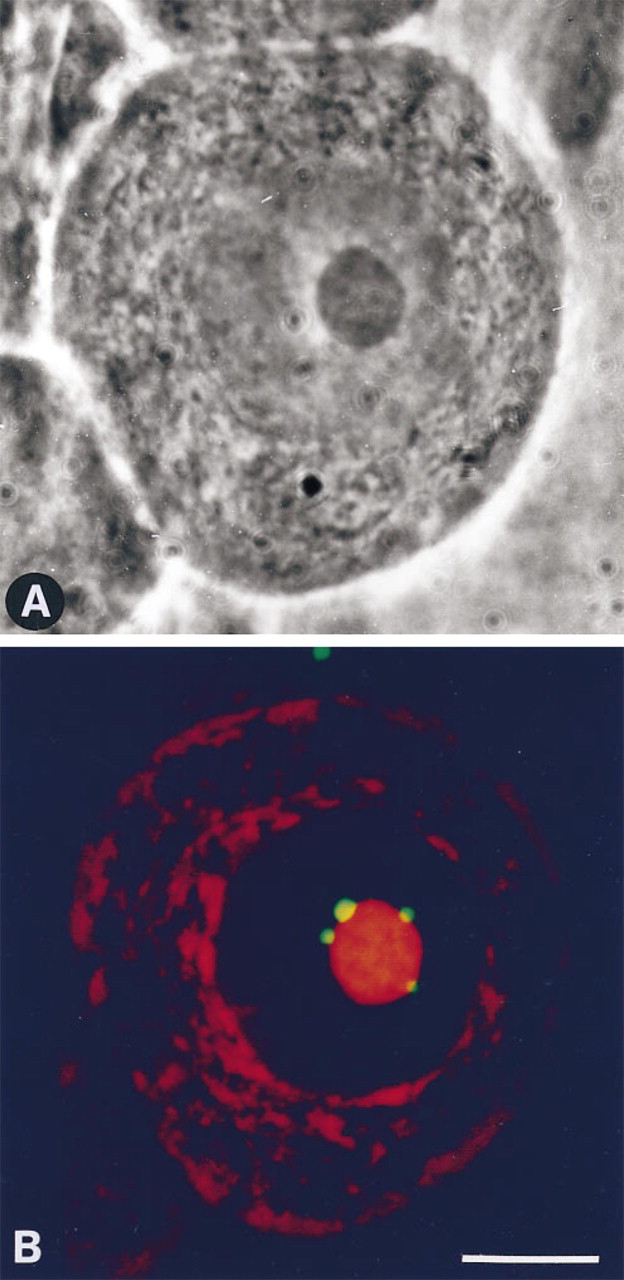
Phase-contrast (
CBs Remain Enriched in snRNPs During NGF-induced Redistribution of Sm Antigens to the Nuclear Periphery
In PC12 cells, snRNPs are recruited to the nuclear periphery in response to NGF (Sahlas et al. 1993). Given that CBs contain Sm antigens in addition to p80-coilin, the association between these two antigens was monitored by double labeling to test whether their association is altered during NGF-induced recruitment of snRNPs to the nuclear periphery. snRNPs and p80-coilin invariably co-localized in TCBs within nuclei of both control and NGF-treated cells. In RLCBs, snRNPs also co-localized with p80-coilin, with both occurring at the periphery of the CB and absent from the CB center. Furthermore, snRNPs remained associated with RLCBs in nuclei of NGF-exposed cells in which the majority of snRNPs were detected at the nuclear periphery (Figures 2B and 2B″). In contrast, in 12 of 30 cells (40%) examined, nucleolar caps in nuclei of PC12 cells with neuronal morphology were devoid of snRNPs as indicated by their green pseudocolor, indicating lack of a red overlapping signal (Figures 2C and 2C″).
CBs Appear as Paranucleolar Caps in Nuclei of DRG Neurons
To compare the distribution of CBs in PC12 cells that exhibit an NGF-induced neuronal morphology to that typically observed within nuclei of normal neurons, CBs were detected by immunocytochemical labeling of associated p80-coilin in nuclei of murine DRG neurons in vitro. In such neurons (Figure 6), CBs most commonly occurred as nucleolar caps (1–5 per nucleolus), with a spatial distribution similar to that observed in NGF-differentiated PC12 cells as described above and as described previously (Raska et al. 1990; Carmo–Fonseca et al. 1993). It is noteworthy that the nucleolar caps in DRG neurons did not stain with ethidium bromide, an indication that the structures apposed to the nucleolus are not additional nucleoli in the process of nucleolar fusion (Park and De Boni 1991).
Discussion
The results of the present study, which show increases in the frequency of RLCBs and nucleolar caps in nuclei of NGF-responsive PC12 cells of neuronal morphology, indicate that NGF-induced neuronal differentiation is accompanied by, in addition to changes in the morphology of CBs, an increased association of CBs with nucleoli (Figures 2 and 4). It is noteworthy that the changes in frequency of occurrence of RLCBs and nucleolar caps were restricted to those cells that were induced to exhibit a neuronal morphology, and were not detected in NGF-exposed cells within the same cultures that failed to differentiate. The mode of action of NGF that results in the above changes in morphology and spatial position of CBs therefore remains unclear. However, in DRG neurons, CBs similarly occur in association with nucleoli, as nucleolar caps. An association between CBs and nucleoli, therefore, appears to be a feature common to neuronal cells and may represent a nuclear organization associated with neuronal differentiation, as previously reported for hippocampal neurons (Santama et al. 1996).
It was also observed that changes in the frequency of occurrence of the three morphological subtypes of CBs on NGF-induced neuronal differentiation of PC12 cells were not accompanied by a change in the number of CBs per nucleus. This suggests that increases in the occurrence of ring-like CBs and nucleolar caps are not associated with the formation of new CBs, as has been proposed to occur through assembly and disassembly of CBs at the periphery of nucleoli in rat hippocampal neurons (Carmo–Fonseca et al. 1993). Moreover, the finding that NGF-induced neuronal differentiation of PC12 cells was not accompanied by a change in the number of CBs is in contrast to studies that have shown an increase in the number of CBs under conditions that alter gene expression (Brasch and Ochs 1992; Ochs et al. 1995; Berciano et al. 1996). It is also in contrast to the decreased number of CBs described to occur during differentiation of rat cerebellar Purkinje neurons (Lafarga et al. 1995). The observation that NGF alters the morphology and spatial distribution but not the number of CBs may therefore be uniquely related to NGF-induced differentiation of PC12 cells.
It must also be considered that the increase in the incidence of ring-like CBs and nucleolar caps may have resulted from changes in the morphology and distribution of existing CBs, in which ring-like CBs may represent intermediate structures before their association with nucleoli as caps. It is interesting to note that, similar to RLCBs, nucleolar caps also exhibit peripheral p80-coilin with an unlabeled core, as demonstrated in the present study in differentiated PC12 cells. Such peripheral labeling of nucleolar caps was demonstrated in previous studies in which p80-coilin was described as labeling a cap-like patch at the nucleolar periphery (Carmo–Fonseca et al. 1993). In fact, there is a resemblance between the ring-like CB and the hemispheric structure apposed to the nucleolus (Figure 2). It can therefore be speculated that nucleoplasmic RLCBs represent an intermediate form of nucleoplasmic CBs that become associated with nucleoli during differentiation of PC12 cells into a neuronal morphology. This possibility is supported by the absence of an increase in the number of both RLCBs and nucleolar caps in NGF-treated PC12 cells that failed to respond morphologically to NGF.
The association of CBs with nucleoli during NGF-induced neuronal differentiation may be a consequence of changes in nucleolar organization associated with nucleolar fusion, a process that has been shown to accompany neuronal differentiation (Buschmann and LaVelle 1981; Crespo et al. 1988; Park and De Boni 1991; Lafarga et al. 1995). This possibility is supported by evidence suggesting that CBs are frequently associated with nucleoli in mammalian cells that aggregate multiple NORs in one nucleolar domain (Brodsky et al. 1988; Lafarga et al. 1995). In fact, Lafarga and co-workers (1995) suggest that cells with a relatively high number of NORs, such as in rat and in mouse, have a strong tendency towards NOR aggregation. This leads to predominantly mononucleolated cells such as rat cerebellar Purkinje neurons, a cell type that often exhibits nucleolus-associated coiled bodies. Therefore, the increased association of CBs with nucleoli may reflect changes that are secondary to nuclear reorganization rather than to their functional interaction. In fact, such an association has been shown to occur in developing hippocampal neurons, independent of stage of differentiation and the characteristic transcriptional profile associated with that stage (Santama et al. 1996).
Given that CBs contain snRNPs (Fakan et al. 1984; Raska et al. 1991) and U3 snRNA (Jimenez–Garcia et al. 1994), components involved in pre-mRNA and pre-rRNA processing, it has been proposed that CBs function in either the assembly, disassembly, or storage of such components, or that they function directly in splicing reactions (Ochs et al. 1995). The results of the present study do support the existence of an association between CBs and nucleoli, and therefore favor a role for CBs in some aspect of pre-rRNA processing. Previous studies also support a role for CBs in assembly, dissasembly, or storage of components of pre-RNA processing. Analysis of CB number per nucleus in liver cells of hibernating mice revealed that CBs, which were abundant during hibernation, disappeared on arousal (Malatesta et al. 1994). These results are in keeping with the view that CBs have a role in the storage of components of pre-RNA processing. The results of the present study also support the concept of a role for CBs in storage of splicing components, because changes in the snRNP content of CBs occurred only with NGF-induced changes to a neuronal morphology and not in response to NGF-exposure alone, a feature that is potentially attributable to increases in rRNA metabolism, characteristic of neuronal differentiation (Lafarga et al. 1995).
The results of the present study, which show that CBs alter their morphology and exhibit an increased association with the nucleolus during NGF-induced neuronal differentiation, support the view that CBs are dynamic structures, undergoing cell state-dependent changes in morphology and in their intranuclear spatial distribution.
Footnotes
Acknowledgements
Supported by the National Science and Engineering Research Council of Canada.
The authors acknowledge the gift of R288 anti-coilin antibody from Dr E.K.L. Chan.