
Abstract
The microenvironment plays an important role in the homing in and differentiation of stem cells to repair injured tissue. Infrapatellar fat pad stromal cells (IFPSCs) are a promising source of such cells for the repair of articular injury-induced degeneration. This study investigated the chemotaxis of IFPSCs to chondrocytes and the effect of hyaluronan (HA) on the biological and regenerative properties of IFPSCs. The IFPSCs were obtained from patients undergoing arthroscopy and cultured via a standard 2-week culture protocol that yielded more than 10 million cells on passage 3. The results showed that the IFPSCs had a higher capacity for chondrogenic differentiation than mesenchymal cells from body fat, bone marrow, and Wharton's jelly of the umbilical cord. The IFPSCs cultured on 25% or 50% HA showed better osteogenic and adipogenic capabilities than those without HA or with 75% HA (p < 0.001). Cultures of the IFPSCs on 25% HA had a fourfold increase in chondrogenic differentiation compared to cultures without HA, which was better than with 50% and 75% HA (p < 0.05). Cell proliferation was not affected by the presence of HA. In conclusion, IFPSCs have a strong potential for chondrogenic regeneration, which can even be augmented in a 25% HA microenvironment.
Introduction
Osteoarthritis (OA) is a worldwide health problem among the elderly, affecting over 70% of Americans between the ages of 55 and 70 years (35). It is caused by progressive chondral damage due to chronic trauma or diseases (32). Articular cartilage has a limited capacity for repair because of the avascularity and low cellular mitotic activity (19). Several pathological mechanisms are involved, including enzymatic degradation of the extracellular matrix (ECM), deficient new matrix formation, cell death, and abnormal activation and hypertrophic differentiation of cartilage cells (15). Many treatment methods have been developed to decrease pain and improve the level of function of affected individuals. However, current strategies such as total joint arthroplasty are unable to restore the native structure of cartilage and may even increase the risk of further damage (2, 14). Consequently, recent research has focused on cell regeneration therapy with autologous stem cells (5).
Bone marrow mesenchymal stem cell (BMSC)-engineered bone and cartilage have been developed for OA treatment (12, 28). While this technique is appealing, the harvesting of bone marrow is painful and yields a limited number of stem cells, especially in the elderly. Thus, an extensive expansion in culture is required. Adipose-derived mesenchymal stem cells (ASCs) are currently the best choice since they can be obtained in larger amounts with less morbidity and can be easily expanded to large amounts in vitro (1, 13). In addition to a regenerative activity, ASCs have been shown to exhibit immunosuppressive properties. Recently, cells with stem cell characteristics have been reported in the infrapatellar fat pad (IFP) (9, 40). Owing to the orthotopic localization and practical accessibility, infrapatellar fat pad stromal cells (IFPSCs) have attracted interest as a potential source of cells for the repair of focal cartilage defects in the knee (22, 27, 30, 40).
The ECM provides a microenvironment for cells to maintain homeostasis and differentiate for specific tissues (3, 25). In the ECM, hyaluronan (HA) is the main glycosaminoglycan in the mesenchyme of early chondrogenesis and is native in cartilage tissue (16, 38). As the major physiological component of the articular cartilage matrix, HA is particularly abundant in synovial fluid. This polymer plays a role in cartilage homeostasis and is involved in cellular processes such as cell morphogenesis, proliferation, and wound repair. It is also widely used for the intra-articular treatment of OA (11, 34). However, the effect of HA microenvironment on MSC differentiation has rarely been reported (4).
The present study aimed to explore the role of the chondrogenesis ability of IFPSCs in a HA-rich microenvironment. Multipotent IFPSCs were derived from articular bone fat. These IFPSCs were more chondrogenic than ASCs, BMSCs, and Wharton's jelly stem cells (WJSCs), and were readily chemotactic to chondrocytes. They also showed enhanced differentiation capability in a 25% HA-coated culture.
Materials and Methods
Procurement of IFP and Articular Cartilage Tissue
Primary human articular cartilage fragments were isolated from an adult male undergoing arthroscopy. After obtaining approval from the Research Ethics Committee of the Buddhist Tzu Chi General Hospital and obtaining informed consent from the participants, viable IFPs were harvested during total knee arthroplasty surgeries as well as other sources of stem cells (BMSCs, ASCs, and WJSCs). The volume of IFPs was around 2 × 2 × 2 cm3. The patients (n = 2, both females) were both aged >60 years (65 and 68 years).
Derivation of Stromal Cells From Human IFPs (IFPSCs)
The IFPSCs were extracted from fat pads following a previously published protocol (43). Briefly, the harvested IFPs underwent a series of washes with phosphate- buffered saline (PBS; Biowest, Nuaille, France) and were digested with 0.1 mg/ml collagenase Ia (Sigma-Aldrich, St. Louis, MO, USA) at 37°C for 60 min. After enzyme digestion, the resulting cells were collected and cultured in keratinocyte serum-free medium (KSFM; Gibco, Grand island, NY, USA) with 5% fetal bovine serum (FBS; Biological Industry, Kibbutz, Israel), n-acetyl cysteine (NAC; Sigma-Aldrich), and L-ascorbic acid 2-phosphate (Sigma-Aldrich). The supernatant and debris in the culture dish were removed on the second day of culture, and the resulting IFPSC culture was denoted as passage 0. To prevent spontaneous differentiation, the cultures were maintained at subconfluent levels (<80% confluence). Passage of the cultures was performed using 2.5% trypsin/0.23 mM ethylenediaminetetraacetic acid (EDTA) (Gibco). XTT cell proliferation assays (Roche, Mannheim, Germany) were conducted in 96-well dishes at a cell density of around 2,000 cells/cm2 on days 0, 3, and 7.
Culture of BMSCs, ASCs, and WJSCs
BMSCs (n = 1) were provided by the bone marrow bank of our hospital. They were cultured with α-modified minimum essential medium (α-MEM; Gibco) supplemented with 10% FBS (Biological Industry). ASCs were derived from the subcutaneous tissue of a patient who underwent gynecologic surgery (n = 1, female, 60 years of age). Human adipose tissue was cut into small pieces (1–2 mm3), dissociated with 0.1 mg collagenase Ia, and incubated for 60 min at 37°C. The resulting cells were collected and cultured in KSFM (with the addition of epidermal growth factor and bovine pituitary extract; 17005-042; Gibco) with 5% FBS, NAC, and L-ascorbic acid 2-phosphate. Supernatant and debris were removed from the culture dish on day 2 of culture. To prevent spontaneous differentiation, the cultures were maintained at subconfluent levels (<80% confluency). The WJSC derivation process was the same as reported in our previous published paper (8). Briefly, the collected human umbilical cord tissues (n = 1) were mechanically cut by scissors in a midline direction, and the vessels of the umbilical arteries, veins, and outlining membranes were dissociated from the WJ. The jelly was then extensively cut into pieces smaller than 0.5 cm3, treated with collagenase type I (Sigma-Aldrich), and incubated for 14–18 h at 37°C in a 95% air/5% CO2 humidified atmosphere. The explants were then cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) containing 10% FBS and anti biotics at 37°C in a 95% air/5% CO2 humidified atmosphere. They were then left undisturbed for 5–7 days to allow for migration of the cells from the explants.
Flow Cytometry
Surface molecules of the IFPSCs and BMSCs cultured on the third or fourth passages were characterized by flow cytometry. The cells were detached with 2 mM EDTA in PBS, washed with PBS containing 2% bovine serum albumin and 0.1% sodium azide (Sigma-Aldrich), and incubated with the respective antibodies conjugated with fluorescein isothiocyanate or phycoerythrin, including cluster of differentiation 29 (CD29), CD34, CD44, CD45, CD73, CD90, human leukocyte antigen (HLA)-ABC, and HLA-DR (1:200; BD Bioscience, San Jose, CA, USA). Thereafter, the cells were analyzed using a Becton Dickinson flow cytometer (Becton Dickinson, San Jose, CA, USA).
Induction of Adipogenesis
The IFPSCs were seeded in a 12-well plate at a density of 5 × 104 cells per well with adipogenic medium (DMEM supplemented with 10% FBS), 1 μmol/L dexamethasone (Sigma-Aldrich), 5 μg/ml insulin (Sigma-Aldrich), 0.5 mmol/L isobutylmethylxanthine (Sigma-Aldrich), and 60 μmol/L indomethacin (Sigma-Aldrich). They were then allowed to grow for 7 days, with the medium being changed every 3 days, and then stained with Oil red O (Sigma-Aldrich). After washing twice with PBS, lipids in the sample were extracted with 1 ml 100% isopropanol (Sigma-Aldrich) and shaken gently for 5 min. The concentration of the lipids was measured based on absorbance at 510 nm (37). The quantity of lipids in each sample was measured in triplicate.
Induction of Osteogenesis
The IFPSCs were seeded in a 12-well plate at a density of 1 × 104 cells per well and grown with osteogenic medium [DMEM supplemented with 10% FBS, 0.1 μmol/L dexamethasone, 10 mmol/L β-glycerol phosphate (Sigma-Aldrich), and 50 μmol/L ascorbate (Sigma-Aldrich)], and changed every 3 days. The cells were allowed to grow for 21 days and were then stained with Alizarin red (Sigma-Aldrich). For quantification of the staining, 800 μl 10% (v/v) acetic acid (Baker, Phillipsburg, NJ, USA) was added to each well, and the plate was incubated at room temperature for 30 min with shaking. The monolayer, now loosely attached to the plate, was then scraped from the plate with a cell scraper (Corning Inc., Corning, NY, USA) and transferred with 10% (v/v) acetic acid to a 1.5-ml microcentrifuge tube with a wide-mouth pipette.
After vortexing for 30 s, the slurry was overlaid with 500 μl mineral oil (Sigma-Aldrich), heated to exactly 85°C for 10 min, and transferred to ice for 5 min. The slurry was then centrifuged at 20,000 × g for 15 min, and 500 μl of the supernatant was moved to a new 1.5-ml microcentrifuge tube (Scientific Specialties Inc., Lodi, CA, USA). Then, 200 μl of 10% (v/v) ammonium hydroxide (Baker) was added to neutralize the acid. Aliquots (150 μl) of the supernatant were read in triplicate at 405 nm in a 96-well (Costar; Corning Inc.) format using opaque-walled, transparent-bottomed plates (Costar; Corning Inc.) (18).
Induction of Chondrogenesis
The IFPSCs, ASCs, BMSCs, and WJSCs were seeded in a 12-well plate at a density of 1 × 105 cells/cm2 and grown in chondrogenic media consisting of DMEM, 10% FBS, 10 ng/ml transforming growth factor-b1 (Pepro Tech Inc., Rocky Hill, NJ, USA), 50 μg/ml ascorbic acid-2-phospate (Sigma-Aldrich), and 6.25 μg/ml of insulin (Sigma-Aldrich). The media were changed every 3 days. The cells were incubated with the chondrogenic media at 37°C with 5% CO2 for 3 weeks. After fixing in paraformaldehyde (Bionovas, Toronto, Canada), the cells were mounted on slides and stained using standard Alcian blue (Fluka, Sigma-Aldrich Chemie GmbH, Buchs, Switzerland) protocols. For quantification of the Alcian blue incorporation into the proteoglycan-rich ECM, the cultures were incubated with 6 M guanidine hydrochloride (Sigma-Aldrich) overnight and subjected to photometric measurement at an optical density of 595 nm (36).
RNA Extraction and Quantification
Total RNA was extracted using RNEasy® (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions. Real-time PCR analysis of sex-determining region Y box 9 (SOX9) and collagen type 2 a 1 (COL2A1) genes (primers and annealing temperatures listed in Table 1) was performed following the methods in previous reports (29, 41). Briefly, complementary DNA was synthesized using a SuperScript III One-Step RT-PCR kit (Invitrogen, Grand Island, NY, USA) and amplified by PCR with an AmpliTaq Gold Kit (Applied Biosystems, Foster City, CA, USA). Real-time PCRs were performed and monitored using FastStart universal SYBR green master (Rox; Roche, Indianapolis, IN, USA) and a quantitative real-time PCR detection system (ABI Step One Plus system; Applied Biosystems). The gene products were analyzed with the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene as a reference. The expression level of each target gene was then calculated as 2ΔΔCt, as previously described (29). Four readings of each experimental sample were performed for each gene of interest, and the experiments were repeated at least three times.
Sequences of Primers and Conditions Used in Real-Time Polymerase Chain Reaction

SOX9, sex-determining region Y box 9; COL2A1, collagen type 2 a 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Cell Culture in HA Microenvironment
Different amounts of hyaluronate solution (molecular weight 5,000–10,000 kDa, 20 mg/2 ml, Suplasyn; Bioniche, Galway, Ireland) dissolved in culture media were added to a 12-well plate for final concentrations (v/v) of 25%, 50%, 75%, and 100%, and allowed to solidify in room air. The IFPSCs were seeded at a density of 5,000 cells in 100 μl KSFM containing 5% FBS, NAC, and L-ascorbic acid 2-phosphate. The medium was changed every 2 days for 14 days. The cells were then collected for further differentiation and proliferation analysis at different time points.
Chemotaxis Migration Assay
Cartilage fragments were minced into 1-mm3 pieces and digested with type II collagenase (0.1%; Worthington, Lakewood, NJ, USA) solution overnight at 37°C. The digested contents were then filtered through a 100-μm filter and washed with PBS. The isolated chondrocytes were then plated at 5,000 cells per cm2 and grown to confluence with DMEM/F12 (Gibco) containing 2 mM l- glutamine and 10% FBS (Gibco), 1 × penicillin/streptomycin, 50 μg/ml ascorbic acid, and 0.1 M nonessential amino acids (Gibco, Invitrogen). Chondrocyte-conditioned medium was collected at 48 h by incubating chondrocytes with high-glucose DMEM (Gibco) further supplemented with 10% FBS and 1% penicillin/streptomycin.
The IFPSCs, BMSCs, ASCs, and WJSCs were seeded in the upper well of a 24-well Transwell Boyden chamber (8-μm pore size, Costar; Corning Inc.) and allowed to migrate toward cell-free media derived from chondrocytes placed in the bottom wells. Migration was assessed 48 h later. The migrated cells were stained with crystal violet (Sigma-Aldrich) and counted using bright field microscopy.
Statistical Analysis
The results were expressed as mean ± SD. The Student's t-test was used to evaluate mean differences between the control and experimental groups. A value of p < 0.05 was considered to be statistically significant. One-way ANOVA with post hoc Bonferroni test was used for multiple comparisons such as the proliferation rate of different stem cells.
Results
The IFPSCs Had a Phenotype Resembling Adipose-Derived Stem Cells
The derived IFPSCs had a fibroblast-like appearance with a high degree of homogeneity and readily reached confluence in 14 days of culture. Like other MSCs, the IFPSCs expressed surface markers of CD29, CD44, CD73, CD90, and HLA-ABC. However, unlike BMSCs, the IFPSCs expressed higher levels of CD34 and CD45 (Fig. 1). Compared to the BMSCs, WJSCs, and ASCs, the IFPSCs had cell surface markers resembling ASCs and unlike BMSCs (Fig. 1, Table 2). The growth kinetics of the BMSCs and WJSCs were significantly better than the ASCs on culture day 7 (p < 0.01 and p < 0.001, respectively) (Fig. 2). The proliferation of IFPSCs on day 7 compared with ASCs was not significant.

Cell surface markers of IFPSCs. Representative flow cytometry histograms of (A) IFPSCs, (B) BMSCs, (C) WJSCs, and (D) ASCs at passage 3 revealed that IFPSCs and ASCs were negative for HLA-DR, partially positive for CD34, CD45, and positive for CD29, CD44, CD73, CD90, and HLA-ABC.

Growth kinetics of IFPSCs, BMSCs, WJSCs, and ASCs from abdominal subcutaneous fat. The cell numbers of cultured IFPSCs, BMSCs, WJSCs, and ASCs were evaluated by XTT assay at days 0, 3, and 7. ***p < 0.001, **p < 0.01 by one-way ANOVA with post hoc Bonferroni test by comparing to ASCs.
Comparison of the Percentage of CD Markers in Different Kinds of Stem Cells

Abbreviations: CD, cluster of differentiation; IFPSCs, infra-patellar fat pad stromal cells; WJSCs, Wharton's jelly stem cells; BMSCs, bone marrow mesenchymal stem cells; ASCs, adipose-derived stem cells from abdominal subcutaneous fat.
The IFPSCs Were Chemotactic to Chondrocytes
The basic migration activity of the IFPSCs in the control medium was similar to the ASCs and WJSCs, but less than that of the BMSCs (Fig. 3). All of the cells, including the IFPSCs, were highly chemotactic to the conditioned medium of chondrocytes. Nonetheless, the IFPSCs had a migration ratio 2.95-fold higher than that of the control medium (Fig. 3).
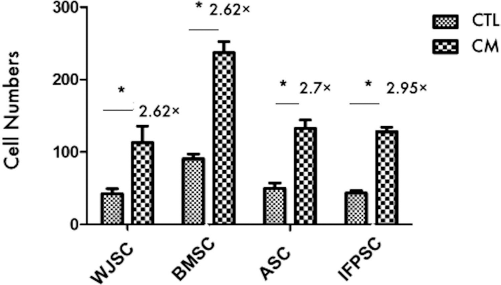
Mesenchymal stem cells of different origins were chemotactic to the conditioned medium (CM) of chondrocytes. The IFPSCs, BMSCs, ASCs, and WJSCs were subjected to Transwell migration toward the CM of chondrocytes or control medium for 18 h. The number of cells passing through the filter was counted after staining with crystal violet, and fold changes of migrated cell number were shown. IFPSCs, infra-patellar fat pad stromal cells; WJSCs, Wharton jelly stem cells; BMSCs, bone marrow mesenchymal stem cells; ASCs, adipose-derived stem cells from abdominal subcutaneous fat. *p < 0.05.
The IFPSCs Could Differentiate Into Fat, Bone, and Cartilage
Upon induction of differentiation in culture, the IFPSCs readily differentiated to fat, bone, or cartilage lineages. By 14 days postinduction in adipogenic and osteogenic conditions, the differentiated IFPSCs showed large, Oil red O-positive lipid droplets within the cytoplasm (Fig. 4A) and became positive for Alizarin red staining with a change of cell morphology to a cuboid shape (Fig. 4B). By 21 days of chondrogenic induction, the IFPSCs displayed cell aggregation (Fig. 4C) with strong Alcian blue staining (Fig. 4D).

Multipotent differentiation capability of IFPSCs. The IFPSCs could be induced for (A) adipogenesis, as shown by Oil red O staining; (B) osteogenesis, as shown by Alizarin red-positive extracellular matrix calcification; and (C, D) chondrogenesis as shown by Alcian blue stain. Scale bar: 100 μm in (A) and (D), 1,000 μm in (C).
The IFPSCs were more Chondrogenic than the Mesenchymal Stem Cells Derived from other Sites
Upon induction of chondrogenic differentiation, on day 21, the chondrogenic-induced IFPSCs expressed much larger amounts of glycosaminoglycans (Alcian blue) than the induced BMSCs, ASCs, and WJSCs (p < 0.001) (Fig. 5A, B). The IFPSCs progressively expressed chondrogenic genes such as SOX9 and COL2A1 (Fig. 5C, D). In comparison, these two genes were either expressed less (ASCs and WJSCs) or were inconsistently expressed at different durations of induction (BMSCs) (Fig. 5C).
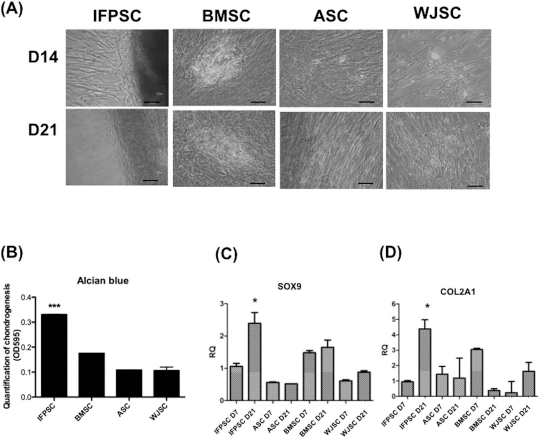
The IFPSCs were more chondrogenic than other mesenchymal stem cells. The capability for chondrogenesis of various mesenchymal stem cells was shown by (A) Alcian blue stain, and the amount of staining at day 21 was measured by (B) spectrophotometry at OD595. ***p < 0.001. Expressions of (C) SOX9 and (D) COL2A1 at different induction days were measured by qRT-PCR. *p < 0.05.
The HA Microenvironment Enhanced the Differentiation of IFPSCs, But Was Neutral to Its Growth
At different concentrations, HA was coated on the culture wells and tested for the effect on the differentiation and growth of IFPSCs. Other than the 75% HA-coated culture, which showed a poor differentiation capability, the IFPSCs cultured on either 25% or 50% HA had enhanced osteogenic, adipogenic, and chondrogenic differentiation. For chondrogenesis, the IFPSCs cultured on 25% HA showed the best capacity for differentiation, with two- to threefold increases compared to the culture without HA (p < 0.05) (Fig. 6). Moreover, cultures with HA coatings of concentrations varying from 25% to 75% did not affect the proliferation of the IFPSCs (Fig. 7).

HA 25% enhanced adipogenesis, osteogenesis, and chondrogenesis of human IFPSCs. (A) Gross pictures and (B) amount of osteogenesis, adipogenesis, and chondrogenesis of IFPSCs cultured at different concentrations of HA were shown as indicated by alizarin red, oil red, and alcian blue staining, respectively. At an HA concentration of 25%, IFPSCs showed the best differentiation capability for all three cell lineages, compared to other HA concentrations. *p < 0.05, **p < 0.01, ***p < 0.001.

HA had no effect on IFPSC proliferation. (A) Gross pictures of IFPSCs after 5 days of culture with 25%, 50%, 75%, and 100% of HA. (B) Proliferation of IFPSCs was not affected by HA at different concentrations. Scale bar: 1,000 μm.
Discussion
This study isolated and investigated stromal cells from IFPs for the chemotaxis to chondrocytes, capability for differentiation, and effect of HA on the regenerative characteristics. There were several major findings. First, isolated stromal cells from the IFPs could be prepared within a short time frame. The cultured IFPSCs resembled ASCs in terms of growth kinetics, cell surface marker profile, and multidifferentiation capacity. Lastly, the IFPSCs seeded in HA-coated dishes showed better differentiation capability, including significantly enhanced chondrogenesis and formation of chondrogenic ECM.
Articular cartilage contains a cell density ranging from 6,800 to 24,000 cells/mm3 (19), and regeneration toward cartilage of this density may require an estimated 1.6 × 107 cells for repair of a 600-mm3 area of cartilage (23). The time to prepare IFPSCs for this therapeutic utility was estimated in a culture system, and resection and manual processing of the IFPs took approximately 1 h. Subsequent cultivation of IFPSCs in the laboratory took 4–5 days to grow to subconfluency and another 10 days (at passage 3) to expand to 1.6 × 107 cells. This preparation can be accomplished within 2 weeks. Moreover, the quality and history of the implanted cells may influence regeneration outcomes, although the optimal cell-seeding density for cartilage tissue engineering has yet to be determined (22).
The presence of stromal cells with multipotent differentiation activity in IFPs was first described by Wickham et al. (40). Other studies have shown that these cells can differentiate into chondrocytes, osteoblasts, and adipocytes (9, 10, 24). Later, the freshly isolated stromal vascular fraction of IFP tissue was also proven to be applicable for one-step transplantation after cultivation (22). In the present study, both the cell surface marker profile and growth kinetics of the IFPSCs were in concordance with ASCs from abdominal subcutaneous fat tissue (21, 39) (Table 2). As in previous studies (22), there were relatively higher levels of CD34 and CD45 in the IFPSCs (9.0% and 11.0%) than in the BMSCs (0.3% and 0.5%). Although the IFPSCs shared most of the surface markers as the ASCs, the CD45 level in the IFPSCs was much higher (0.4%). The constitution of fat pad tissue may differ from that of subcutaneous fat, since the former has significant amounts of dense connective tissue adjacent to the cartilage (40).
In the presence of chondrogenic media, IFPSCs are induced to express glycosaminoglycans, which are major components of the ECM of chondrogenic tissue and can be stained by Alcian blue. Compared to other mesenchymal cells such as ASCs, BMSCs, and WJSCs, IFPSCs show significantly more Alcian blue staining upon chondrogenic induction, as well as higher expressional levels of chondrogenic genes such as SOX9 and COL2A1. In addition, IFPSCs are induced to express these genes earlier than other mesenchymal cells. By day 21 of chondrogenic differentiation, SOX9 and COL2A1 were significantly upregulated in the IFPSCs (p < 0.05), but not in the ASCs or BMSCs, and only mildly increased in the WJSCs (p > 0.05). Thus, there is a propensity of differentiation of mesenchymal stem cells into a tissue of its orthotopic origin.
The first concern after stem cell transplantation is where the cells go. The present study also demonstrated the chemotactic activity of the IFPSCs toward growing chondrocytes, indicating a potential homing of IFPSCs toward the injured cartilage where the growing chondrocytes are located. This chemotactic effect did not seem to be tissue specific, since the mesenchymal cells of nonarticular origin also did well. Recently, conditioned medium of chondrocytes has been shown to enhance the chondrogenesis of ASCs (20). Thus, growing chondrocytes in the injured cartilage may provide a good microenvironment to attract the mesenchymal stem cells for repair.
The age of the donor, number of passages of the cells, and culture media can affect the stem cell characteristics. We previously showed that the donor's age does not affect the proliferation rate, doubling time, telomere length, or osteogenic and chondrogenic differentiation capacity of ASCs (6). In our previous study, MSCs cultured at passages 3 to 6 had stable and comparable cell properties to progenitor cells (6–8). Replication senescence often occurred after passage 10. The most suitable culture medium for each kind of MSC varies (6, 8, 31) and may affect the comparison of their cell properties. In this study, the BMSCs were cultured with α-MEM, the IFPSCs and ASCs were cultured with KSFM, and the WJSCs were cultured with DMEM low glucose. Other than a lower proliferation of BMSCs, we did not see any significant differences in cell proliferation and chemotaxis activity among these cells. This suggests that conditioned medium has a neutral effect on IFPSC proliferation, although this effect warrants further study.
In the current study, various concentrations of HA had no promoting effect on the proliferation of IFPSCs compared with the control. A previous study reported that 150 μg/ml of HA could stimulate cell division (17). In the current study, the studied concentrations of HA were 10 mg/ml (100%), 7.5 mg/ml (75%), 5 mg/ml (50%), and 2.5 mg/ml (25%), which were more than 20-fold higher.
HA may have a potent enhancing effect on the chondrogenesis of IFPSCs. As a native component of cartilage tissue, HA represents the natural microenvironment for the derivation of chondrocytes from their progenitors in the joint. HA is essential in the ECM for cell-to-cell cross bridging for cell aggregation prior to condensation of pre-cartilage (26). HA is also the major component of synovial fluid (33), which surrounds IFP. Therefore, HA in the synovial fluid may contribute to a microenvironment that can induce chondrocytic differentiation of IFPSCs during repair of the injury. Consistent with this hypothesis, the findings of this study showed that HA significantly enhanced various differentiation capabilities of IFPSCs, including the enhancement of chondrogenesis and osteogenesis, which were more prominent than adipogenesis, reflecting the origin of this mesenchymal cell as well as HA from the joint. An optimal concentration of 25% worked best for the chondrogenic differentiation. These results indicate that HA provides a suitable microenvironment for a proper differentiation of IFPSCs.
HA has also been shown to enhance the chondrogenic differentiation of ASCs (41). The authors suggested that the total effect of chondrogenesis of IFPSCs in 25% HA is greater than that in ASCs, since the latter has a higher baseline chondrogenic potential in the absence of HA. In contrast, a previous study reported that a higher concentration of HA (0.5 mg/cm2) led to more chondrogenesis than lower concentrations (0.05 and 0.005 mg/cm2) (41). In the current study, a much higher concentration (7.5 mg/ml, 75% = 3.9 mg/cm2) of HA was used compared to the previous studies, and this may be responsible for the difference in differentiation capacity.
The present study also demonstrated a high level of CD44 surface antigen in IFPSCs. CD44 is the primary receptor for HA (26) and has been reported to mediate the chondrogenic differentiation in ASCs (42). However, signal transduction during IFPSC chondrogenesis through CD44 is not well understood. The molecular mechanism of the interaction between CD44 and HA on initiating differentiation of IFPSCs requires further investigations.
Intra-articular injections of HA are a common practice in orthopedics. This study suggests a combined injection of IFPSCs and 25% HA to exert a better effect of chondrogenesis. The results here provide evidence of the beneficial effects of HA supplementation in future preclinical studies of intra-articular injections of IFPSCs for the regeneration of damaged joint cartilage.
Conclusions
The present study reveals that IFPSCs derived from the fat pads in human joints have a higher chondrogenic capability than other mesenchymal stem cells. These IFPSCs are chemotactic to growing chondrocytes and show an enhanced differentiation capability in a HA microenvironment. The results emphasize that a HA-enhanced microenvironment may be applied to IFPSC-mediated cellular therapy for more effective articular cartilage repair.
Footnotes
Acknowledgments
The authors sincerely thank the National Science Council of the Republic of China, Taiwan, for financially supporting this research (Contract No. NSC 98-2314-B-303-009-MY3), and the Buddhist Tzu Chi General Hospital, for the financial support (Contract Nos. TCRD99-12 and TCSP98-07). The authors declare no conflicts of interest.