
Abstract
Cellular fate of human neural stem cells (hNSCs) transplanted in the brain of nonhuman primates (NHPs) with no immunosuppression was determined at 22 and 24 months posttransplantation (PTx) regarding survival, differentiation, and tumorigenesis. Survival of hNSCs labeled with magnetic nanoparticles was successfully detected around injection sites in the brain at 22 months PTx by MRI. Histological examination of brain sections with H&E and Prussian blue staining at 24 months revealed that most of the grafted hNSCs were found located along the injection tract. Grafted hNSCs were found to differentiate into neurons at 24 months PTx. In addition, none of the grafted hNSCs were bromodeoxyuridine positive in the monkey brain, indicating that hNSCs did not replicate in the NHP brain and did not cause tumor formation. This study serves as a proof of principle and provides evidence that hNSCs transplanted in NHP brain could survive and differentiate into neurons in the absence of immunosuppression. It also serves as a preliminary study in our scheduled preclinical studies of hNSC transplantation in NHP stroke models.
Keywords
Introduction
Stroke is the fourth major cause of death in the US after heart failure, cancer, and lower respiratory disease, and ischemic stroke represents 87% of the total stroke (31). In ischemic stroke, administration of tissue plasminogen activator (TPA) within 3 h after the stroke clinical onset has shown good outcomes. However, only a small number of ischemic stroke patients are helped by TPA treatment, and for the majority of ischemic stroke patients alternative treatment is needed. Stem cell therapy may have the potential to address this need.
Starting in the late 1990s, stem cells received wide attention because of their therapeutic potential in repairing or replacing injured or diseased human tissues and cells. Stem cells are defined as cells that have the ability to renew themselves continuously and possess pluripotent ability to differentiate into many cell types. Embryonic stem cells (ESCs), derived from the inner cell mass of blastocysts, have been identified as stem cells that give rise to various organs, tissues, and cells (34). Recently, there has been an exciting development in generation of a new class of pluripotent stem cells (PSCs): induced pluripotent stem cells (iPSCs) from adult somatic cells such as skin fibroblasts by introduction of embryogenesis-related genes (30,33,37). In addition to ESCs and iPSCs, tissue-specific stem cells could be isolated from various tissues of more advanced developmental stages, such as hematopoietic stem cells, bone marrow mesenchymal stem cells, adipose tissue-derived stem cells, amniotic fluid stem cells, and neural stem cells (NSCs). Among these, existence of multipotent NSCs has been known in developing or adult rodent and human brain with properties of indefinite growth and multipotent potential to differentiate into three major cell types of the CNS: neurons, astrocytes, and oligodendrocytes (5–7,14,15,26,27).
NSCs should be of great value as the source for the cell replacement and gene transfer for the treatment of human neurological disorders such as Parkinson's disease (PD), Huntington's disease (HD), Alzheimer's disease (AD), amyotrophic lateral sclerosis (ALS), spinal cord injury (SCI), and stroke. We have previously demonstrated that human NSCs (hNSCs) transplanted in rodent models of PD (17,35), HD (25,32), AD (23,29), ALS (10), SCI (9,14), and stroke (2,11,20–22,24) survived and differentiated into neurons and induced functional recovery. However, for the future clinical application of the hNSCs, long-term safety, survival, and differentiation of the hNSCs should be investigated in nonhuman primates (NHPs). In the case of neuroprotective stroke therapies, all neuroprotective compounds thought to be effective in rodent stroke models have been ineffective in humans (12,28). The anatomical and physiological differences between the rodent and human brain were cited as reasons for clinical failures (1,3,4,8). Thus, it is essential to provide evidence of promise for hNSC-based cell therapy in human trials in patients with neurological diseases, and such evidence requires preclinical studies in NHPs.
In the present study, we grafted hNSCs without immunosuppression in the brains of NHP cynomolgus monkeys and evaluated the fate of grafted hNSCs at 24 months posttransplantation (PTx) regarding survival, differentiation, and potential tumorigenesis via MRI, histopathological, and immunohistochemical analyses. This study was performed as a proof-of-principle study to provide evidence that hNSCs transplanted in the NHP brain could survive and differentiate into neurons in the absence of immunosuppression.
Materials and Methods
Human Neural Stem Cell Line
A stable clonal human NSC line, HB1.F3 (F3), was generated by retroviral transduction of primary fetal hNSCs with a retroviral vector-encoding v-myc as previously reported (16,19). F3 hNSCs were grown in a serum-free medium (DM4) consisting of Dulbecco's modified Eagle medium with high glucose (DMEM; Invitrogen, Grand Island, NY, USA) containing 10 μg/ml insulin, 10 μg/ml transferrin, 30 nM sodium selenate, 50 nM hydrocortisone, 0.3 nM triiodothyronine, and 20 μg/ml gentamicin. Recombinant human fibroblast growth factor 2 (FGF-2, 10 ng/ml; PeproTech, Rocky Hill, NJ, USA) was supplemented to the DM4 during the routine feeding. All chemicals except FGF-2 were obtained from Sigma (St. Louis, MO, USA).
RT-PCR
PCR Primer Sequences for Cell Type-Specific Markers (all Human)

NGF, nerve growth factor; NF-L, neurofilament light; NF-M, neurofilament medium; GFAP, glial fibrillary acidic protein; CNPase, 2′,3′-cyclic-nucleotide 3′-phosphodiesterase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Labeling of hNSCs with Fluorescent Magnetic Nanoparticles (MNPs)
Fluorescent silica magnetic nanoparticles (MNPs) 50 nm in size and containing rhodamine isothiocyanate (RITC) conjugated to terminal silanol groups (NEO-STEM TSR50, MNP) were purchased from Biterials (Seoul, Korea). F3 hNSCs were incubated in DMEM-10% fetal bovine serum (FBS) medium containing NEO-STEM TSR50 (RITC, 1 mg/ml) overnight in a 37°C, 5% CO2 incubator, followed by incubation in DMEM-10% FBS medium for 6 h and washing in saline before injection. MNP labeling of F3 hNSCs was confirmed by fluorescence microscopy (IX70; Olympus, Tokyo, Japan).
Experimental Animals
Two 6-year-old female cynomolgus monkeys (2.8 ~ 3.0 kg; Korea National Primate Research Center, Ochang, Korea) were used and housed in an indoor individual cage [60 (W) × 80 (L) × 80 (H) cm3] and fed commercial monkey chow (Harlan Laboratories, Indianapolis, IN, USA) supplemented daily with various fruits and were supplied water ad libitum. Environmental conditions were controlled to provide a temperature of 24 ± 2°C, a relative humidity of 50 ± 5%, 100% fresh air at a rate of ≥12 room changes per hour, and a 12:12 h light/dark cycle. All housing and experiments were performed in accordance with the Korea Research Institute of Bioscience and Biotechnology Institutional Animal Care and Use Committee Guidelines.
Transplantation of Neural Stem Cells
Cynomolgus monkeys were anesthetized by intramuscular injection of 5 mg/kg ketamine HCl (Yuhan, Seoul, Korea) followed by inhalation of 2% isoflurane (JW Pharmaceutical, Seoul, Korea). After the skin incision, the frontal skull was exposed and then drilled about 1 mm in diameter. Through the craniotomy site, a total of 20 μl (1 × 106 cells) of hNSC suspension was injected for 10 min with a Hamilton syringe (fixed needle syringe, 87942; Hamilton, Reno, NV, USA) in the brain at a depth of 1 cm from the skull. The needle was removed after remaining in place for another 5 min. One monkey received MR detectable MNP-labeled hNSCs, and the other received unlabeled hNSCs. After suture of muscle and skin, monkeys were treated with general postoperation medication, such as antibiotics, but without any use of an immunosuppressant.
MRI Procedure
Representative MRI Protocols for Diagnostic Image Analyses

MRI, magnetic resonance imaging; FLAIR, fluid-attenuated inversion recovery; SWI, susceptibility weighted imaging; TR, time to repeat; TE, time to echo; TI, time to inversion; FOV, field of view.
Histology and Immunohistochemistry
Cynomolgus monkeys were sacrificed at 24 months PTx and brains removed for histology. For tissue preparation, animals were anesthetized and perfused via carotid artery with phosphate-buffered saline (PBS; Sigma) followed by 10% neutral-buffered formalin (NBF; Sigma). The brains were removed and fixed additionally in 10% NBF for 1 week. The brains were embedded in paraffin and sectioned at 5 μm in thickness using a microtome (HM325; Thermo Scientific, Tewksbury, MA, USA). Serial sections were processed for hematoxylin and eosin (H&E; Sigma), Prussian blue staining, and immunohistochemistry. Twelve days prior to sacrifice, the same monkeys were injected with bromodeoxyuridine (BrdU, 50 mg/kg; Sigma) dissolved in 0.9% NaCl daily intravenously for 5 consecutive days and the animals sacrificed 7 days later.
To detect MNP-labeled hNSCs in the brain, brain sections were processed for Prussian blue staining (1.2% potassium ferrocyanide with 1.8% hydrochloric acid; both from Sigma) (13). Brain sections were processed for double immunostaining with anti-human mitochondria antibody (hMit, 1:200; Chemicon International, Temecula, CA, USA) and GFAP (1:1,000; Chemicon) or neurofilament heavy (NF-H) protein (1:200; Chemicon). To determine whether grafted F3 cells continue to proliferate in vivo, brain sections were processed for double immunofluorescence staining of hMit and BrdU (1:100; Chemicon). Brain sections were incubated in primary antibodies overnight at 4°C as free-floating sections, followed by Alexa Fluor 484 anti-mouse IgG or Alexa Fluor 594 anti-rabbit IgG (1:500; Molecular Probes, Eugene, OR, USA) for 1 h at RT. Negative control sections were prepared in an identical manner except that the primary antibodies were omitted. Stained sections were then examined under an Olympus confocal fluorescence microscope (Olympus).
Results
Characterization of Human Neural Stem Cells
F3 hNSCs are bipolar or tripolar in morphology and are 8–10 μm in size (Fig. 1A). Immunocytochemical staining demonstrated that F3 hNSCs expressed phenotypes specific for NSCs, including nestin and Musash-1, as reported previously (16,19). Cytogenetic analysis of F3 NSCs showed a normal karyotype of human cells with a 46, XX, without any chromosomal abnormality as previously described (Fig. 1B). Results of RT-PCR analysis for mRNA isolated from F3 hNSCs grown in serum-containing medium (10% FBS) are shown in Figure 1C. The transcript for nestin, a cell type-specific marker for NSCs, transcripts for NF-L and NF-M, cell type-specific markers for neurons, transcript for GFAP, a cell type-specific marker for astrocytes, and transcript for CNPase, a cell type-specific marker for oligodendrocytes, are all expressed by NSCs and their progeny (Fig. 1C). These results indicate that F3 hNSCs grown in serum-containing medium undergo asymmetrical division by which one daughter cell remains a NSC and continues cell division, while another one undergoes terminal differentiation into a neuron or glial cell.
Basic characteristics of hNSCs. (A) F3 hNSCs under phase-contrast microscopy are bipolar or tripolar in morphology. Scale bar: 20 μm. (B) Cytogenetic analysis indicates that F3 hNSCs carry normal human karyotype of chromosomes, 22 pairs of autosomes, and two X chromosomes. (C) Gene expression of cell type-specific markers in F3 hNSCs as studied by RT-PCR. F3 hNSCs express cell type-specific markers nestin (for neural stem cells), NF-L, and NF-M (both for neurons), GFAP (for astrocytes), and CNPase (for oligodendrocytes).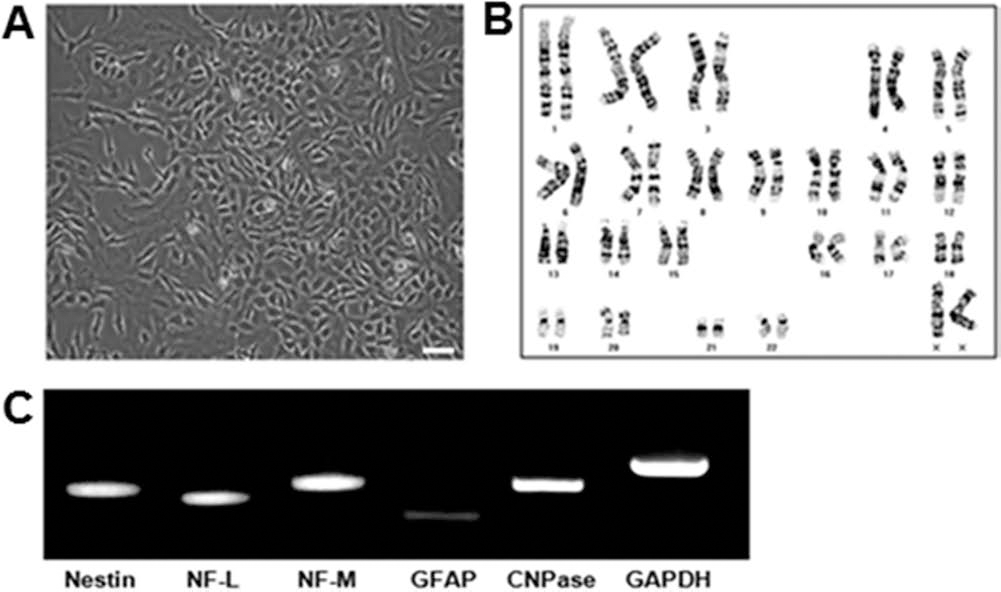
Detection of Grafted hNSCs in NHP Brain by MRI
To detect grafted hNSCs in the monkey brain with in vivo MR imaging, hNSCs were labeled with iron MNPs before the transplantation in a monkey. MRI showed hNSCs labeled with MNPs at 3 days and 22 months after hNSC injection in the monkey brain (Figs. 2, 3). No pathological changes were detected on MRIs at 3 days and 22 months post-hNSC transplantation (Figs. 2, 3).
MRIs of the cynomolgus monkey brain at 3 days after the hNSC injection. (A) TSE T2 images show linear low-signal intensity caused by MNPs (black arrows). hNSCs were labeled with MNPs before the transplantation. (B) T1 and T2* MR images do not visualize the presence of unlabeled hNSCs. Note soft tissue injuries due to the surgical wounds (white arrows). MR images of cynomolgus monkey brain at 22 months after injection of hNSCs. (A) T2, SWI, and FLAIR MRI images show the linear low signal caused by needle injection tract (arrows) of MNPs. (B) Unlabeled hNSCs could not be detected. There was no evidence of brain tumor or any other brain lesion in both monkeys.

Long-Term Survival of hNSCs in NHP Brain with no Immunosuppression
To detect survival and differentiation of grafted hNSCs in the monkey brain without immunosupression at 24 months PTx, brain tissues were analyzed histopathologically and immunohistochemically. In the histopathological analysis, many hNSCs with MNPs were detected around the injection tract (Figs. 4, 5). In the brain sections processed for H&E and Prussian blue staining, most of the grafted hNSCs were found to migrate from the injection site to other brain parenchyma. Interestingly, all hNSCs survived without immunosuppression (Figs. 4, 5). No histopathological changes such as tumor formation were detected in the monkey brains examined.
Histological analyses in the cynomolgus monkey brain at 24 months after injection of MNP-labeled hNSCs. (A) Injection needle tract is noted and filled with erythrocytes. Scale bar: 500 μm. (B and C) Grafted hNSCs (arrowheads) are evident in the magnified square. Scale bars: 100 and 20 μm, respectively. Prussian blue staining in the cynomolgus monkey brain at 24 months after injection of MNP-labeled hNSCs. (A) Prussian blue-positive hNSCs are located around the injection tract. Scale bar: 500 μm. (B and C) In the magnified square, grafted hNSCs are clearly shown. Scale bars: 100 and 20 μm (the border line is artificial.).

Differentiation of Grafted hNSCs in NHP Brain
Following transplantation into monkey brains, hMit (anti-human mitochondria antibody)-positive hNSCs were found selectively along the injection sites (Fig. 6). A large number of hMit/NF double-positive cells were found in the brain sections along the injection site (Fig. 6A–C), while no hMit+/GFAP+ cells were detected (Fig. 6D–F). These results indicate that a majority of grafted F3 hNSCs differentiated into neurons in response to signals provided by the local microenvironment at the injected sites. To detect possible replication of grafted hNSCs, BrdU was injected for 5 consecutive days and the animals sacrificed 7 days later. The hMit+ cells were all negative for BrdU staining, indicating that the grafted hNSCs did not undergo replication in vivo (Fig. 6G–I).
Immunohistochemical analyses in the cynomolgus monkey brain at 24 months PT of F3 hNSCs. Double immunofluorescence staining of engrafted F3 hNSCs in the monkey brain (A–I). hMito-positive F3 hNSCs are also NF positive, indicating that F3 hNSCs differentiated into NF-positive neurons (A–C), but hMito-positive hNSCs are GFAP negative (astrocyte marker) (D–F). hMito-positive hNSCs are BrdU negative, indicating that the F3 NSCs did not undergo replication in the monkey brain at 24 months PT (G–I). Scale bar: 50 μm.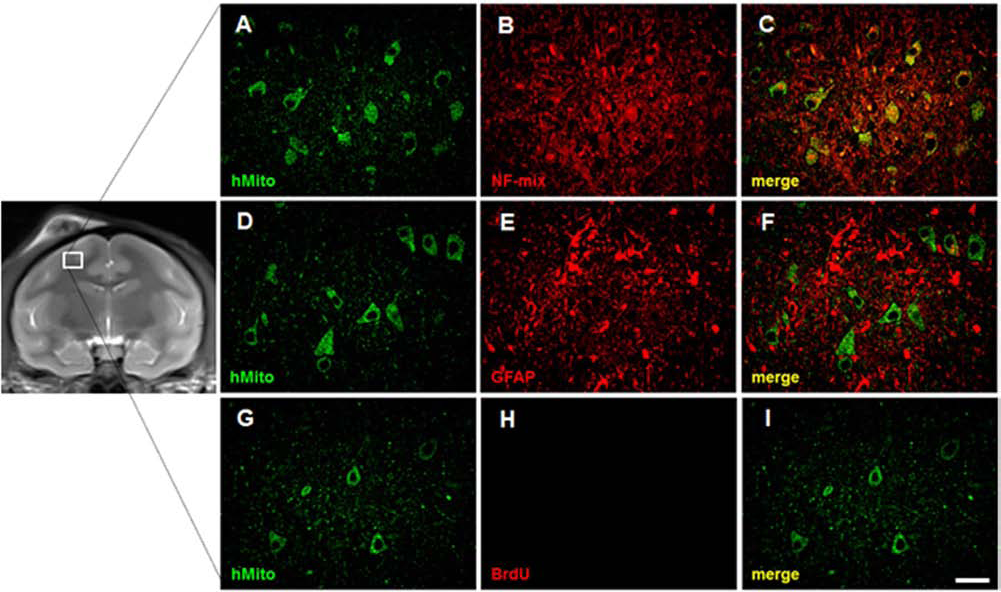
Discussion
The present study reported the long-term survival and neuronal differentiation of grafted hNSCs in the brain of cynomolgus monkeys with no immunosuppression. At 24 months PTx, we were able to detect the hMit-positive hNSCs labeled with nanoparticles, and there was no evidence of immune rejection or tumorigenesis in the NHP brain. Literature searches indicate that this study is the first study reporting the long-term survival of hNSCs in the brain of NHPs without the use of an immunosuppressant.
The effect and the necessity of immunosuppression to elude host immune rejection of a xenograft are controversial. Generally, immunosuppressive treatment in the xenograft transplantation is well accepted as a routine procedure. However, survival of transplanted neural precursor cells or NSCs without immunosuppression in rodents has often been reported. For example, xenografted hNSCs in the rodent brain without any immunosuppression survived successfully for 4–12 weeks in rodent models of PD, HD, AD, ALS, SCI, and stroke (2,11,17,20–25,29,32,35). In the present study, hNSCs transplanted into the brain of cynomolgus monkeys survived successfully for 24 months without the use of an immunosuppressant, and most of the grafted hNSCs had differentiated into neurons. The mechanism by which grafted hNSCs elude immune rejection is not known.
For in vivo detection and follow-up of xenografted hNSCs with MRI, MNPs were used. Safety and feasibility in the application of MNPs as the cellular marker were relatively well known. In the right cerebral hemisphere of one cynomolgus monkey, we were able to locate hNSCs bearing MNPs along the needle tract initially at 3 days after the xenograft (Fig. 2). At 22 months PTx, a follow-up MRI demonstrated surviving hNSCs with MNPs and no evidence of xenograft-associated brain tumors or brain lesions was found (Fig. 3). Magnetic susceptibility artifacts from the MNPs were found along the needle injection tract (Figs. 2A, 3A). Histopathological analysis at 24 months PTx revealed that MNPs exist in the cytoplasm of the grafted hNSCs (Figs. 4, 5). These results suggest that MNPs used in this study are compatible with the F3 hNSCs in the cynomolgus monkey brain. When MNPs were detected on the long-term follow-up MRI, it indicated that hNSCs were alive in the cynomolgus monkey brains.
In the present study, hNSC transplantation was conducted as a “free hand” surgical implantation into the brain parenchyma. In the planned future studies, however, intracerebral transplantation should be conducted in a steretaxic guided surgery to assure exact placement of cell transplantation.
We injected hNSCs into the frontal lobe and the putamen of the monkey brain because they are included in the middle cerebral artery (MCA) territory, which is the main target in the development of ischemic lesion in animal stroke models. Thus, research on survival and differentiation of hNSCs in the MCA territory should provide more meaningful information about cell transplantation in the MCA occlusion stroke model.
BrdU is a thymidine analog that is incorporated into replicating DNA of dividing cells, and thereby the BrdU-positive immunoreaction is taken as a specific indicator for the cellular proliferation (18,36). In this study, BrdU was injected 5 days before euthanasia of the monkeys. No BrdU-positive hNSCs were found in the NHP brain (Fig. 6H). No sign of tumor formation was detected in the brain (Figs. 4, 5). In addition, karyotyping of hNSCs did not show any major chromosomal abnormalities (Fig. 1). These results suggest that xenografted hNSCs did not replicate in the NHP brain, and there is very low possibility of tumor formation by the grafted hNSCs in the NHP brain.
Since this study deals with the fate of the hNSCs following transplantation in the normal NHP brain, it might work differently in an injured or damaged brain. However, this is the first study to show the long-term survival and differentiation of the hNSCs without the need for immunosuppression. Our study has direct clinical implications for the use of hNSCs in human patients with brain diseases and injuries, as well as safety of cell therapy. Furthermore, we are conducting experimental studies of NHP models for brain diseases such as stroke, PD, and AD.
In conclusion, grafted hNSCs in the brain of cynomolgus monkeys survived for 22–24 months in the absence of immunosuppression and differentiated into neurons. Their long-term survival was identifiable with the use of MNPs by MRI follow-up. MRI, histopathological, and immunohistochemical analyses at 24 months PTx showed no evidence of tumor formation by the grafted hNSCs. The results of this study serve as a proof of principle to provide evidence that human NSCs transplanted in the NHP brain could survive and differentiate into neurons in the absence of immunosuppression. It also serves as a preliminary study in our planned preclinical studies of hNSC transplantation in NHP stroke models.
Footnotes
Acknowledgments
This study was supported by grants from Chungcheongbuk-Do (LCC0021211), KRIBB Research Initiative Program (KGM4611512), and NRF Research Program funded by the MEST, Republic of Korea (20100023426). The authors declare no conflicts of interest.