
Abstract
Cell therapies consist in transplanting healthy cells into a disabled tissue with the goal to repopulate it and restore its function at least partially. In muscular diseases, most of the time, myoblasts are chosen for their expansion capacity in culture. Nevertheless, cell transplantation has limitations, among them, death of the transplanted cells, during the days following the graft. One possibility to counteract this problem is to enhance the proliferation of the transplanted myoblasts before their fusion with the existing muscle fibers. AG490 is a specific inhibitor of janus tyrosine kinase 2 (JAK2). The hypothesis is to block myoblast differentiation with AG490, thus permitting their proliferation. The inhibition of myoblast fusion by AG490 was confirmed in this study by gene expression and with a myosin heavy chain staining (MyHC). Moreover, cell survival was estimated by flow cytometry. AG490 was found to protect myoblasts in vitro from apoptosis induced by H2O2 or by preventing attachment of cells to their substrate. Finally, in an in vivo model of muscle regeneration, when AG490 was coinjected with the myoblasts their survival was increased by 45% at 5 days after their transplantation.
Introduction
Cell-based therapies have been used in recent clinical trials of regenerative medicine, particularly for myocardial injury (17, 31, 42) and muscular diseases (6, 20, 34–36, 40, 41). In these cases, the technique consists of transplanting healthy cells into the injured muscle. The objective of the cell transplantation is to repopulate the muscle, promote muscle regeneration, and improve muscle function (6). Myoblasts are often chosen for these cell-based approaches (17, 20, 34, 39, 42) because they can be prepared from a simple muscle biopsy from the host themselves for cardiac therapy (autologous transplantation) or from a cadaver or a related donor in muscular therapy (allogenic transplantation). Myoblasts result from the proliferation and differentiation of satellite cells, which are muscle-specific stem cells (34). One great advantage of using myoblasts for cell therapy is their high proliferating capacity in culture before their transplantation (23, 41).
However, myoblast-based therapy has some limitations. The first limitation is the poor capacity of the transplanted cells to migrate from the site of injection and fuse with distant fibers (34, 36, 41). Another problem of myoblast transplantation is the rapid and massive death of the transplanted myoblasts (34, 36, 39, 41). Previous studies have suggested that the majority of myoblasts grafted into skeletal muscles or myocardium dies in the first 2–4 days (1, 4, 33, 39). For example, 3 days after myoblast transplantation in muscles, Guerette et al. reported about 70% of cell death. This percentage continued to increase during the following 3 days (14).
One option to counteract the massive cell death and thus increase the transplantation efficiency is to increase proliferation in the muscle of the transplanted myoblasts before their fusion with the existing muscle fibers. Some articles pointed out that tyrphostin, also called AG490, a specific inhibitor of the janus tyrosine kinase JAK2, inhibits the fusion of myoblasts in culture (43, 44). JAK2 is one of the four members of the Janus kinase/signal transducer, which work in association with the activator of transcription (STAT) signaling pathway. JAK/STATs are involved in the regulation of myoblast differentiation. Sun et al. reported that JAK1-STAT1-STAT3 pathway not only is required for myoblast proliferation but also prevents myoblast premature differentiation. The shutdown of this pathway is a prerequisite for myoblasts to initiate their differentiation program (38). JAK1 and JAK2 have opposite roles. JAK2-STAT2-STAT3 positively regulates myogenic differentiation by regulating the expression of myogenic differentiation (MyoD), myocyte enhancer factor 2 (MEF2), and hepatocyte growth factor (HGF) (43). AG490 blocks myoblast fusion, so the proliferation of these cells should be increased.
We have thus investigated the effects of AG490 on the differentiation, proliferation, and survival of myoblasts in vitro and in vivo. Our work confirms that AG490 inhibits myoblast fusion in a dose-dependent manner. Moreover, we also observed that AG490 improved myoblast survival not only in culture after an apoptotic stress but more importantly after their intramuscular transplantation.
Materials and Methods
In Vitro Studies
Reagents
Fetal bovine serum (FBS), Trizol, trypsin, Dulbecco's modified Eagle's medium (DMEM), horse serum, penicillin/streptomycin, DNaseI, Superscript® reverse transcriptase, DTT, and CyQuant cell proliferation assay kit were obtained from Invitrogen (Burlington, Canada). Hank's balanced salt solution (HBSS), collagenase, phenol/chloroform/isoamyl alcohol, and Hoechst dye were purchased from Sigma Aldrich (Oakville, Ontario, Canada). Proteinase K was purchased from Novagen (Gibbstown, NJ, USA). The [methyl-14C]-thymidine was purchased from PerkinElmer (Boston, MA, USA). The Random primers, Go Taq, Oligo(dT) 15 primer, and RNasin® were obtained from Promega (Madison, WI, USA). Basic fibroblast growth factor (bFGF) was purchased from Gibco (Burlinton, Canada), and the Annexin V–fluorescein isothiocyanate (FITC) detection kit from PromoKine®, Bioscience Alive (San Jose, CA, USA).
Cell Culture
Myoblasts were grown from a muscle sample obtained from a 23-year-old man (a cadaveric organ donor) following informed consent of the family and approval by the Human Ethic Committee of the Centre Hospitalier de l'Université Laval's Research Center (CRCHUL). Human myoblasts were obtained after enzymatic digestion of the human muscle sample with collagenase (2%) in HBSS for 45 min at 37.5°C. The cells were then cultured at 37°C in a humidified atmosphere with 5% CO2 in proliferating medium (PM) MB1 (HyClone® MB-1 Medium, HyClone Laboratories, Inc., Logan, UT, USA) supplemented with 15% FBS, 10 ng/ml of bFGF, and 1% penicillin/streptomycin. Cells were used between passages 2 and 4 (12). The percentage of myoblasts in culture was determined by desmin immunostaining. At passage 3, more than 90% of the cells still expressed desmin.
Cell Differentiation Assay
Myoblast differentiation was determined by myotube formation, myotubes were defined as cells with three or more nuclei. The myoblasts were cultured, 30,000 cells in 24-well plate in the differentiation medium (DM) composed of DMEM containing only 2% horse serum for 4 days with or without AG490 (10-80 μM) or for 4 days longer without AG490. Myotubes were visualized by myosin heavy chain (MyHC) labeling. The cells were fixed with 95% ethanol and then washed twice with phosphate-buffered saline for 10 min and blocked with 10% FBS in phosphate-buffered saline. The samples were incubated for 2 h at room temperature with anti-MyHC antibody (MF20; DSHB, Department of Biological Sciences, Iowa City, IA, USA) (1:100) and for 1 h with anti-mouse Alexa 546-conjugated secondary antibody (red) or with anti-mouse Alexa 488-conjugated secondary antibody (green) diluted 1:100. Finally, the cells were washed with phosphate-buffered saline, and the nuclei were stained with Hoechst dye. The experiments were made three times in duplicate. Three to five pictures of each well were taken for the analysis (n ≥ 25). The number of nuclei incorporated in multinucleated myotubes was counted to estimate the terminal differentiation and normalized with the total number of nuclei. Results were expressed as a percentage of fusion.
Gene Expression
The total RNA was extracted using Trizol reagent from the myoblasts cultured in PM or DM with or without AG490. First-strand cDNAs were synthesized using 1 μg total RNA with the Oligo(dT) primer and Superscript III reverse transcriptase. Polymerase chain reaction (PCR) was performed with 1 μg of the cDNA solution using the primers specific for glyceraldehyde 3-phosphate dehydrogenase (GAPDH, as reference gene), MyoD, myogenin, Myf5 (myogenic factor 5), and MyHC. The annealing temperatures and the numbers of cycles of PCR are presented in Table 1. PCR products were then separated by electrophoresis in 1% agarose gels and were stained with ethidium bromide. A picture of the gel was obtained with the GelDoc program (BioRad) under UV light.
PCR Information on Genes Used in This Study

MyoD, myogenic differentiation; myf5, myogenic factor 5; myHC, myosin heavy chain; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Cell Proliferation Assay
The cell proliferation assay was performed using a CyQuant Cell Proliferation Kit, which measures the nucleic acid content in the test samples, according to the manufacturer's instructions. Human myoblasts were seeded in 24-well plates at initial densities of 30,000 cells per well and cultured in presence of AG490 or not (10–80 μM). The cells were harvested after various treatment times and stored at −80°C until the analysis. The frozen microplates were then thawed at room temperature, and the CyQUANT GR dye/cell lysis buffer was added. After incubating for 5 min, the fluorescence was measured (excitation/emission: 495/520 nm) using a microplate reader. The experiments were made three times in triplicate.
Cell Death Assay
The myoblasts were cultured on 24-well plates at 50,000 cells per well. The cells were pretreated or not with different concentrations of AG490 for 24 h. The toxicity of AG490 was estimated at this time. Different conditions were tested to simulate the cell conditions after transplantation: oxidative stress with H2O2 (10 mM, 2 h) and the anoikis (24 h). Anoikis was induced by culturing the cells over a 2% agarose gel to avoid the adhesion of the myoblasts to the culture surface (5, 27). After the induction of apoptosis, the cells were collected and centrifuged. The pellet was suspended in a buffer, and Annexin V-FITC and propidium iodide were added, according to the manufacturer protocol. Flow cytometry was used to determine the percentage of living cells and apoptotic cells. In Figure 1, we verified by fluorescence-activated cell sorting (FACS) that the treatment with H2O2 led to an apoptotic death of the cells. The cells without any staining for Annexin V and PI (Q3 percentage) were living cells. The Q1 represented only the cells in necrosis. The apoptotic cells were positive for Annexin V and were in Q4 (early apoptosis) and Q2 (late apoptosis and necrosis). As illustrated in Figure 1, the cell death was predominantly apoptotic. Results were expressed as percentage of living cells and compared to the control culture. The experiments were made three times in triplicate.

Flow cytometry was used to determine the percentage of living cells and apoptotic cells. The cells without labeling represented living (Q3) cells; the cells labeled with Annexin V-FITC (Q2, Q4) or propidium iodide (Q1, Q2) were respectively cells in apoptosis and in necrosis.
In Vivo Studies
Animals
The transplantations were done in 2-month-old Rag/mdx mice produced in our animal facilities by crossing mdx/mdx mice [a Duchenne muscular dystrophy (DMD) model due to a mutation leading to dystrophin deficiency] with Rag-/- mice (an immunodeficient mouse that accepts human grafts). Mice were maintained under pathogen-free conditions. All the experiments with mice were conducted after approval by the CRCHUL Animal Care Committee and in agreement with the guidelines of the Canadian Council for Animal Care.
Cell Transplantation Technique
Five hundred thousand human male myoblasts were transplanted in the tibialis anterior (TA) muscle. Briefly, the animals were anesthetized with isoflurane USP (Abbott Laboratories, Montreal, Canada). The skin was opened to expose the TA and myoblasts, resuspended in 10 μl of HBSS, were slowly injected obliquely using a glass micropipette (Drummond Scientific Co., Broomall, PA, USA) into 7–12 sites of the TA (3). The skin was closed with absorbable sutures.
Cell Death Assay at 5 Days
To quantify the in vivo cell mortality, myoblasts were radiolabeled by culturing them 48 h in growth medium containing 0.25 μCi/ml [methyl-14C]thymidine (50 mCi/mmol) before the transplantation. Radiolabeled cells (500,000 cells) were injected throughout the TA using a glass micropipette as previously described. The muscles were removed at days 0 and 5 after cell transplantation, snap-frozen in liquid nitrogen, and stored at −80°C. DNA was extracted from transplanted TAs. Briefly, muscles were minced and incubated with 50 μl of proteinase K (10 mg/ml) at 56°C until the solution became clear. Digested muscles were then mixed with 500 μl of a solution of phenol/chloroform/isoamyl alcohol (25:24:1) and centrifuged 3 min at 13,000 rpm. The upper solution was recovered and mixed with the same volume of chloroform and centrifuged again. The upper solution was recovered, and 25 μl of 3 M citrate sodium was added before the addition of 1 ml of 100% ethanol. After centrifugation 8 min at 13,000 rpm, the pellets were washed in 70% alcohol before another centrifugation. The pellets were then dried before the suspension of the DNA in 100 μl sterile water. The amount of radiolabel within each TA was measured on DNA extracts using liquid scintillation counter (Mod. Wallac 1409, Woodbridge, Ontario, Canada). The amount of residual radioactivity at day 5 was expressed as a percentage compared to day 0.
Experimental Groups
In the first experiment, the cultured myoblasts were treated 24 h before the transplantation with AG490 (20-80 μM). The cells treated and not treated were resuspended in HBSS as previously described for the transplantation.
In the second experiment, the cells were cultured without AG490 but suspended in HBSS or in AG490 solutions (4 μM to 4 mM) before the transplantation.
In the last experiment, the myoblasts were not treated. The mice received AG490 [5 mg/kg/day, based on a study from El-Adawi et al. (8)] or a saline solution intraperitoneally (IP) 1 day before cell transplantation and for the 5 days thereafter. Each experiment was made twice with at least four muscles per group (n ≥ 8).
Statistical Analysis
Statistical analysis was performed by Kruskal–Wallis chi-squared. Differences were considered statistically significant at p ≤0.05 (*) and p ≤0.01 (**). The experimental values were presented as means ± SD. The number of points or animals used for each study is indicated in the figure legends.
Results
In Vitro Studies
AG490 Blocks Myoblast Fusion
Myoblasts were cultured 4 days in differentiation medium (DM) with or without AG490 (10–80 μM). After 4 days, the cells were fixed and stained for myosin heavy chain (MyHC) in Figure 2A. The number of nuclei incorporated in MyHC-positive fibers was expressed as a percentage compared to the total nuclei (percentage of fusion, Fig. 2B). After 4 days in differentiation medium, the fusion of control cells reached roughly 50%. This percentage decreased when the cells were cultured in presence of AG490 and became significantly different with concentrations higher than 10 μM of AG490. Another experiment consisted in culturing the cells with AG490 during 4 days as previously described and then to replace the medium with the differentiation medium without any AG490 for another 4 days. MyHC-positive fibers were then stained to visualize the step of differentiation. The pictures in Figure 2C showed that the differentiation process was still occurring at 8 days, even after a treatment with 40, 60, and 80 μM AG490 for 4 days.
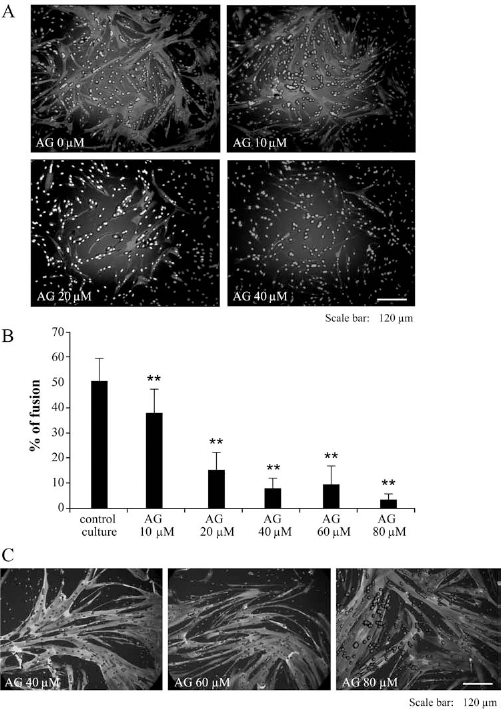
AG490 (AG) blocks the fusion of myoblasts. Myoblasts were cultured in differentiation medium (DM) for 4 days in presence of AG (10–80 μM) or not. The percentage of fusion (B) was estimated in each condition from a MHC staining (A). The presence of AG during the culture diminished the percentage of fusion in a dose-dependent manner (*significantly different from the control DM; *p ≤0.05, **p ≤0.01; n ≥ 25). In (C), the cells were cultured as previously described for 4 days and then cultured for another 4 days only with the differentiation medium. When the myoblasts were cultured in the differentiation medium, even after a treatment with AG490 at 40–80 μM, the differentiation process occurred, which showed a reversible inhibitory effect of AG490 on differentiation.
RNA extraction was performed on the cells cultured or not with AG490 in the DM, followed by an RT-PCR for different myogenic genes. GAPDH was used as a reference gene in Figure 3A. In the differentiation medium (DM condition), the expression of MyHC was stronger than the expression of MyoD or myogenin. Myogenin and MyoD expression was a little decreased in presence of high concentration of AG490. However, the expression of MyHC was the most affected by the presence of AG490. MyHC is expressed later during the differentiation progress compared to MyoD and myogenin, which suggested that the terminal differentiation of myoblasts was inhibited by AG490.

Effect of AG490 (AG) on gene expression and proliferation. Myoblasts were cultured in differentiation medium (DM) for 4 days in presence of AG or not. The expression of genes involved during the differentiation process was analyzed (A). The expression of MHC and in a lesser extends of MyoD and myogenin were decreased in presence of AG especially with 80 μM. The proliferation was estimated after 4 days of culture with or without AG (B) (*significantly different from the day 0; †significantly different from the control medium at day 4; *p ≤ 0.05, **p ≤ 0.01; the experience was made three times in triplicate).
The proliferation of the cells, in Figure 3B, was also estimated in DM culture by measuring the OD with the CyQuant cell proliferation assay kit. After 4 days in DM, the optical densities (OD) of the cells cultured without AG490 and with 10 and 20 μM AG490 were significantly higher than the control at day 0 indicating that they have proliferated. However, the OD of cells cultured with 40–80 μM AG490 were not significantly different from the OD of control cells at day 0 but also not significant different from the OD of cells cultured without AG490 during 4 days for the conditions of 40 and 60 μM. Only the concentration of 80 μM AG490 gave an OD significantly lower than the cells cultured without AG490 during 4 days in DM. Thus the presence of AG490 affected the cell fusion from 10 μM and the proliferation at 80 μM.
AG490 Acts Reversibly on Myoblast Proliferation
The myoblasts were cultured 4 days in proliferation medium (PM) with or without AG490 (Fig. 4A). At day 4, the cells cultured without AG490 and with 10, 20, and 40 μM of AG490 had an OD significantly higher than the OD at day 0. The OD values of cells cultured in presence of 60 and 80 μM were not significantly different from the control at day 0 but significantly lower than the OD value from cells cultured without AG490 at day 4.

AG490 (AG) acts reversibly on the proliferation of myoblasts. The proliferation of the myoblasts was estimated after 4 days in a proliferation medium (PM) in presence of AG or not (A) (*significantly different from the day 0; †significantly different from the control medium PM at day 4; *p ≤0.05, **p ≤0.01; n = 9). The expression of genes involved in the differentiation process were also analyzed (B). In (C), the proliferation was tested after a pretreatment of 24 h with AG (D-1 to D0) or not (PM condition) and after a culture in a PM during two other days (D2) (*p ≤0.05, **p ≤0.01; the experiment was made three times in triplicate).
Expression of various genes was also investigated in these cells at day 4 in Figure 4B. The expression of genes characteristic of differentiation (MHC, MyoD, and myogenin) was reduced in presence of AG490, especially with the higher concentrations. The expression of these genes was also lower compared to the cells cultured in DM in Figure 2B. Myf 5 gene was highly expressed compared to the other genes investigated.
Myoblasts were also cultured 24 h with or without AG490 (D-1 to D0) and then cultured for 2 days without AG490 (Fig. 4C). The proliferation was measured after cell plating (D-1), after 24 h with or without AG490 (D0), and after 2 days in PM without AG490 (D2). The OD value after 24 h in presence of 80 μM AG490 was significantly lower than the OD from the cells cultured without AG490. After 2 days in PM, only the cells pretreated with AG490 at 60 and 80 μM were not as high as the cells cultured without AG490. Nevertheless, the OD values at day 2 were all significantly different from the day -1, indicating that the cells had proliferated.
AG490 Protects Myoblasts Against Apoptosis
First of all, the myoblasts were cultured with or without AG490 and analyzed to estimate the toxicity of this product. In Figure 5A, the percentage of living cells was between 75% and 85% for all the conditions of culture. Myoblasts were then cultured with or without a 24 h pretreatment with AG490 and then exposed to H2O2 for 2 h to mimic the oxidative stress after cell transplantation. The results were expressed as percentage of living cells (Fig. 5B). When the cells were exposed to H2O2 without any pretreatment with AG490, the survival rate decreased from 73% (percentage of living cells in the culture) to 45% of living cells. A pretreatment with AG490 at 20–80 μM provided a significant increase of the survival compared to the cells without pretreatment. The percentages of survival obtained with an AG490 pretreatment were not significantly different from the survival of the control cells.
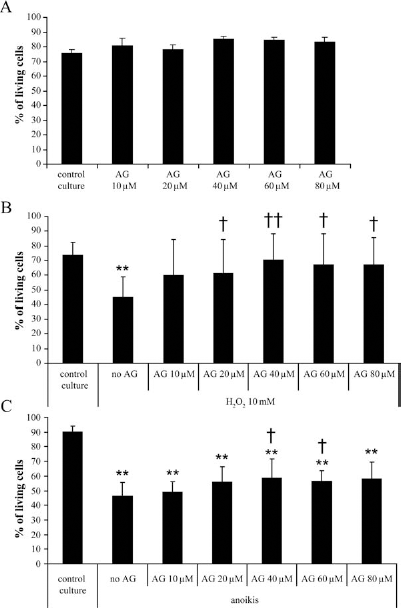
AG490 (AG) protects the myoblasts against apoptosis. The cells were pretreated or not with different concentrations of AG and analyzed by FACS using Annexin V (FITC-A axis) and PI staining (PI-A axis). The percentage of living cells was shown in (A). The myoblasts were also pretreated or not with AG490, as previously described, and then exposed to H2O2 for 2 h. The treatment with H2O2 induced apoptosis, as indicated by the presence of Annexin V-positive cells. The percentage of living cells in the different AG concentrations after the treatment with H2O2 is represented in (B). Some myoblasts were also plated over an agarose gel to induce anoikis. The percentage of living cells is represented in (C) in function of AG concentrations (*significantly different from the PM; †significantly different from the H2O2 treatment or anoikis without AG; *p ≤0.05, **p ≤0.01; the experiment was made three times in triplicate).
The cells were also cultured with or without AG490 24 h and then cultured over a 2% agarose gel to induce cell death through a form of apoptosis, called anoikis (5, 27). The percentage of apoptotic cells was also determined by FACS, and the results were expressed as percentage of living cells (Fig. 5C). When the cells were not pretreated with AG490, the survival rate fell from 90% to 46% in anoikis conditions. The percentage of living cells increased significantly to 59% and 56% with pretreatment with 40 and 60 μM AG490, respectively. However, the percentages of living cells obtained with a pretreatment with AG490 never reached the percentage of living cells in a culture without anoikis.
In Vivo Studies
Coinjection of AG490 Ameliorates the Survival of Transplanted Myoblasts
Different treatments were tested to establish which one best improved the survival of myoblasts transplanted in a muscle. First, the cells were pretreated with AG490 during 24 h before the transplantation. In a second experiment, the cells were cultured without AG490, but the cells were resuspended in a solution containing AG490 at the time of transplantation. Finally, based on an article from El-Adawi et al., the mice were treated or not with AG490 by daily IP injections (8).
The myoblasts were radiolabeled and cultured 24 h with (0, 20, 40, 60, and 80 μM AG490) before their transplantation in the TA of RAG/mdx mice. After 5 days, the muscles were taken, and the residual radioactivity was measured. When the myoblasts were pretreated with AG490 (20–80 μM), they did not survive better than the nontreated cells (Fig. 6A). Moreover, when the cells were pretreated 24 h with the highest concentration of AG490 (80 μM), the survival was significantly lower than in the control without AG490.

Coinjection of AG490 (AG) ameliorates the survival of transplanted myoblasts. The survival of transplanted myoblasts was evaluated by measuring the residual radioactivity at day 5 compared to day 0. In the first experiment, the myoblasts were radiolabeled and cultured 24 h with or without AG before the transplantation (A; n ≤; 9). In the second experiment, the cells were suspended in AG solutions and immediately transplanted in the muscles of mice (B; n ≤; 8). Note that, in the last experiment, only the mice were treated or not with AG and not the cells. One group of mice was injected IP with saline (HBSS) and the other group with AG (5 mg/kg/day) (C; n = 8). The muscles were recovered at day 0 and day 5. The residual radioactivity was expressed as a percentage of day 0 value (*significantly different from the control HBSS; *p ≤ 0.05; **p ≤ 0.01).
The second experiment was to deliver the AG490 in the same time as the myoblasts during the graft. The cells were suspended in HBSS containing AG490 or not (4 μM, 40 μM, 400 μM, or 4 mM) and then transplanted into the muscle. In this case, the three highest concentrations gave a significantly higher survival of the transplanted cells (Fig. 6B). The residual radioactivity (indicating the percentage of survival) increased from 5.5% to 8.0%, 7.3%, and 7.8%, respectively, which correspond to 45%, 33%, and 41% of increase of survival compared to the control.
Based on a study on postinfarction remodeling (8), we also treated the mice with AG490 1 day before the graft and daily until their sacrifice at day 5 after the transplantation. However, the residual radio activity obtained after 5 days was similar when the cells were transplanted in mice or in mice treated with AG490 (Fig. 6C). In both cases, the percentage of survival stayed around 5.7% and 6.2%, respectively, for mice treated with HBSS or AG490.
Discussion
AG490 and Myoblast Differentiation
The expression of different genes was analyzed after 4 days of culture in proliferation medium or in differentiation medium. In the proliferation state, myoblasts expressed Myf5 at a higher degree than the other genes. On the contrary, when the cells were induced to differentiate, stronger expressions were observed for myogenin and MyHC. All these observations were in accordance with the literature. During the proliferation of myoblasts, MyoD and Myf5 were expressed, then during differentiation Myf5 expression was replaced by myogenin expression. Lastly, muscle-specific genes such as MyHC were expressed (7, 24).
Wang et al. previously described that concentrations of 20 and 40 μM AG490 inhibited the fusion of C2C12 and of mouse primary culture myoblasts in a dose-dependent manner (43, 44). They also confirmed their results by RT-PCR with a decrease in MyHC and myogenin expression in cells treated with AG490. In our study, we also found that AG490 reduced the fusion of human myoblasts at concentrations above 10 μM. Furthermore, the expression of myogenic genes was also altered by the treatment with AG490. Yang et al. used the C2C12 cell line to define the implication of STAT3 during the differentiation process (44). When the cells were shifted from growth medium to differentiation medium, the expression and the phosphorylation of STAT3 were increased. During muscle regeneration, growth factors, such as leukemia inhibitory factor (LIF), are acting with other transcription factors to regulate the proliferation and the differentiation of myoblasts. Yang et al. showed that myogenic differentiation induced by LIF was mediated through JAK2-STAT3 pathway (44). These results could explain how AG490 acted on myoblast differentiation by inhibiting this pathway.
AG490 and Proliferation
The proliferation of myoblasts was evaluated in differentiation medium (DM) and in proliferation medium (PM). In both cases, we observed that the proliferation of cells was decreased starting at a concentration of 40 μM AG490 This inhibition of proliferation was also observed in Jurkat T-cells to be dose dependent. The authors attributed the result to an arrest of the cell cycle rather than to apoptosis (10, 25).
LIF was also used by Spangenburg et al. to enhance the proliferation of C2C12 and rat primary myoblasts (37). With 10 μM AG490 in the medium, the proliferation was drastically decreased after 48 h. These researchers also found that LIF increased the proliferation of the cells in particular through JAK2-STAT3 pathway.
These two previous articles using LIF (37, 44) showed that LIF enhanced either myoblast proliferation or the differentiation in vitro, depending of the medium culture (proliferation medium or differentiation medium, respectively). Moreover, LIF acts through JAK2/STAT3 pathway. Thus, AG490, as a specific inhibitor of JAK2, inhibits this pathway, affecting the proliferation or the differentiation, depending on the situation.
AG490 and Apoptosis
In our experiments, two different methods were used to induce apoptosis. The first one was by treating the cells with H2O2, which is classically used in cell culture (9, 13, 16, 19, 21), at a concentration less than 10 mM because higher concentration leads to necrosis instead of apoptosis (11). The death was confirmed to be due to apoptosis by Annexin V staining. In the Jiung et al. study, apoptosis induced by H2O2 could be reduced or eliminated by a heat shock treatment of cardiomyocytes and C2C12 cells (16). Our study also showed that a 24-h pretreatment with AG490 (20–80 μM) completely prevented the apoptosis of cells induced by H2O2. This protection against oxidative stress-induced apoptosis was also observed in lens epithelial cells with 40 μM AG490 (19), in astrocytes with 30 μM AG490 (13), and in vascular smooth muscle cells (VSMCs) with 100 μM AG490 (32). In this last article, the authors showed that hydrogen peroxide-induced apoptosis was mediated through JAK2 tyrosine kinase activation. This pathway was blocked using AG490, leading to the survival of the VSMCs. Another article showed that AG490 could also prevent apoptosis though the activation of Akt and ERK signaling pathway in osteclast (21) or the protection of neurons against a mitochondrial toxin (18).
Preventing the attachment of myoblasts to the plastic of the culture flask using an agarose gel induced another form of apoptosis called anoikis. Bouchentouf et al. showed that the percentage of apoptotic cells induced by this method in 13 months old human myoblasts reached less than 20% after 72 h (5). In our experiment, we found a percentage of apoptosis of about 40% after 48 h in anoikis conditions. The differences observed between these two studies in term of cell survival could be explained by the fact that the cells came from two different biopsies. The age of the patients at the time of the biopsy is of great importance on the behavior of the cells in culture and after transplantation.
Bouchentouf et al. also showed that the induction of anoikis requires the Bit1 and FADD pathways (5). Using AG490 in our experiment helped the cells to survive to anoikis condition, suggesting that JAK2 pathway will also play a role during anoikis. This supposition was confirmed in endothelial cells, where the inhibition of JAK2 with AG490 strongly protected these cells against cell detachment-dependent death (anoikis) (30).
AG490 and Cell Survival
Last but not least, the survival of transplanted myoblasts was evaluated in our study with transplanted myoblasts in the TA of mice. Myoblast transplantation was also studied by other groups. Suzuki et al. transplanted 500,000 myoblasts in intact hearts of mice. They found only 7.4% of survival 72 h after this transplantation. However, if the myoblasts were transplanted in a solution containing CuZn-SOD, the percentage of cell survival was increased by twofolds (39). Haider et al. also observed a twofold increase when the myoblasts were pretreated with diazoxide before the transplantation in an acute myocardial infarction (15). Unfortunately, no difference in cell survival was obtained in our study when the cells were pretreated with AG490 during 24 h before transplantation.
Some authors tried to increase the survival of the transplanted myoblats through genetic modifications. For example, Merly et al. and Bouchentouf et al. overexpressed VEGF and TGF-β. In both cases, the survival of the transplanted cells increased by 40% at day 4 with VEGF and by 86% at day 3 with TGF-β (4, 28). Other studies tried to improve the survival of the transplanted myoblasts without genetic modification by using growth factors or a biomaterial. Lafreniere et al. transplanted the myoblasts in a saline solution containing 50 ng of bFGF or IGF1 or not (22). The authors evaluated the residual radioactivity 2 days later and found no difference in cell survival between the different treatments. However, the use of a biomaterial-like fibrin gel induced a 46% increase of the survival of myoblasts 5 days after transplantation (12). This result is similar to the one obtained in this study with a 45% increase at day 5 of residual radioactivity when the cells were coinjected with 40 μM AG490. As mentioned in the previous study, it is very difficult to compare the results obtained because of the many experimental differences between studies. For instance, the types of cells transplanted, the transplantation technique, and the degree of muscle injury are all factors that differ from one study to another (12).
Another approach to increase the survival of transplanted cells is to treat the mice rather than the cells. Indeed Fakhfakh et al. used losartan to block the TGF-β activity and enhance the proliferation of the myoblasts (9). Improved in vivo cell survival was also observed in that article by treating the mice instead of the cells. The authors found a 67% increase of survival 72 h after the transplantation when the mice were treated everyday with losartan compared to nontreated mice (9). In our study, some mice were given a daily injection of AG490 1 day before and until the 5th day after transplantation. Unfortunately, no difference in cell survival was observed after transplantation compared to the nontreated mice. However, El-Adawi et al. studied post-myocardial infarction (MI) and showed that daily injection of AG490 in rat enhanced apoptosis in the border zone of the heart 1 day post-MI (8). On the other hand, Mascareno et al. showed cytoprotective effects of STAT2 inhibition by AG490 in an ischemia–reperfusion rat model (26). Moreover, Beckles et al. revealed that a treatment with AG490 blocked the formation of cardiac hypertrophy after a transverse aortic constriction through inhibition of JAK2 and STAT3 (2). Thus, the role of the JAK-STAT pathway seems to be very different from a protocol to another.
Our article describes the AG490 effects on myoblasts. In vitro, when myoblasts were cultured during 24 h, their proliferation was inhibited with AG490 concentrations above 40 μM. Cell proliferation was restored when AG490 was removed. This was also the case for the differentiation process indicating the reversible effects of AG490. Moreover, AG490 strongly protected myoblasts against apoptosis. In vivo, the coinjection of AG490 with the transplanted cells was sufficient to increase the survival rate after 5 days. All these results together underline the fact that AG490 should be present to have a real action on the cells. This would also explain why no effect was observed in vivo with a pretreatment of the cells. Because of the short half-life and the low solubility of AG490, it should be given daily by a subcutaneous osmotic pump or by intraperitoneal injections. Two studies used daily injection of AG490 ranging from 300 to 800 μg per mouse (29, 45). In our experiments, we used 5 mg/kg/day, which is only about 175 μg of AG490 per mouse daily. Probably our dose was not high enough to protect the transplanted cells.
Conclusion
In spite of all the efforts put on the amelioration of the cell survival after transplantation these last years, no one had yet found a real solution. In this article, the in vivo survival of myoblasts was increased by 45% at 5 days after transplantation by simply coinjecting the cells with a 40-μM AG490 solution. This result is interesting but not sufficient for a clinical assay. Because the poor success of cell-based therapies is due to many factors, we probably have to find a complex treatment (cocktail of components) to enhance simultaneously not only myoblast survival but also their migration inside the muscle and their fusion with the existing muscle fibers. AG490 should be a part of such a cocktail by acting on cell survival in the first hours or days of transplantation before the natural elimination of AG490 by the body, permitting the migration and fusion of the transplanted myoblasts.
Footnotes
Acknowledgments
This work was supported by grants from the Jesse's Journey Foundation for Gene and Cell Therapy of Canada and the Association Française contre les Myopathies (AFM). Dr. Jacques P. Tremblay has shares in CellGene, Inc., a biotechnological company created to accelerate the development of cell therapies. The authors declare no conflicts of interest.