
Abstract
For clinical islet transplantation, isolated islets deteriorate rapidly in culture, although culturing islets prior to transplantation provides flexibility for evaluation of isolated islets and pretreatment of patients. In the present study, we compared human fresh islets to cultured islets with in vitro and in vivo assays. After culture for 24, 48, and 72 h, islet yield significantly decreased from 2,000 to 1,738 ± 26 (13% loss), 1,525 ± 30 (24% loss), or 1,298 ± 18 IEQ (35% loss), respectively. The ATP contents were significantly higher in the 6-h cultured group (near fresh group) than in 48-h culture groups. The stimulation index was relatively higher in the 6-h cultured group than in 48-h cultured group. Human islets with or without culture were transplanted into diabetic nude mice. The attainability of posttransplantation normoglycemia was significantly higher in fresh group than in the culture groups. Intraperitoneal glucose tolerance testing (IPGTT) showed that the blood glucose levels of mice transplanted with fresh islets were significantly lower than with cultured islets at 30, 60, 90, and 120 min after injection. These data suggest that human islet transplantation without culture could avoid the deterioration of islets during culture and improve the outcome of islet transplantation. Based on these data, we have transplanted fresh islets without culture for our current clinical islet transplantation protocol.
Introduction
Transplantation of islets of Langerhans can improve glycometabolic control in type 1 diabetic patients (38, 39). However, each recipient usually requires islets from two or three donor pancreata. Current isolation techniques usually recover fewer than half of the islets from a given pancreas (19,30,38). It is generally assumed that considerable islet loss occurs due to ischemia during the immediate transplantation period combined with immunological destruction.
Recently, many centers have introduced the culturing of human islets prior to transplantation (7,8,12,13,37) because it provides many benefits to clinical islet transplantation. In vitro culture may reduce islet immunogenicity by depletion of viable hematogenous and lymphoid cells (11) and reduce exocrine contamination of transplanted tissue. Other benefits are additional quality control testing of isolated islets, initiating time-dependent immunosuppressive protocols, and it preserves the islets during travel time for recipients living far away from transplant centers. However, some groups have shown that freshly isolated islets are superior to cultured islets for islet transplantation (2,14,31).
We previously reported that clinical outcome in patients who received cultured islets was significantly lower than in patients who received fresh islets, although the number of subjects evaluated was not sufficient to conclude that fresh islets are better than cultured islets (29). It is well documented that isolated islets deteriorate rapidly in culture for clinical islet transplantation (9,12,13,29). Since barely half of the processed pancreata meet the criteria for clinical transplantation in most centers, islet loss during culture results in an even lower transplant rate.
In the present study, we compared islet transplantation outcomes of fresh islets to cultured islets.
Materials and Methods
Human Islet Isolation
The islet isolation protocol was approved by the Institutional Review Board of Baylor Health Care System. Thirty-four pancreata from brain-dead donors, which were procured from either Southwest Transplant Alliance (Dallas, TX) or LifeGift (Fort Worth, TX) between May 2007 and April 2009, were used in this study. All pancreata were procured using a standardized technique to minimize warm ischemia. University of Wisconsin (UW) solution was used for in situ perfusion of the donor. The pancreas was excised immediately after the liver and before the kidneys and was placed on ice. After the removal of the spleen and duodenum, we immediately inserted a cannula into the main pancreatic duct. The pancreas was weighed and 1 ml/g pancreas weight of modified Kyoto solution (Kyoto solution with ulinastatin, Kyoto solution, Otsuka Pharmaceutical Factory, Inc., Naruto, Japan; ulinastatin, Mochida Pharmaceutical, Tokyo, Japan) was infused through the intraductal cannula (16,27). Pancreata were placed into a modified Kyoto solution/perfluorochemical two-layer preservation container at 4°C (25,26,28) for less than 8 h until the islet isolation procedure.
Human islet isolation was conducted as previously described (16,17,21,22) in the standard Ricordi technique (35) with modifications introduced in the Edmonton protocol (18,33,38). In brief, after decontamination of the pancreas, the ducts were perfused in a controlled fashion with a cold enzyme blend of Liberase HI (1.4 mg/ml; Roche Molecular Biochemicals, Indianapolis, IN) or Serva collagenase NB1 (20 PZ U/ml) with neutral protease (1 DMC U/ml) (SERVA Electrophoresis GmbH, Heidelberg, Germany). The distended pancreas was then cut into 7–9 pieces, placed in a Ricordi chamber, and shaken gently. While the pancreas was being digested by recirculating the enzyme solution through the Ricordi chamber at 37°C, we monitored the extent of digestion with dithizone staining by taking small samples from the system. Once digestion was confirmed to be complete, dilution solution (Mediatech, Inc., Manassas, VA) was introduced into the system. Then the system was cooled to stop further enzymatic activity. The digested tissue was collected in flasks containing 25% HSA and washed with fresh medium to remove the enzyme. Islets were purified with a continuous density gradient of iodixanol-based solution (Optiprep®, Sigma-Aldrich, St. Louis, MO) in an apheresis system (COBE 2991 Cell Processor, Gambro Laboratories, Denver, CO) as previously reported (16,20,22).
Islet Count and Evaluation
The crude number of islets in each diameter class was determined by counting islets after dithizone staining (3 mg/ml final concentration, Sigma Chemical Co., St. Louis, MO) using an optical graticule. The crude number of islets was then converted to the standard number of islet equivalents (IEQ; diameter standardizing to 150 μm) (34).
Islet function was assessed by monitoring the insulin secretory response of the purified islets during glucose stimulation using a procedure described by Shapiro and colleagues (38,39). Briefly, 1,200 IEQ were incubated with either 2.8 or 25 mM glucose in RPMI-1640 for 2 h at 37°C and 5% CO2. The supernatants were collected and insulin levels were determined using a commercially available enzyme-linked immunosorbent assay (ELISA) kit (ALPCO Insulin ELISA kit; ALPCO Diagnostics, Windham, NH). The stimulation index was calculated by determining the ratio of insulin released from islets in high glucose to the insulin released in low glucose. The data were normalized to total DNA. The data were expressed as mean ± SE.
Islet Culture
Isolated islets from human pancreata were cultured for 6, 24, 48, and 72 h at 37°C and 5% CO2 in culture medium. Culture medium was CMRL-based Miami-defined media #1 (MM1; Mediatech-Cellgro, Herndon, VA) containing 0.5% human serum albumin, which has been used for human islet transplantation previously (9). Islets were seeded as 100–150 IEQ/cm2 dish area and the height of the media was 1.5–2.0 mm; these conditions are similar to culture conditions for clinical islet transplantation (20,000–30,000 IEQ per T-175 flask, 30–50 ml per T-175 flask).
Determination of ATP Production
To measure adenosine triphosphate (ATP) production, human islets were cultured for 6 or 48 h at 37°C and 5% CO2 in culture medium. They were then washed twice with ice-cold PBS and solubilized. The amount of ATP was measured using an ATP assay system (ATP-lite, Perkin Elmer, Groningen, Netherlands), according to the manufacturer's instructions. The data were normalized to total DNA. The data were expressed as mean ± SE.
In Vivo Assessment
Nude mice were rendered diabetic by a single IP injection of streptozotocin (STZ) at a dose of 220 mg/ kg. Hyperglycemia was defined as a glucose level of >350 mg/dl detected twice consecutively after STZ injection. Immediately after isolation, 2,000 IEQ of human islets were incubated at 37°C and 5% CO2 in culture medium for 48 h. The human islets with or without culture were transplanted into the renal subcapsular space of the left kidney of a diabetic nude mouse, as previously described (24). During the 30-day posttransplantation period, the nonfasting blood glucose levels were monitored three times per week. Normoglycemia was defined when two consecutive blood glucose level measurements showed less than 200 mg/dl. The intraperitoneal glucose tolerance testing (IPGTT) was performed 30 days after transplantation. The mice were fasted overnight, after which glucose (2.0 g/kg body weight) was injected intraperitoneally. The blood glucose levels were measured before injection and at 5, 30, 60, 90, and 120 min after injection. No statistical differences in either pretransplantation blood glucose levels or pretransplantation body weight were observed between two groups of mice. Mouse studies were approved by the Institutional Animal Care and Use Committees (IACUC).
Statistics
Two groups were compared by Student's t-test or the Kaplan-Meier log-rank test. The differences between each group were considered significant if the value was p < 0.05.
Results
Islet Equivalents After Culture
To evaluate islet equivalents after culture, 2,000 IEQ of isolated islets from human pancreata were cultured for 24, 48, and 72 h at 37°C. After culture for 24, 48, and 72 h, islet yield significantly decreased from 2,000 to 1,738 ± 26 (13% loss), 1,525 ± 30 (24% loss), or 1,298 ± 18 IEQ (35% loss), respectively (Fig. 1). These data suggest that isolated islets deteriorate time dependently in culture.

Number of islets after culture. Immediately after each isolation, 2,000 islet equivalents (IEQ) of islets were cultured for 72 h at 37°C and 5% CO2 in culture medium. After 24, 48, or 72 h of culture, the islets were counted to calculate IEQ in each group (n = 5).
In Vitro Assessment
To assess the islet quality in each group in vitro, human islets were evaluated by ATP contents and stimulation index. Since islet isolations are performed mainly at 4°C, islets immediately after isolation have lower activity than islets in 37°C culture, resulting in lower ATP contents and stimulation index. Therefore, we compared short-time cultured islets (6 h) with 48-h cultured islets. The ATP contents were significantly higher in the 6-h cultured group (near fresh group) than in 48-h culture groups (6 h, 126.6 ± 5.7 pmol/μg; 48 h, 101.5 ± 7.5 pmol/μg) (Fig. 2). The stimulation index was relatively higher (but not significant) in the 6-h cultured group (near fresh group) than in 48-h culture groups (6 h, 6.21 ± 0.80; 48 h, 2.64 ± 0.25) (Fig. 3). These data suggest that the quality of fresh islets may be higher than cultured islets.
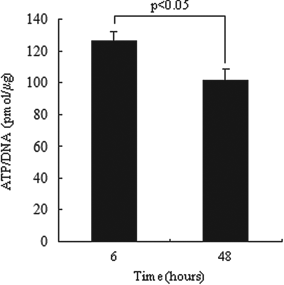
ATP content of human islets. After 6- or 48-h culture at 37°C and 5% CO2, the adenosine triphosphate (ATP) concentration of the cell lysate was measured using an ATP assay system. The ATP was normalized to DNA concentration (each group, n = 5). Data are expressed as the mean ± SE.

Stimulation index of human islets. After 6- or 48-h culture at 37°C and 5% CO2, islet function was assessed by monitoring the insulin secretory response of the purified islets during glucose stimulation. One thousand two hundred IEQ were incubated with either 2.8 or 25 mM glucose in RPMI-1640 for 2 h at 37°C and 5% CO2. The supernatants were collected and insulin levels were determined using a commercially available ELISA kit. The stimulation index was calculated by determining the ratio of insulin released from islets in high glucose to the insulin released in low glucose. The data were normalized to total DNA (each group, n = 6). The data were expressed as mean ± SE.
In Vivo Assessment
To evaluate the human islets after culture in vivo, 2,000 IEQ of human islets were incubated at 37°C in culture medium. After 48 h of culture, all of the human islets were transplanted into the renal subcapsular space of the left kidney of STZ-induced diabetic nude mice. This model is similar to the situation of clinical islet transplantation (all of isolated islets are cultured for 24–48 h, and all of the cultured islets are transplanted into the patients) and can evaluate islet quality and quantity. To evaluate the fresh islets, 2,000 IEQ of human islets without culture were transplanted. The blood glucose levels of 4 of the 26 mice (15.4%) receiving islets after 37°C culture decreased gradually and reached normoglycemia. The blood glucose levels of 23 of the 28 mice (82.1%) receiving islets without culture reached normoglycemia. The blood glucose levels remained stable thereafter and returned to pretransplantation levels after kidney-bearing islets were removed (30 days after transplantation) (Fig. 4). The attainability of posttransplantation normoglycemia was significantly higher in fresh groups than in the 37°C culture group. These data suggest that fresh islet transplantation is superior to cultured islet transplantation.

Evaluation of quality and quantity of isolated islets with or without culture in vivo. Immediately after isolation, 2,000 IEQ islets were cultured for 48 h at 37°C. The islets with or without culture were transplanted below the kidney capsule of diabetic nude mice. Normoglycemia was defined as two consecutive posttransplant blood glucose levels showing less than 200 mg/dl. Fresh (n = 28, closed circles); culture (n = 26, open circles).
Intraperitoneal Glucose Tolerance Testing (IPGTT)
The IPGTT was carried out on both groups 30 days after transplantation. The mice in both groups that were hyperglycemic were excluded. The mice were fasted overnight, after which glucose (2.0 g/kg body weight) was injected intraperitoneally. The blood glucose levels were measured before injection and at 5, 30, 60, 90, and 120 min after injection. The blood glucose levels of mice transplanted with fresh islets were significantly lower than with cultured islets at 30, 60, 90, and 120 min after injection (30, 90, and 120 min, p < 0.01; 60 min, p < 0.05) (Fig. 5). These data suggest that the engrafted islet number may be higher in fresh group than in cultured group.
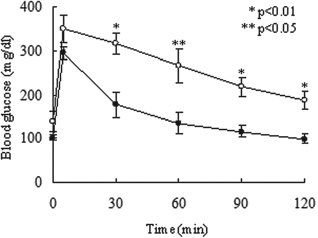
IPGTT in mice with fresh or cultured islets. Intraperitoneal glucose tolerance testing (IPGTT) was performed 30 days after transplantation. The mice were fasted overnight, after which glucose (2.0 g/kg body weight) was injected intraperitoneally. The blood glucose levels were measured before injection and at 5, 30, 60, 90, and 120 min after injection. No statistical differences in either pretransplantation blood glucose levels or pretransplantation body weight were observed between the two groups of mice. Fresh (n = 4, closed circles); culture (n = 4, open circles).
In summary, human islet transplantation without culture could avoid the deterioration of islets during culture and improve the outcome of islet transplantation.
Discussion
Our study shows that transplantation of human islets without culture is more effective than 37°C culture, which some groups have used. It is well known that there is a loss of islets, to varying degrees, during 22°C and 37°C cultures, even in culture conditions optimized for cell recovery (3,9,12,13,40). It has been reported that, using clinical-grade pancreata, islet yield after a 22°C culture for 20 h (median) significantly decreased preculture yield (15% loss) (13), as did a 37°C culture for 32.5 h (median) (20% loss) (9). Our data in this study showed a 24% loss in the 37°C culture after 48 h. The mechanisms for the reduction in islet mass during culture involves cell death by apoptosis and the prerecruitment of intracellular death signaling pathways, such as the activation of c-Jun NH2-terminal kinase (JNK) (1,2,5,6,10,23). One of the disadvantages for islet culture is the deterioration of islet number during culture.
King et al. showed that 89% of mice transplanted with 150 syngeneic islets without culture were cured and that only 22% of mice transplanted with same number of islets with culture were cured (14). They also showed that the fresh islets did have increased DNA content compared with cultured islets, suggesting that fresh islets have greater viable β-cell mass than cultured islets. Another possibility of increased success of the transplantation of fresh islets may be the intactness of their capillary network. The potential importance of donor endothelium in the revascularization process has recently been shown (4). Indeed, it has been shown that grafts from fresh islets have higher oxygen contents than those from cultured islets, even 1 month after transplantation (31).
On the other hand, advantages for culture are to test additional quality control of isolated islets, to initiate time-dependent immunosuppressive protocols, and to preserve the islets during travel time for recipients living far away from transplant centers. Another advantage for culture is volume reduction of transplanted tissue by eliminating acinar cells and/or dead cells. One of the ideas behind islet culture is to select healthy islets from predamaged ones in order to minimize proinflammatory and proapoptotic effects in the graft (36). Moreover, culture at room temperature was initially introduced as a means of reducing islet antigenicity (15,32). We will evaluate the relationship of the effect of islet equivalent and tissue volume carefully and the antigenicity of fresh and cultured islets for a future study.
In conclusion, fresh islets are superior to cultured islets for human islet transplantation. The fresh islet transplantation could avoid the deterioration of islets during culture and improve the outcome of islet transplantation. Although culture of islets provides many benefits to clinical islet transplantation such as islet quality testing, stability during travel and treatment time, we should avoid islet culture or, at least, curtail culture time until the establishment of a good culture condition for human islets. In our recent clinical study, we performed fresh islet transplantation. Since islet culture before islet transplantation has several advantages, our future work will optimize the culture conditions to minimize islet loss before clinical islet transplantation.
Footnotes
Acknowledgments
The authors wish to thank Ms. Yoshiko Tamura and Mr. Greg Olsen for technical support. This work was supported in part by the Juvenile Diabetes Research Foundation International (JDRFI); Otsuka Pharmaceutical Factory, Inc.; and the All Saints Health Foundation. The authors declare no conflict of interest.