
Abstract
Pluripotent human embryonic stem (hES) cells are an important experimental tool for basic and applied research, and a potential source of different tissues for transplantation. However, one important challenge for the clinical use of these cells is the issue of immunocompatibility, which may be dealt with by the establishment of hES cell banks to attend different populations. Here we describe the derivation and characterization of a line of hES cells from the Brazilian population, named BR-1, in commercial defined medium. In contrast to the other hES cell lines established in defined medium, BR-1 maintained a stable normal karyotype as determined by genomic array analysis after 6 months in continuous culture (passage 29). To our knowledge, this is the first reported line of hES cells derived in South America. We have determined its genomic ancestry and compared the HLA-profile of BR-1 and another 22 hES cell lines established elsewhere with those of the Brazilian population, finding they would match only 0.011% of those individuals. Our results highlight the challenges involved in hES cell banking for populations with a high degree of ethnic admixture.
Introduction
The pluripotency of embryonic stem (ES) cells has rendered them a potential source of tissues for transplantation. For the last two decades, this property has been demonstrated in animal models of several conditions, including diabetes [reviewed in (19)], spinal cord injury, and Parkinson's disease [reviewed in (14)]. The establishment of hES cells in 1998 (35) has increased the expectations regarding the therapeutic use of these cells in patients. However, in addition to the concerns about the tumorigenic potential of hES cells, issues regarding the immunogeneicity of tissues derived from these cells, and the standardization of production of cells as pharmacological agents may hinder their use in humans.
As for other sources of tissues and cells for transplantation, the risk of immune rejection of hES-derived cells is correlated with the presence of cell surface antigens at the major histocompatibility complex (MHC), in humans represented by the HLA (human leukocyte antigen) proteins. Thus, the higher the similarity among HLA types, the lesser the chance of rejection, increasing the likelihood of transplant survival (31). However, the genes encoding for HLA are very polymorphic in our species (34), making it extremely difficult to obtain a perfect match between donor and recipient.
A review of the literature shows that the issue of the immunogenicity of hES cell-derived tissues is still not clear. These cells express HLA-I genes, albeit in lower levels than somatic cells do (9). On the other hand, these pluripotent cells show imunogeneicity in mice, where immunosuppressive therapy must be employed in order to avoid rejection of hES cells by the host immune system (32).
Strategies to reduce immunogenicity of allogenic hES cell-derived grafts include the generation of patient-specific cell lines by somatic cell nuclear transfer [reviewed in (39)] or by reprogramming adult cells into a pluripotent state [induced pluripotent stem (iPS) cells] with different transcription factors [reviewed in (38)]. Although in theory these methods would circumvent the issue of immunocompatibility, the generation of autologous tissues for transplantation is in practice extremely demanding and expensive. Furthermore, current methods for generating iPS cells involve genetic modifications, which increase the risk of their therapeutic use. Finally, because patient-specific pluripotent cell lines would not be universal therapeutic agents, each cell line generated will probably be treated as a separate product, and in that case will have to go through extensive preclinical and clinical testing before being approved by the regulatory bodies for therapeutic use (7).
An alternative to reduce immunogenicity of allogenic hES cell-derived grafts is banking of MHC-compatible hES cell lines (33). It is still debatable how many genetically diverse cell lines such a bank would need in order to attend different populations. The numbers vary from hundreds (28,33) to millions (5,8) of lines, depending on the acceptable degree of immune mismatch, and on the genetic diversity of the population under consideration.
Equally important for regenerative therapies is the standardization of defined conditions and processes to be used in manufacturing hES cell-based therapeutic cells (i.e., culture and differentiation of the cells) (11). Although the culture of hES cells in defined medium has been reported by different groups (24,40), there is only one report of the establishment of new lines of hES cells in defined medium (25). However, one of the lines was aneuploid already in early passages, while the other became aneuploid after 4–7 months in culture.
Here we describe the derivation of a new hES cell line, called BR-1, using a defined commercial medium. This hES cell line expresses markers of pluripotency, can be differentiated into tissues of the three germ layers, and presents a stable normal karyotype after 6 months of continuous culture in defined medium. We compare its HLA profile to those of a subset of the Brazilian population, and discuss the implications for hES cell-based therapy in ethnically diverse populations. To our knowledge, BR-1 is the first hES cell line derived in South America, and the characterization of its genomic ancestry raises important issues regarding hES cell banking to attend genetically admixed populations.
Materials and Methods
Establishment of hES Cell Lines
Couples donated surplus embryos from in vitro fertilization (IVF) cycles according to the Brazilian legislation (Bio-safety Law 11.105, March 25, 2005), signing the informed consent, and with the approval of the Ethics Committee of the Instituto de Biociências of the Universidade de São Paulo. Whole embryos at blastocyst stage or mechanically dissected inner cell masses were plated on four-well plates coated with hES-certified Matrigel (BD Biosciences) and cultured with mTeSR1 defined medium (StemCell Technologies), a serum-free medium that includes high levels of growth factors like bFGF and TGF-β. No additional component was added to the medium. After 48 h, half of the medium was changed every other day for 7–12 days. Outgrowths were mechanically passaged into new wells coated with Matrigel or mouse irradiated feeder cells (MEFs; Chemicon Millipore). After the first passage (p1), half-medium change was performed every day with mTeSR1. Colonies were mechanically passaged every 5–7 days, and after the third passage half of the cell clumps obtained from mechanical dissociation of the colonies was frozen in 90% fetal bovine serum (FBS, Hyclone) and 10% DMSO (Sigma).
Immunocytochemistry
Cells from passages 6 and 29 were fixed and immunostained following standard protocols. Briefly, cells were fixed with 4% paraformaldehyde for 10 min, permeabilized in 0.1% Triton X-100 solution (Sigma), and blocked in 1% fetal bovine serum (FBS; Hyclone) for 1 h. Cells were then incubated with primary antibodies overnight at 4°C, washed in PBS, and incubated with Cy3-conjugated mouse IgG antibodies (1:200; GE Healthcare) for 60 min at room temperature. Slides were prepared using Vectashield with DAPI (Vector Laboratories, Inc) for nuclei counterstaining. Primary antibodies used were: monoclonal anti-mouse stage-specific embryonic antigen-1 (SSEA-1) and -4 (SSEA-4), TRA-1-60, TRA-1-81 (dilution 1:50; Chemicon), and anti-mouse OCT-4 (dilution 1:100; Santa Cruz Biotechnology, Inc.) Human foreskin fibroblasts (HS27 cell line, ATCC) were used as negative controls.
In Vitro Differentiation
For embryoid body (EBs) formation, cell aggregates were grown in suspension in nonadherent plates as described by Itskovitz-Eldor et al. (16). The differentiation medium consisted of 80% DMEM, 20% FBS (Hyclone), 1% Glutamax (Invitrogen), and 1% penicillin-streptomicin (Invitrogen). After 1 week, EBs were collected and transferred to 0.1% gelatin-coated dishes for one more week, and then fixed in 4% PFA and subjected to immunocytochemistry assay as described above. The primary antibodies used were: endoderm: anti-goat SOX-17 and anti-goat HNF-3β (Santa Cruz Biotechnology, Inc.); mesoderm: α-actinin (Sigma), and mouse anti-MYOD (BD Pharmingen); ectoderm: MAP2 (Sigma) and NF200 (Sigma). For secondary antibodies anti-goat IgG-FITC (Santa Cruz Biotechnology, Inc.) or anti-mouse IgG-Cy3 (GE Healthcare) were used.
Teratoma Assays
Cells from passage 12 were cultured both on Matrigel and on feeder cells. Undifferentiated colonies were dissected mechanically, and approximately 100 cell aggregates (approximately 1 × 105 cells) were injected subcutaneously in C57/Bl6 nude mice. Teratoma formation was followed for 10-12 weeks after injection. Tumors were dissected, fixed in 4% PFA, embedded in paraffin, and submitted to histological analysis as described (35).
Karyotype and Microarray Comparative Genomic Hybridization (CGH) Analysis
Standard G-banding karyotype analysis was performed in BR-1 cells from passage 6 and passage 29 as described elsewhere (13). At least 20 metaphases were analyzed in each group. CGH-array was performed using an array containing 44,000 oligonucleotides (Human Genome Microarray CGH 44A, Agilent Technologies), as described elsewhere (20). Briefly, genomic DNAs from passages 4 and 29 of BR-1, and a normal female were labeled by random priming with Cy3- and Cy5-dCTPs, respectively. Purification, hybridization, and washing were carried out as recommended by the manufacturer. Scanned images of the arrays were processed with Agilent Feature Extraction v9.5.1 and analyzed using CGH Analytics software 3.4.40 (both from Agilent Technologies) with the statistical algorithm ADM-2, and sensitivity threshold 6.0. At least three consecutive oligonucleotides had to have aberrant log2 ratios to be called by the software. The average resolution was approximately 70 kb.
HLA Typing and Analysis of Genomic Ancestry
Genomic DNA from BR-1 was used for HLA-typing of loci A, B, Cw, DRB1, and DQB1 at medium resolution, and was performed by the Laboratório de Histocompatibilidade (UNIFESP, Brazil) using the LABType® SSO typing system according to the manufacturer's instructions (OneLambda, Inc). HLA profile of different hES cell lines was obtained from the literature (17,21) or from the International Stem Cell Registry (15) and the European Human Embryonic Stem Cell Registry (10). HLA matching at low resolution for loci A, B, and DRB1 was performed against 1,312,334 individuals from the Brazilian National Registry of Bone Marrow Donors (REDOME) using the Sismatch software (4).
Genomic ancestry was determined by analysis of 40 insertion/deletion (indel) polymorphisms along the autosomes as described by Bastos-Rodrigues et al. (3). These experiments were performed by Laboratório Gene (Belo Horizonte, MG, Brazil).
Results
Seventy-nine surplus embryos produced for reproductive purposes were thawed and allowed to develop until the blastocyst stage, on day 5. Inner cell masses (ICM) were mechanically isolated from 5 of a total of 45 blastocysts obtained, and the other 40 poor quality blastocysts had only the zona pellucida (ZP) mechanically removed. Both isolated ICMs and whole blastocysts were plated on Matrigel-coated plates and cultured in mTeSR1 medium (StemCell Technologies). From this day on, outgrowths were observed and had half of the medium changed every other day until the first passage (p1), after which complete medium change was performed every day.
Twenty one plated blastocysts (3 isolated ICMs and 18 whole blastocysts) attached and grew in culture with mTeSR1 and Matrigel for 7–10 days. We then added mouse irradiated feeder cells (MEFs) to 7 surviving outgrows, and 13 days later, cell clumps from one outgrow (from a whole blastocyst) were mechanically passaged to Matrigel-coated dishes with mTeSR1. Colonies exhibited the characteristic morphology of hES cells, with each cell showing a high nucleus/cytoplasm ratio, forming tightly, compact colonies that were mechanically dissociated every 6–8 days and cultured in feeder-free conditions (Fig. 1). This line, named BR-1, was successfully propagated and characterized.

Establishment of a new line of hES cells: light microscopy of cells at different days after plating. Dotted circles indicate cell masses with hES cell-like morphology that were mechanically passaged. Inset shows colony at higher magnification (50×), where cells with the characteristic high nucleus to cytoplasm ratio can be observed. Scale bar: 100 μm.
Cells were assayed for antigens correlated with pluripotency by immunofluorescence (Fig. 2). Colonies show a clear nuclear staining pattern for OCT4, and cell surface staining for TRA-1-60, TRA-1-81, and SSEA-4.

Expression of pluripotency markers. Immunofluorescence of BR-1 cells at passage 6 with antibodies against markers of pluripotency. Cy3-conjugated anti-mouse IgG (red) was used as secondary antibody. Nuclei were stained with DAPI (blue). Scale bar: 100 μm.
We verified the potential of BR-1 cells to differentiate into the three germ lineages in vitro. Cells were cultured in basic medium without FGF in nonadherent culture plates, allowing the formation of embyoid bodies (EBs). After 1 week in suspension, EBs were transferred and cultured 1 more week in gelatin-coated wells. Immunostaining of differentiated cell confirmed differentiation in endoderm [SOX17 and HNF3β (FOXA)], mesoderm (α-actinin and MYOD), and ectoderm [NF200 (NEFH) and MAP2] (Fig. 3).

In vitro differentiation of BR-1. Immunofluorescence of in vitro differentiated BR-1 cells, expressing markers of endoderm (SOX-17 and HNF-3β), mesoderm (α-actinin and MYOD), and ectoderm (MAP2 and NF200). FITC-conjugated anti-goat (green) and Cy3-conjugated anti-mouse (red) were used as secondary antibodies. Nuclei were stained with DAPI (blue). Scale bars: 100 μm.
Pluripotency of BR-1 cells was also tested by teratoma formation assays. Ten to 12 weeks after subcutaneous injection of undifferentiated cell clumps into nude mice, tumors with both cystic and solid areas were visualized. Histological analysis of the teratomas revealed the presence of tissues derived from mesoderm, endoderm, and ectoderm, demonstrating the pluripotency of BR-1 (Fig. 4).

Teratoma formation in nude mice. Histological analysis of hematoxylin-eosin-stained images of sections of teratomas derived from BR-1 hES cell line showing derivatives from the three embryonic germ layers. ad, adipose; c, cartilage; ge, glandular epitethelium; m, muscle tissues; ne, neural epithelium; nr, neural rosettes; v, vessel. Scale bars: 100 μm.
Chromosomal stability of BR-1 was assayed by both conventional karyotyping and by CGH-array (Fig. 5). Cells at passage 6 and 29 were karyotyped by standard G-banding. At least 20 metaphases were counted, all showing a normal 46,XY chromosome constitution (Fig. 5A). For analysis at higher resolution, CGH-array was performed, detecting no chromosomal loss or gain at the 70-kb resolution in cells cultured continuously for 6 months, at passage 29 (Fig. 5B). As a positive control, sex chromosome imbalances between BR-1 and the female reference sample were detected. These data indicate that the culture conditions were adequate for derivation and propagation of a karyotypically normal and stable hES cell line. BR-1 has also been successfully propagated on MEFs from passage 2 on, and will be characterized at a later date.

Genomic stability of BR-1 cells. (A) Karyotype with G-banding and (B) CGH-array of cells at passage 29. DNA from a normal female (46,XX) was used as reference. Blue lines indicate differences between BR-1 and the reference DNA. The detailed view of chromosomes X and Y shows lower (green dots) and higher (red dots) content of X and Y sequences, respectively, in BR-1 when compared to the reference DNA.
The HLA profile at low resolution of BR-1 and those of 22 other hES cell lines was compared with the Brazilian Bone Marrow Donor Registry (REDOME), consisting of HLA typing of 1,312,334 Brazilians (Table 1). In that subset of the Brazilian population we found no full match for BR-1, nor for cell lines derived in Israel, India, or Spain. In contrast, at least one full match was found to lines established in the US, Singapore, and Sweden. Taken together, the 23 lines analyzed were a match for 0.011% (150/1,300,000) of the Brazilian population represented in REDOME's database.
HLA Matching of Different hES Cell Lines to the Brazilian Population
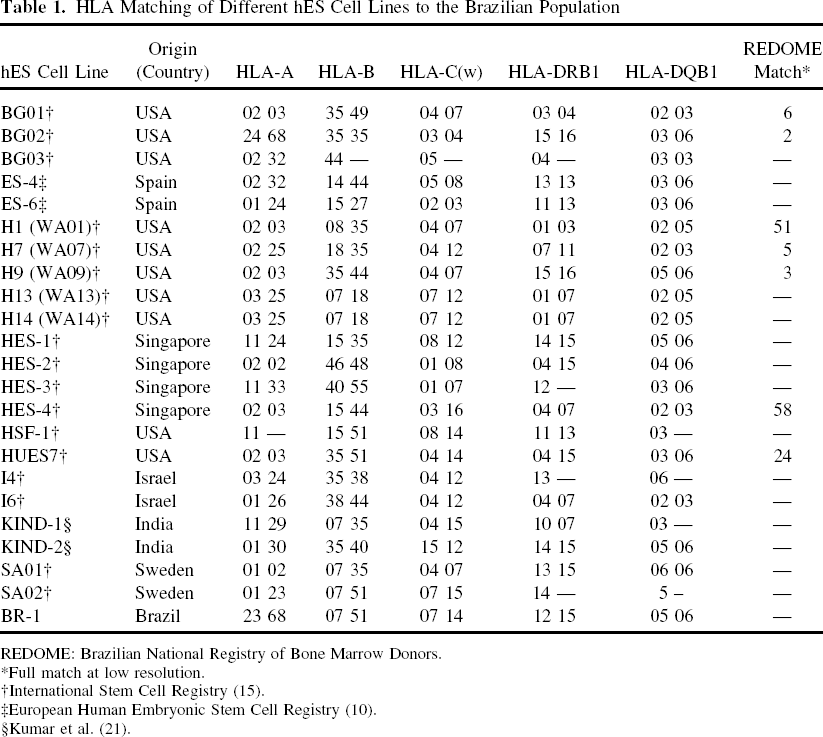
REDOME: Brazilian National Registry of Bone Marrow Donors.
Full match at low resolution.
International Stem Cell Registry (15).
European Human Embryonic Stem Cell Registry (10).
Kumar et al. (21).
Genotyping of 40 indel polymorphisms on autosomes (genotypes available upon request, or at www.ib.usp.br/lance_usp) revealed that the genomic ancestry of BR-1 is 98.4% European, 0.8% Amerindian, and 0.8% African (Table 2). The same analysis showed that the genomic ancestry of H1 is also predominantly European (96.0%).
Genomic Ancestry of BR-1 and H1

Values are shown as percentage of each contribution (confidence interval).
Discussion
As of August 2009 there were more than 300 different lines of hES cells deposited in the International Stem Cell Registry (15). Therefore, what are the motivations to establish new lines? One important reason is the standardization of protocols for derivation and in vitro maintenance of hES cells. In particular, defined conditions for establishing and culturing hES cell should facilitate replication of experiments with those cells in both the basic research and clinical settings. Although defined conditions for culturing hES cells have been described by several groups [reviewed in (37)], to date there is only one report of derivation of hES cell lines in defined medium, TeSR, which differs from mTeSR1 used in this study only by the replacement of bovine by human albumin (25,26). However, while one cell line was aneuploid in early passages, perhaps due to an alteration already present in the original embryo, the second line became trisomic for chromosome 12 after 7 months in culture. In contrast, BR-1 remained karyotypically stable at passage 29 even at the 70-kb resolution. Therefore, our results show that a defined medium can support the establishment of a karyotipically normal hES cell line. We are currently working on the establishment of new hES cell lines in defined xeno-free medium, which will further facilitate the clinical use of tissues derived from those cells.
The second issue relates to the need to increase the diversity of HLA types in stem cell banks, which in turn will be important to cater to ethnically heterogeneous populations in the clinical setting. HLA genes are among the most polymorphic in the entire genome (18,34). There are hundreds of reported alleles for each locus, and the number is still rising as new ones are described. Currently, there is little information about HLA allele frequencies in admixtured populations (1). In particular, Brazil has a population of approximately 190 million individuals with a uniquely high degree of admixture, having different degrees of African, European, and Amerindian ancestry (30). For Brazilian patients subjected to bone marrow transplants, most nonconsanguineous donors are found within the Brazilian population. Therefore, we hypothesized that BR-1 would be a better match for the Brazilian population than hES cells derived elsewhere. However, BR-1 was a worse match to the Brazilian subpopulation represented by approximately 1.3 million individuals registered at the REDOME than seven lines derived elsewhere (Table 1). Interestingly, the hES cell lines with most matches to individuals in that register were derived in the USA (H1; 51 matches) and Singapore (HES4; 58 matches). Nevertheless, those 23 cell lines were full matches at low resolution to only 0.011% of the Brazilian population. This is in strong contrast with results obtained by a similar analysis performed in the Chinese population (23), where it was found that a local hES cell bank with 188 lines provides a full match to 24.9% of that population. It will be important to have a greater number of hES cell lines derived from the Brazilian population in order to evaluate whether international or national hES cell banks may better cater to this population.
Finally, another motivation for the derivation of new hES cell lines is to increase the number of genetically variant cell lines available for research and therapy. Indeed, two recent studies of the genetic background of 86 hES cell lines concluded that, despite being from different geographical areas, they were from very restricted ethnic origins, basically European and Middle Eastern/East Asian (22,27). Both studies concluded that there is currently a lack of representative lines from populations worldwide, and reinforced the need for derivation of hES cells from underrepresented groups.
The Brazilian population is one of the most heterogeneous worldwide, a result of its history of colonization and five centuries of interethnic crosses among Europeans (mostly Portuguese), Africans, and Amerindians (29). Therefore, the genomes of most Brazilians are mosaics, having alleles from different phylogeographical origins (29). The high degree of admixture of the Brazilian population can thus be explored for the derivation of hES cell lines with novel genetic backgrounds. However, the analysis of the genomic ancestry of BR-1 showed that it is mostly of European origin (98.4%), similar to the H1 line derived in the US, and not representative of the mosaic genome characteristic of the Brazilian population, where the European contribution ranges from 37% to 82% (30).
Despite being based on a single cell line, our results highlight a limitation that may exist in the pool embryos available for research in Brazil. Although the legislation approved in 2005 was pivotal in allowing hES cell research in Brazil (36), it permits only the use of embryos produced for reproductive reasons, and that have been frozen for more than three years. However, assisted reproduction offered by the Brazilian public health system in general does not include cryopreservation of surplus embryos, and therefore the only material available for research comes from private clinics. Assuming that only those couples with a monthly income above 10 minimum wages (approximately US$2,500) are able to afford the high cost of private assisted reproduction, that Brazilian subpopulation would be mostly composed of people self-identified as caucasian (81% according to the demographic census of year 2000 from the Brazilian Institute of Geography and Statistics) (6). Despite the limitations of this classification (30), that group is likely not representative of the Brazilian ethnic admixture. Therefore, in order to explore the full genetic diversity of the Brazilian population for the establishment of ethnically and HLA-diverse hES cells lines, it will be important to have access to embryos from the more mixed population assisted by the Public Health System. These ethnically mixed hES cell lines may be a valuable resource for research and, in the future, for therapy in genetically heterogeneous populations.
In summary, we have shown that defined medium can support the derivation of a karyotipically stable hES cell line, and we have established the first line from the Brazilian population that adds to the pool of different HLA profiles of pluripotent cells available. Finally, we make an assessment of the compatibility of 23 hES cell lines with the Brazilian population, identifying a large limitation of currently available hES cells to provide compatible tissues for cell therapy in that large population. Recently, a first clinical trial with hES-derived cells was approved by the FDA (2). Although being currently on hold (12), those phase I experiments should provide important data on the real dimension of the issue of immunogeneicity of hES cells in humans. That in turn will enable the scientific community to design more efficient strategies to implement hES cell banking for therapy in different populations.
Footnotes
Acknowledgments
This work was supported by a grant from Conselho Nacional de Desenvolvimento Científico e Tecnológico/Departamento de Ciência e Tecnologia do Ministério da Saúde (CNPq/MS/DECIT). P.R. received a fellowship from Instituto Mara Gabrilli (img.org.br), and CordVida (www.cordvida.com.br). A.M.F. has a Ph.D. fellowship from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP). We are grateful to Dr. Jeanne Loring and the Center for Regenerative Medicine at The Scripps Research Institute for scientific and technical support, to Dr. Ric Ross for assistance with embryos manipulation, to Maria R. Pinheiro and Dr. Carla Rosenberg for genomic stability analysis, and to the Instituto Nacional do Cancer (INCA) and the REDOME for the HLA matching analysis.