
Abstract
Parkinson’s disease (PD) pathology involves progressive degeneration and death of vulnerable dopamine neurons in the substantia nigra. Extensive axonal arborization and distinct functions make this type of neurons particularly sensitive to homeostatic perturbations, such as protein misfolding and Ca2+ dysregulation. Endoplasmic reticulum (ER) is a cell compartment orchestrating protein synthesis and folding, as well as synthesis of lipids and maintenance of Ca2+ homeostasis in eukaryotic cells. When misfolded proteins start to accumulate in ER lumen the unfolded protein response (UPR) is activated. UPR is an adaptive signaling machinery aimed at relieving of protein folding load in the ER. When UPR is chronic, it can either boost neurodegeneration and apoptosis or cause neuronal dysfunctions. We have recently discovered that mesencephalic astrocyte-derived neurotrophic factor (MANF) exerts its prosurvival action in dopamine neurons and in an animal model of PD through the direct binding to UPR sensor inositol-requiring protein 1 alpha (IRE1α) and attenuation of UPR. In line with this, UPR targeting resulted in neuroprotection and neurorestoration in various preclinical animal models of PD. Therefore, growth factors (GFs), possessing both neurorestorative activity and restoration of protein folding capacity are attractive as drug candidates for PD treatment especially their blood-brain barrier penetrating analogs and small molecule mimetics. In this review, we discuss ER stress as a therapeutic target to treat PD; we summarize the existing preclinical data on the regulation of ER stress for PD treatment. In addition, we point out the crucial aspects for successful clinical translation of UPR-regulating GFs and new prospective in GFs-based treatments of PD, focusing on ER stress regulation.
Keywords
INTRODUCTION
Is ER stress inducing PD pathology?
Parkinson’s disease (PD) is characterized by progressive loss of midbrain dopamine (DA) neurons and accumulation of proteinaceous intracellular inclusions called Lewy bodies. These inclusions are mostly composed of misfolded phosphorylated α-synuclein (αSyn), as well as other proteins, membrane fragments and dysmorphic organelles [1, 2]. To date, misfolded αSyn is considered to be the main inducer of PD pathology and neurodegeneration through multiple mechanisms, such as disruption of axonal transport and loss of synapses, as well as impairment of mitochondrial, lysosomal, proteasomal and endoplasmic reticulum (ER) functions [3].
About 7 million people worldwide have PD; the majority of the cases are sporadic and only 5–10% are familial. To date, more than 20 genes mutated in familial cases of PD and genetic risk factors have been identified; many of these genes are involved in the maintenance of ER and mitochondrial protein homeostasis (Table 1). Mutations in the GBA gene, encoding lysosomal enzyme glucocerebrosidase 1, are responsible for 5–10% of all PD cases and considered to be the most common risk factor; this highlights the importance of maintaining proteostasis through lysosomal activity. Other common risk factors for PD are mutations in the leucine-rich repeat serine/threonine-protein kinase 2 (LRRK2) gene and the SNCA gene encoding for αSyn, present in 2% and 1–2% of all PD cases, respectively. Mutations in other genes, indicated in Table 1, are risk factors for PD as well, although they are less common and autosomal recessive.
Genes mutated in PD are involved in the maintenance of protein homeostasis
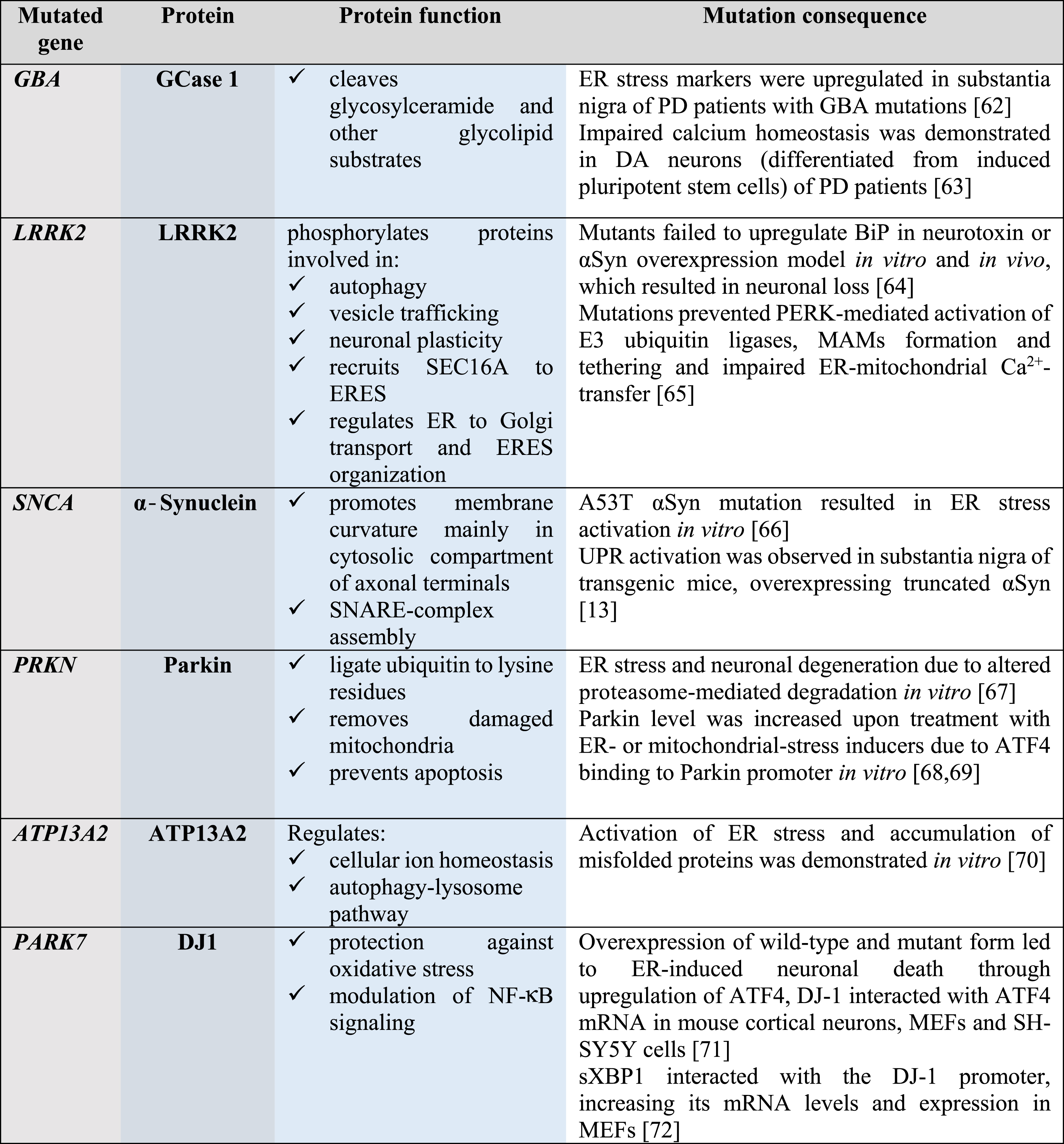
GCase 1, glucocerebrosidase 1; LRRK2, leucine-rich repeat kinase 2; αSyn, α-Synuclein; Parkin, E3 ubiquitin ligase; ATP13A2, polyamine-transporting ATPase 13A2; DJ1, protein/nucleic acid deglycase 1; SEC16A, protein transport protein; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; ER, endoplasmic reticulum; ERES, ER-exit sites; NF-κB, nuclear factor-κB; PD, Parkinson’s disease; BiP, immunoglobulin heavy-chain-binding protein, major ER chaperone; PERK, protein kinase RNA-like ER kinase; UPR, unfolded protein response; MAMs, mitochondria-associated membranes; ATF4, activating transcription factor 4; SH-SY5Y, neuroblastoma cell line; sXBP1, spliced X-box binding protein-1; MEFs, mouse embryonic fibroblasts.
Exceptionally vulnerable DA neurons of the subtantia nigra (SN) drastically differ in terms of morphology and functions from other neurons. In long and highly branched neurites of aging DA neurons, maintenance of ER protein homeostasis and axonal transport of mRNA, proteins and organelles is particularly challenging. High demand in mRNA transport and protein translation, especially translation of synaptic proteins, including αSyn, overloads the ER proteostasis machinery and results in protein misfolding. Impaired ER proteostasis, i.e., accumulation of misfolded proteins, impairment of Ca2+ homeostasis and dysregulation of axonal transport led to prolonged ER stress, playing an essential role in PD pathology [4].
ER stress was shown to accompany PD pathology; however, to date, the cause-effect relationship between ER stress and PD is uncertain. ER stress was induced in neurotoxin models of PD in vitro. DA neurons generated from human induced pluripotent stem cells derived from PD patients carrying SNCA or GBA mutations showed increased ER stress, elevated ER stress markers and impaired ER-associated degradation (ERAD) [6, 7]. Increased levels of activated ER stress sensor PERK and its downstream target phosphorylated eIF2α (p-eIF2α) were found in postmortem midbrain samples of early stage PD patients [8].
Since impaired ER proteostasis is associated with aging and PD pathology [4], re-establishment of ER proteostasis may be considered as a disease-modifying strategy. This can be achieved through the targeting of components of ERAD, the ER Ca2+ homeostasis, as well as the functioning of ER chaperones or isomerases, and ER stress-activated signaling receptors or their regulators.
Endoplasmic reticulum stress triggers unfolded protein response activation
ER is a cellular compartment responsible for synthesis of both secretory and membrane proteins, their modification, folding and secretion as well as for lipid synthesis and calcium homeostasis. Protein misfolding, aggregation and homeostasis perturbations in the ER lumen lead to ER stress triggering coordinated adaptive signaling machinery called the unfolded protein response (UPR). Adaptive UPR is aimed at restoring cellular homeostasis by increasing protein folding capacity through attenuation of protein synthesis, selective mRNA degradation, degradation of misfolded proteins and upregulation of transcription of genes involved in ERAD, autophagy, lipid biosynthesis, and Ca2+ homeostasis [9]. Prolonged terminal UPR, studied mainly in secretory cells and not well documented in neurons, can lead to activation of pro-apoptotic and pro-inflammatory signaling contributing to cell death [10]. In mammalian cells, UPR occurs through three ER transmembrane proteins termed inositol-requiring protein 1 alpha (IRE1α), protein kinase RNA-like ER kinase (PERK) and activating transcription factor 6 (ATF6) (Fig. 1).
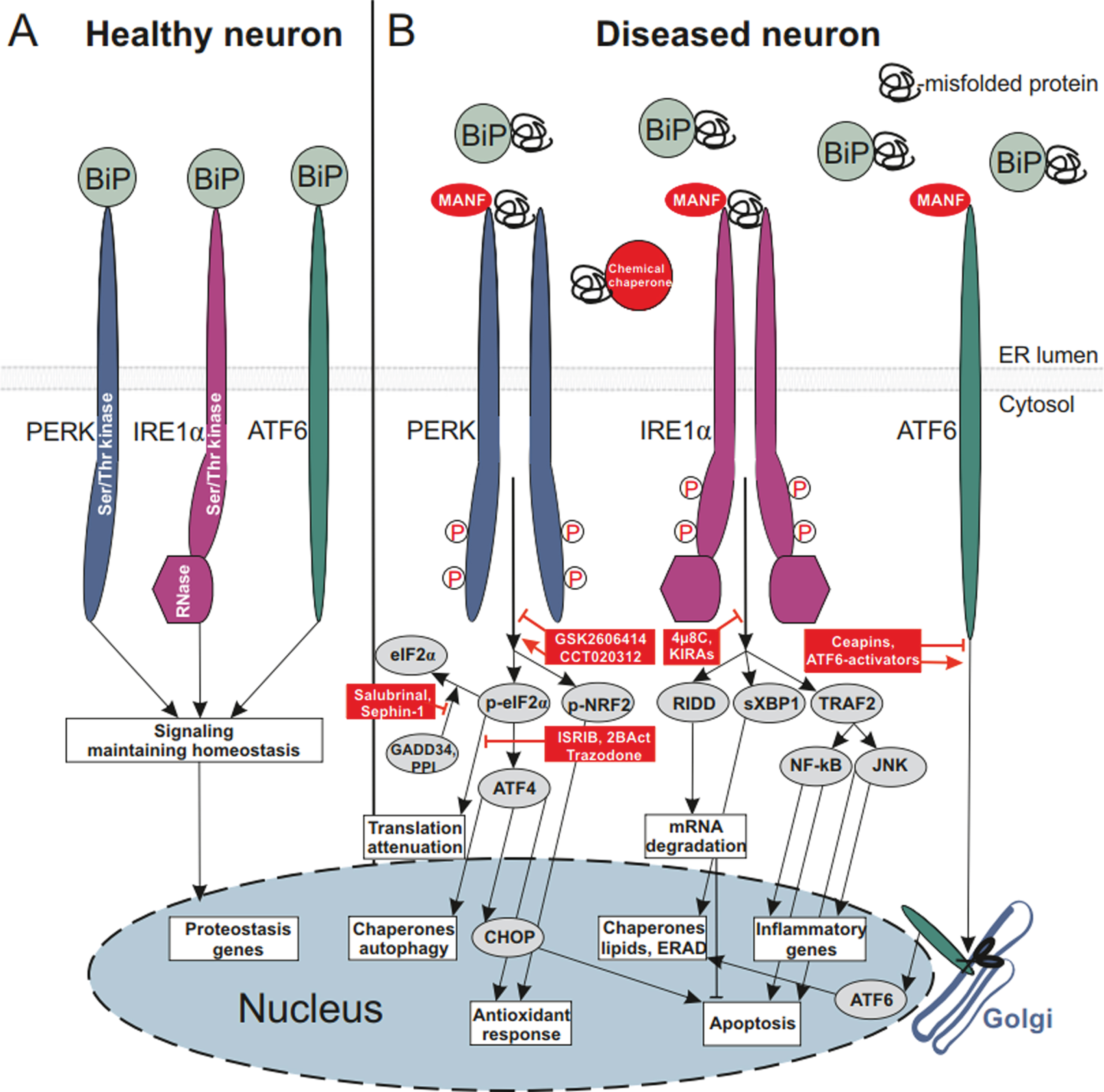
ER-stress signaling and ER stress-modulating proteins and small molecule compounds affecting neuronal survival. A) Healthy neuron, main ER chaperone BiP (alias GRP78) is bound to luminal domains (LDs) of monomeric transmembrane UPR sensors protein kinase RNA-like ER kinase (PERK), inositol-requiring protein 1 alpha (IRE1α) and activating transcription factor 6 (ATF6) and keeps these inactive. Signaling is aimed at maintaining protein homeostasis and classical UPR signaling is not triggered. B) Diseased neuron, ER is overloaded with misfolded proteins, BiP dissociates from LDs of UPR sensors to bind misfolded proteins and re-establish homeostasis; dissociation of BiP from UPR sensors leads to their dimerization and phosphorylation of IRE1α and PERK, and activates cytoplasmic UPR signaling [73]. Alternatively, upon BiP dissociation from UPR sensors their full activation occurs through the direct binding of misfolded proteins to their luminal domains, as was demonstrated for IRE1α and PERK LDs [74, 75]. BiP dissociation from PERK LD leads to dimerization of PERK LDs, allowing dimerization and autophosphorylation of the cytoplasmic kinase domain of PERK. Active PERK phosphorylates two substrates: α-subunit of eukaryotic initiation factor 2 (eIF2α) and nuclear factor erythroid 2-related factor 2 (NRF2). This step can be modulated using the PERK inhibitor GSK2606414 or the PERK activator CCT020312; both of these compounds have been shown to be neuroprotective in different in vitro and in vivo models of neurodegeneration. Through phosphorylation of eIF2α PERK attenuates global protein translation, decreasing the loading of ER with misfolded proteins. However, chronic attenuation of protein translation, continuously repressing the expression of synaptic proteins, can be detrimental for neurons. Few compounds blocking translation attenuation, such as integrated stress response inhibitor (ISRIB), its improved derivative 2BAct, trazodone hydrochloride and dibenzoylmethane (DBM) have been shown to be cytoprotective. Blockade of protein phosphatase 1 (PPI)-mediated dephosphorylation of eIF2α through growth arrest and DNA damage-inducible protein (GADD34) using salubrinal, sephin-1 or guanabenz was neuroprotective. In addition to attenuation of protein translation, phosphorylation of eIF2α selectively enhances translation of activating transcription factor 4 (ATF4), promoting transcription of genes responsible for chaperone expression, autophagy and antioxidant response. ATF4 also induces the expression of a transcriptional factor C/EBP Homologous Protein (CHOP), upregulating the expression of genes involved in apoptosis. PERK phosphorylates NRF2, which is then translocated to the nucleus and activates transcription of genes involved in antioxidant responses. BiP dissociation from IRE1α LD, similarly to dissociation from PERK LD, results in dimerization and autophosphorylation of IRE1α cytoplasmic kinase domain. Activation of the kinase domain leads to activation of IRE1α ribonuclease (RNase) domain. Activation of IRE1α kinase can be inhibited by the family of compounds called kinase-inhibiting RNase attenuators (KIRAs) and IRE1α RNase can be inhibited by 4μ8C. As an endoribonuclease, IRE1α degrades specific mRNAs via regulated IRE1α-dependent decay (RIDD), decreasing the load of misfolded proteins to ER. RIDD can also decrease apoptosis through degradation of death receptor 5 (DR5) mRNA [76]. Endoribonuclease of IRE1α also performs unconventional splicing of X-box binding protein-1 (XBP1), transcription factor spliced XBP1 (sXBP1), in turn, upregulates expression of the genes regulating chaperone, lipid biosynthesis, and ER-associated degradation (ERAD). Activation and oligomerization of IRE1α lead to the recruiting of TNF receptor-associated factor 2 (TRAF2), activating c-Jun N-terminal kinase (JNK) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathways; these regulate apoptosis and transcription-related inflammatory genes [77, 78]. Upon BiP dissociation from ATF6 LD, ATF6 is transported to the Golgi apparatus, where it is cleaved by site-specific proteases. Cleaved ATF6 translocates to the nucleus and activates transcription of the genes involved in chaperone and lipid biosynthesis, ERAD, as well as inducing XBP1 mRNA. ATF6 translocation from ER to the Golgi apparatus can be specifically inhibited by Ceapins or activated by small molecule ATF6 activators, compounds 147 and 263. Mesencephalic astrocyte-derived neurotrophic factor (MANF) directly interacts with UPR sensors and attenuates UPR signaling in DA neurons and in an animal model of PD [34]. MANF, its homologous protein CDNF and their small molecule mimetics represent a strategy to treat PD, attenuating ER stress and inflammation. ER stress-modulating interventions are shown with red color.
Upon ER stress IRE1α, possessing both serine-threonine kinase and endoribonuclease activity performs splicing of the mRNA encoding for X-box binding protein-1 (XBP1) [11]. Spliced form of XBP1 (XBP1s) activates prosurvival genes regulating ERAD, autophagy, and lipid biosynthesis. IRE1α endoribonuclease also decreases the ER load of misfolded proteins by degrading specific mRNAs in a process called regulated IRE1α-dependent decay (RIDD). PERK with cytoplasmic Ser/Thr kinase domain has two main targets: eukaryotic translational initiation factor 2α (eIF2α) and nuclear factor erythroid 2-related factor 2 (NRF2). Through the phosphorylation of eIF2α PERK along with other integrated-stress response kinases inhibits protein synthesis initiation by attenuating global protein synthesis, decreasing ER protein load. A chronic, unresolved UPR can trigger apoptosis through upregulation of C/EBP homologous protein (CHOP). During the global attenuation of protein translation, however, some proteins, such as activating transcription factor 4 (ATF4) are able to be synthesized and further regulate expression of specific UPR genes. Through the phosphorylation of NRF2, PERK triggers an antioxidant response [12]. Upon ER stress, the third UPR sensor ATF6 is transported to the Golgi apparatus where it is cleaved, and its cytoplasmic fragment enters the cell nucleus and triggers transcription of ER chaperones and ERAD genes.
Can α-synuclein induce endoplasmic reticulum stress in dopamine neurons?
To date, the cause-effect relationships between αSyn misfolding/aggregation and ER stress/UPR activation are still unclear. It has been shown that aggregated misfolded αSyn induces BiP (alias GRP78) and ATF4 expression in vitro and in a mouse model overexpressing truncated αSyn [13]. αSyn-overexpression induced UPR, inhibited protein trafficking from ER to Golgi apparatus and resulted in impaired protein maturation in yeast cells; re-establishing of ER-Golgi trafficking was shown to be protective from αSyn-mediated toxicity in yeast, DA neurons and αSyn-overexpressing Drosophila and C. elegans models of PD [14].
Interestingly, αSyn interacted with UPR sensor ATF6 and inhibited its activation by ER stress in in vitro study [15]. Overexpressed major ER chaperone, BiP, and αSyn were shown to form a complex, likely contributing to neuroprotective effect of BiP overexpression in αSyn-overexpressing rat model of PD [16]. Interaction of αSyn with BiP, was demonstrated also under physiological conditions, i.e., in non-transgenic mice, indicating that ER stress may contribute to aggregation of αSyn and progression of PD pathology [17]. Whether αSyn is directly interacting with ER chaperons and directly activating UPR in a physiologically relevant manner remains unknown. It is important to emphasize that αSyn is a cytoplasmic protein, whereas UPR activation occurs in the lumen of the ER. Thus, is it possible that αSyn can move from the cytoplasm to the ER lumen and interact there with BiP or with the luminal domain of ATF6? Many of the previous studies used overexpression of αSyn protein, and in those cases, the protein can be localized into cell compartments that are not occurring for physiological concentrations. Although no experimental data exist on αSyn movement from the cytoplasm to the ER, it can be possible, since αSyn fibrils very efficiently penetrate through cellular membranes of various cell types [18].
Although αSyn being considered a primary suspect in PD pathology, its overexpression is not always sufficient to induce robust degeneration of DA neurons in vitro and in vivo [19]. Noteworthy, in familial cases of PD, SNCA gene duplication or triplication induces PD pathology and neurodegeneration, with the early pathology onset in the case of triplication, suggesting that the level of αSyn correlates with the disease progression [20]. In line with this, a high increase (about 5-fold) in levels of αSyn resulted in pronounced neurodegeneration in AAV-αSyn transgenic rats [21]. However, in some studies, neurodegeneration was also observed with control proteins, such as GFP, after high overexpression [22]. Significant DA neuron loss was observed in animal models combining αSyn overexpression and inoculation with preformed αSyn fibrils [23], or when aggregation-prone truncated αSyn fragments were overexpressed [24]. Although quite a few experimental data demonstrate the induction of ER stress by αSyn and even direct interactions of αSyn with the UPR machinery components, it remains unclear whether αSyn can directly trigger ER stress activation. It needs to be clarified how cytosolic αSyn can modulate the processes occurring in ER lumen.
Can we treat Parkinson’s disease targeting ER proteostasis with UPR-regulating growth factors and small molecules?
To date, classically used for PD treatment GFs, glial cell line-derived neurotrophic factor (GDNF) and neurturin (NTRN), have not been shown to be involved in the regulation of ER stress and UPR, and only a few GFs were demonstrated to have a potential for alleviation of pathology-induced UPR.
Mesencephalic astrocyte-derived neurotrophic factor (MANF) and its paralog cerebral dopamine neurotrophic factor (CDNF) have been shown to protect DA neurons both in vitro and in vivo [25–28]. These neurotrophic factors reside in the ER lumen and alleviate ER stress in DA neurons and other cell types [28–33]. Recently we have discovered that MANF directly interacts with UPR sensor IRE1α and with lower affinities also with PERK and ATF6 and competes with BiP for the interaction with IRE1α [34]. Using computational modeling, mutagenesis, biochemical and cellular assays we have demonstrated that MANF–IRE1α interaction is crucial for pro-survival activity of MANF in DA neurons. We have shown that MANF is improving motor behaviour and increasing the number of DA neurons in SN in a neurotoxin rat model of PD, whereas MANF mutant unable to bind to IRE1α lacks neuroprotective activity in vivo [34]. The ability to be secreted upon Ca2+ depletion-induced ER stress [35] and exert protective properties when added extracellularly [28] hints at the existence of extracellular receptors or other mechanisms of internalization, such as lipid-mediated internalization, which is suggested by their structures [30, 36] and data on lipid binding [37]. Recent data show that binding of MANF, but not of CDNF, to cell surface neuroplastin receptors, reduced inflammatory responses and apoptosis via suppression of NF-κB signaling [38]. Unlike many GFs possessing mainly neuroprotective properties, MANF and CDNF, similar to GDNF and NRTN, also show neurorestorative action in DA neurons [26, 39–41]. Importantly, in addition to their neurorestorative action, MANF and CDNF alleviated UPR and reduced inflammation [38, 43]. Moreover, CDNF reduced αSyn aggregation in neurons, although in this study rather high CDNF concentration was used [44]. CDNF interacted directly with αSyn, reduced the levels of phosphorylated at Ser129 αSyn and alleviated motor deficits in rodents challenged with α-synuclein fibrils [45].
Modulation of the UPR components was already shown to regulate the survival of DA neurons in animal PD models. Interestingly, some UPR components, such as CHOP contributed to neurodegeneration, while others, such as sXBP1 and ATF6 supported neuroprotection [46–48]. Contribution of UPR-related genes in neurodegeneration and PD pathology has been reviewed earlier [4, 49].
Targeting ER proteostasis with small molecules was mainly performed through targeting the process of translational attenuation through PERK/p-eIF2α pathway. In a mouse neurotoxin PD model, the PERK inhibitor GSK2606414 protected DA neurons and restored motor function by increasing the level of DA and synaptic proteins [50]. In prion-diseased mice, GSK2606414 and de-repression of translational attenuation with integrated stress response inhibitor (ISRIB), were neuroprotective as well [51, 52]. Translational attenuator ISRIB had good general tolerability and no toxic effects in prion-diseased mice [52], suggesting that ISRIB and its improved derivative 2BAct [53] along with other non-toxic PERK/p-eIF2α modulators can be considered for future therapeutic use. Increasing p-eIF2α levels with salubrinal, a small molecule that inhibits p-eIF2α dephosphorylation through growth arrest and DNA damage-inducible protein (GADD34), led to improvement of motor function and rescued the phenotype in a mutant αSyn-overexpressing mouse model of PD [17]. However, in prion-diseased mice, having profound neurological deficits, salubrinal treatment led to neurodegeneration likely due to a significant reduction of protein synthesis compromising production of synaptic proteins [51].
Targeting ER proteostasis through IRE1α branch is aimed at regulation of sXBP1 activation or at decreasing the ER load of misfolded proteins through RIDD. Overexpression of sXBP1 was shown to be neuroprotective in an animal neurotoxin model of PD [47]. sXBP1 was also shown to improve synaptic plasticity and cognition through increasing the expression of brain-derived neurotrophic factor (BDNF) in vivo [54]. Effective small molecule modulators of the IRE1α branch, such as KIRA6 and KIRA8, have not yet been studied in animal models of PD. However, considering their cytoprotective action in pancreatic beta cells and their potential for diabetes treatment [10, 55], further pre-clinical studies in animal PD models are warranted.
Since ATF6α was shown to be involved in neuroprotection in a neurotoxin mouse model of PD [48], recently discovered specific activators of ATF6α, compounds 147 and 263 [56] and selective inhibitors of ATF6α, ceapins [57] hold great promise for PD treatment.
Modulation of UPR sensor activities is an attractive target for PD therapy not only because it is a way to prevent chronic terminal UPR, apoptosis and inflammation, but also to stimulate synaptic function and neuronal plasticity. This is why small molecules targeting different UPR components, already shown to be neuroprotective in various studies, is of particular importance (Fig. 1).
It has not escaped our notice that ER-mitochondria contact sites may play an important role in PD pathology. A few of PD-associated proteins, shown in Table 1, namely αSyn, Parkin, DJ1 and also PTEN-induced kinase 1 (PINK1) were demonstrated to localize to mitochondria-associated membranes (MAMs) and mutations in the genes encoding these proteins altered or disrupted ER-mitochondria signaling at MAMs [58]. That is why, MAMs should be considered as alternative targets for mechanistic studies and therapeutic approaches for PD.
CONCLUSION
Search for the effective UPR-regulating growth factor
Several gene therapies and protein clinical trials showed that GFs have a potential for PD treatment [41]. At the same time, the approaches and strategies for PD treatment need to be modified. The complications in the use of GFs and use of GF’s small molecule mimetics have been discussed earlier [41, 59]. We need to find, or develop, on the basis of existing proteins an effective UPR-regulating GF or even “GFs cocktail”. This potent GF should have good diffusion in brain tissue and not only protect, but also restore damaged midbrain DA neurons. In addition, it should alleviate ER-stress induced inflammation, block apoptosis and decrease the aggregation and neurotoxicity of αSyn.
UPR-regulating growth factors CDNF and MANF can protect and restore DA neurons, diffuse in the brain tissue much better, compared to classical GFs, alleviate inflammation and apoptosis and CDNF also decreases αSyn aggregation. MANF and CDNF, shown to act only on injured cells [28, 61], represent a promising strategy for PD treatment. Since MANF, CDNF and likely their mimetics bind only to hyperactivated UPR sensors, when in ER stressed cells BiP has dissociated from UPR sensors, their peripheral delivery is unlikely to cause serious adverse effects. Lack of the effects in healthy cells means the absence of side effects upon peripheral delivery. Systemically delivered GFs likely may be more potent due to treatment of not only motor, but also non-motor symptoms of PD. Taking into account the above-mentioned points and lower costs compared to proteins for production and delivery, MANF/CDNF-like peptides and small molecule mimetics seem to be good alternatives to native proteins in the context of clinical translation.
Since ER proteostasis is crucial for maintenance of functioning of DA neurons, new therapeutic approaches to target and regulate ER proteostasis and UPR components, such as UPR-regulating GFs and their mimetics, should be a prime target.
Footnotes
ACKNOWLEDGMENTS
The authors thank Professor Barry Hoffer, Professor Mikko Airavaara, Dr. Brandon K. Harvey, Dr. Maria Lindahl, Dr. Olesja Bondarenko, and Dr. Yulia A. Sidorova for critical comments on the manuscript. V.K. and M.S. are supported by a grant from the Academy of Finland (No. 1310891) and by Jane and Aatos Erkko Foundation grant.
CONFLICT OF INTEREST
The authors have no conflict of interest to report. MS is the inventor in the MANF- and CDNF-related patents owned by Herantis Pharma Plc. and a shareholder in this company.