
Abstract
BACKGROUND:
Cell surface molecules play important roles in cell signal transduction pathways during microbial infection.
OBJECTIVE:
In this study, the expression and the functions of CD59 was investigated in H. pylori infected gastric cancer (GC).
METHODS AND RESULTS:
The differential expression of CD59 and the influence of H. pylori on the expression of CD59 were analyzed via bioinformatics through Gene Set Enrichment in GC. In addition, the expression of CD59 in GES-1, AGS cells and GC tissues infected with H. pylori was confirmed by Western blot. Bioinformatics results and H. pylori infection experiments showed CD59 decreased obviously in H. pylori infected GC cells and tissues. The expression of CD59 was linked to the survival rate of GC patients, and influenced various immune cells in the immune microenvironment of GC. CD59 interacts with other genes to form a network in H. pylori infected GC. Certainly, CD59 decreased significantly in H. pylori infected GC tissues, GES-1 and AGS cells in vitro.
CONCLUSION:
H. pylori infection could influence the expression of CD59 in GC indicating that CD59 may be a promising treatment target.
Introduction
Gastric cancer (GC) is the fourth major cause of yearly cancer death rate because of its commonly later stage upon recognition, as reported by World Health Organization in 2018 [1]. GC especially attacks East Asia in terms of occurrence and mortality [2]. Basically, GC results from the complicated interaction among numerous critical factors from host genetics [3], lifestyle, the environment, and microbes [4]. Also gut microbiomes such as Helicobacter pylori participate in the host’s mucosal immune replies to discard pathogenic infection [5].
In developing countries, 70% to 90% of people are infected with H. pylori before age 10, but in developed countries, the incidence of infection varies from 25% to 50% [6]. The significant relevance of GC is H. pylori, a group I carcinogen [7]. The link between GC development and H. pylori infection is well established [8]. Proteins of cytotoxicity-associated gene A (CagA) and vacuolating cytotoxic A (VacA) in H. pylori on gastric mucosa cells induce complex bioeffects, including the generation of pro-inflammatory cytokines, involvement of immune cells to the infection site, and survival replies of gastric epithelial cells [9, 10]. Although H. pylori accounts for over 60% of GC worldwide [11], 10% of H. pylori infections are linked with peptic ulcer diseases, and 1%–3% of the infections will develops to GC with a surviving rate about under 5 years [12]. A further 0.1% will suffer mucosa-related lymphoid tissue lymphoma [13]. In addition, the treatment of H. pylori can decrease the risk of conversion to GC, but the risk decrease level relies on the extent of previous damage upon the termination [14].
GC activates the immune system to eliminate the malignant cells by T cells, monocytes, and natural killer cells. These leucocytes generate TNF-
This study will explore the CD59 expression in GC and H. pylori-positive GC cells through bioinformatics, and test the connection between CD59 and H. pylori infection as well as GC occurrence and development.
Materials and methods
Cell culture and H. pylori infection
GES-1 and AGS cells were offered by the Cell Bank of Type Culture Collection of Chinese Academy of Sciences (Shanghai, China). GES-1 and AGS cells were cultivated in culture dishes with DMEM added with 10% FBS, 100 U/mL penicillin, and 0.1 mg/mL streptomycin at 37
H. pylori was donated by Marshall International Digestive Disease Diagnosis and Treatment Center of Shanghai Oriental Hospital. H. pylori was inoculated on a blood culture medium and cultured in 95% CO
Infection was conducted at a multiplicity of cell: H. pylori
Tissues acquisition
GC tissues (10 cases) and GC tissues infected with H. pylori (10 cases) were purchased from Shanghai Xinchao Biotechnology Co., Ltd, and this study has been approved by the Ethics Committee of Kunshan First People’s Hospital (Suzhou, China, approval no. IEC-C-007-A07-V3.0).
Gene set enrichment analysis (GSEA)
RNA sequencing data were attained from GEO through GSEA (
Functional enrichment analysis of differential gene profiles
To test the common features of differential genes (DEGs), we searched the Venn diagram for overlapped differential genes, which were explored by the database for annotation, visualization and integrated discovery bioinformatics (//david.ncifcrf. gov/). Gene Ontology (GO) and KEGG pathway enrichment were analyzed to study possible roles of overlapped differential genes. The cutoff level was an interaction score
Analysis GSE60427 data by GEO database
GSE60427 data were downloaded from the GEO database in the MINiML format. Log2 processing was performed uniformly. The probe ID was converted into gene symbol according to the corresponding platform annotation information. The CD59 gene expression was extracted, and the differential expression of CD59 between H. pylori positive and negative patients was compared. The significance of the two groups was tested by Wilcox analysis. H. pylori-positive patients were divided into group 1 (low expression) and group 2 (high expression) of CD59. Then the differential expression of CD59 mRNA was studied with the package limma of R software 3.40.2. The 10 most significantly different genes were annotated. The GO function of underlying mRNAs was analyzed and the KEGG pathway was enriched on the package Cluster Profiler in R.
CD59 in cancers analyzed through TIMER2.0
The RNAseq data in the HTSeq-FPKM format from TCGA (//portal.gdc.cancer.gov/) STAD (Stomach Cancer) project were download. The format of the RNAseq data in was transformed from FPKM (fregments per kilobase per million) to TPM (transcripts per million reads). Then log2 conversion was done to extract the CD59 expression. The relationship between CD59 acquisition and cancer overall survival was analyzed using GEPIA2.
Analysis of CD59 in tumor immune
The abundance differences of 24 immune cells were predicted using the high and low expression groups of CD59. The unified and normalized pan-cancer datasets viz. TCGA, TARGET, and GTEx were downloaded, and CD59 gene and 60 immune checkpoint pathway genes were extracted and analyzed by Pearson correlation statistics (ns:
Protein-protein interaction (PPI) analysis
Interaction associations between differential genes were investigated by constructing a PPI network through a search tool with interacting genes (www.string-db.org/).
GSEA analysis was finished with the cluster Profiler. Gene set database was acquired from MSigDB Collections. False discovery rate (FDR)
Western blot
Cells and tissues were lysed with lysis buffer and then proteins were harvested. Proteins were resolved by 10% SDS PAGE and transferred onto PVDF membranes. The membranes were immunoblotted with primary antibodies CD59 (Proteintech, Inc., China) overnight at 4
Statistical analysis
All data were analyzed by GradphpadPrism 5 software with one-way analysis of variance (ANOVA) and are presented as mean e (ANOVA) re quantified with Imagepresented as percentage. A statistical significance was defined when
Results
Differential gene profiles and functions of GC cells infected with H. pylori analyzed with GES database
More than one hundred differential genes were revealed in the heat map and volcano map in H. pylori-infected GC cells relative to GC cell lines (Fig. 1A and B).
Differential genes regulated by H. pylori were involved in bioprocesses (BP), cellular components (CC) and molecular function (MF) analyzed through GO analysis (Fig. 1C). Differential genes in GC cells infected with H. pylori are enriched in PI3K-Akt, human cytomegalovirus infection, Ras and TNF signaling pathways through KEGG pathway analysis (Fig. 1D).
The interaction relationships of differential genes were displayed evidently in the network map (Fig. 1E). The top 20 interesting genes in the differential genes of H. pylori-infected GC cell lines were listed in Table 1. These results indicated that H. pylori infection could regulate various gene expression in GC involved in numerous biological functions of GC cells through multiple signaling pathways.
Top 16 interesting genes in GC cells infected with Hp analyzed by GESA database
Top 16 interesting genes in GC cells infected with Hp analyzed by GESA database
Differential gene profiles in GC cells infected with or without H. pylori by GES70394 analysis. (A) Heat map of differential genes in GC cells with or without H. pylori. (B) Volcano diagram and (C) GO enrichment map of differential genes including BP, MF and CC in two GC cell groups. (D) Bubble plot of differentially enriched genes in the KEGG pathway. (E) Interaction of differential genes in two GC cell populations.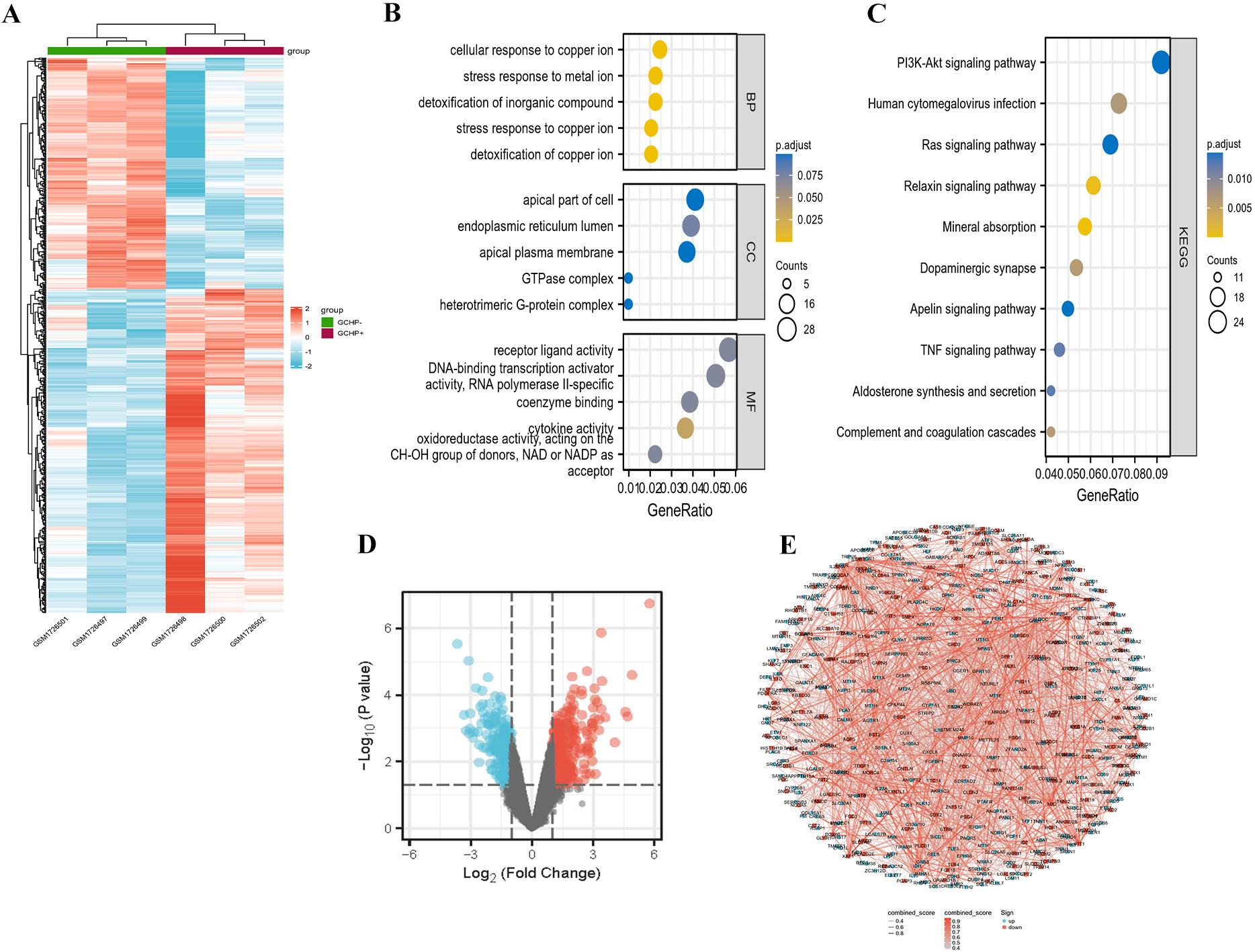
Analysis of association between CD59 expression and H. pylori infection. (A) CD59 gene expression between Hp negative and positive patients. (B) Heat-map of different gene expression patterns in Hp-positive patients between group 1 (low expression) and group 2 (high expression). (C) Volcano diagram of differential genes in the two subgroups, and the 10 annotated genes with the most significant differences. (D) GO functions of underlying differential genes and the KEGG pathway enrichment.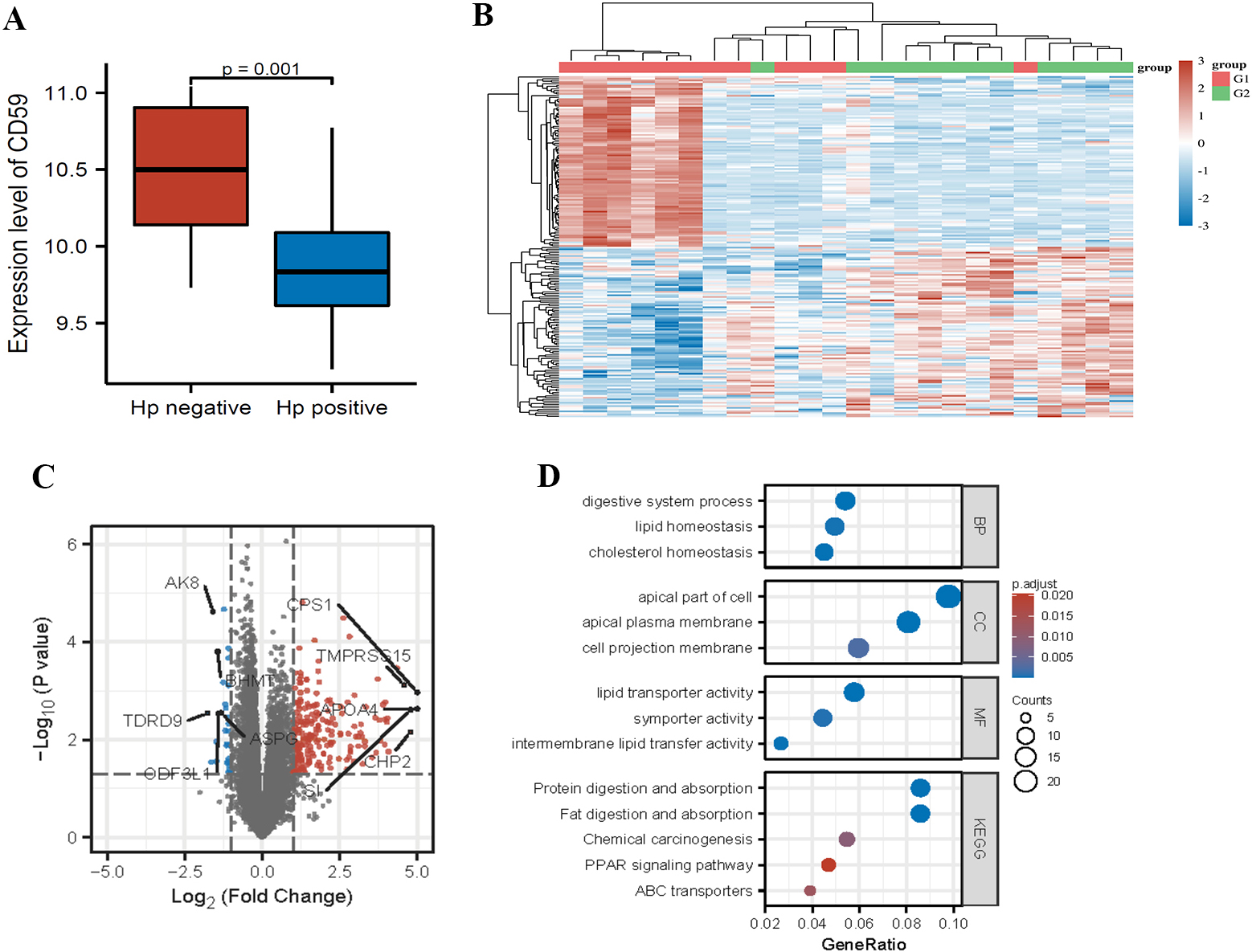
Function of CD59 in the occurrence and progression of malignant tumors. (A) Analysis of CD59 expression in various cancer tissues. (B and C) CD59 expression in STAD (Gastric cancer) project (unpaired or paired samples). (D–H) relationship between CD59 tumor overall survival in STAD.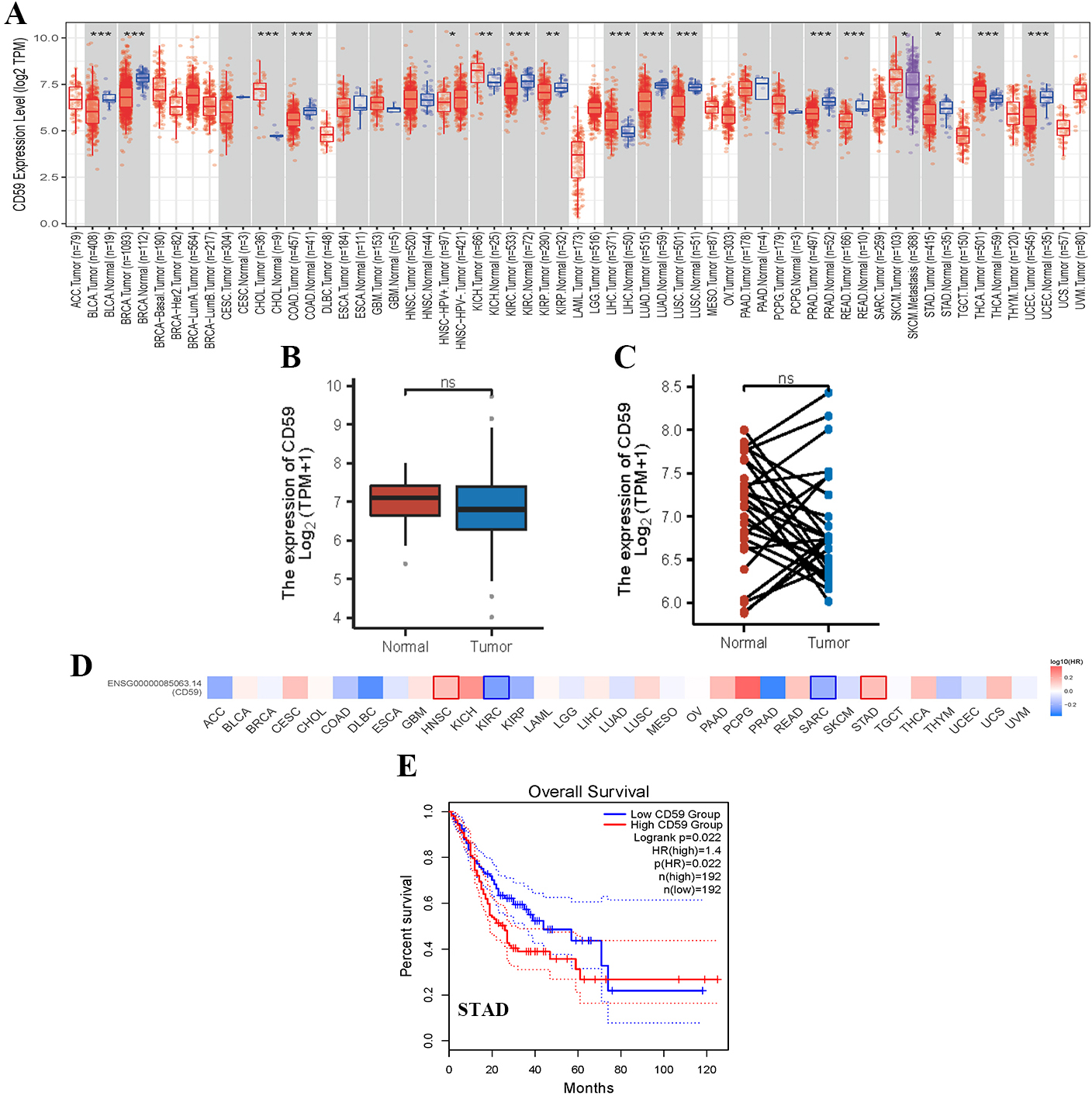
Possible role of CD59 in H. pylori infection and probable association with tumor immunity in gastric cancer. (A) Abundance difference of 24 immune cells predicted using the high and low CD59 expression subsets. (B) Relationship between CD59 gene and 60 immune checkpoint pathway genes, and expression data of marker genes in each sample as well as the correlation. (C) Significant enrichment of genes analyzed by GSEA. (D) Protein interactions predicted by String database, and CD59 with a complex interaction network of CD55, CD46, CD58, CD2 and other genes. (E) Mechanism of CD59 in gastric cancer reanalyzed by KEGG and GO analyses. (F) Expressions of the closely interacting proteins in GC, correlation analysis, and visualization in a heat map.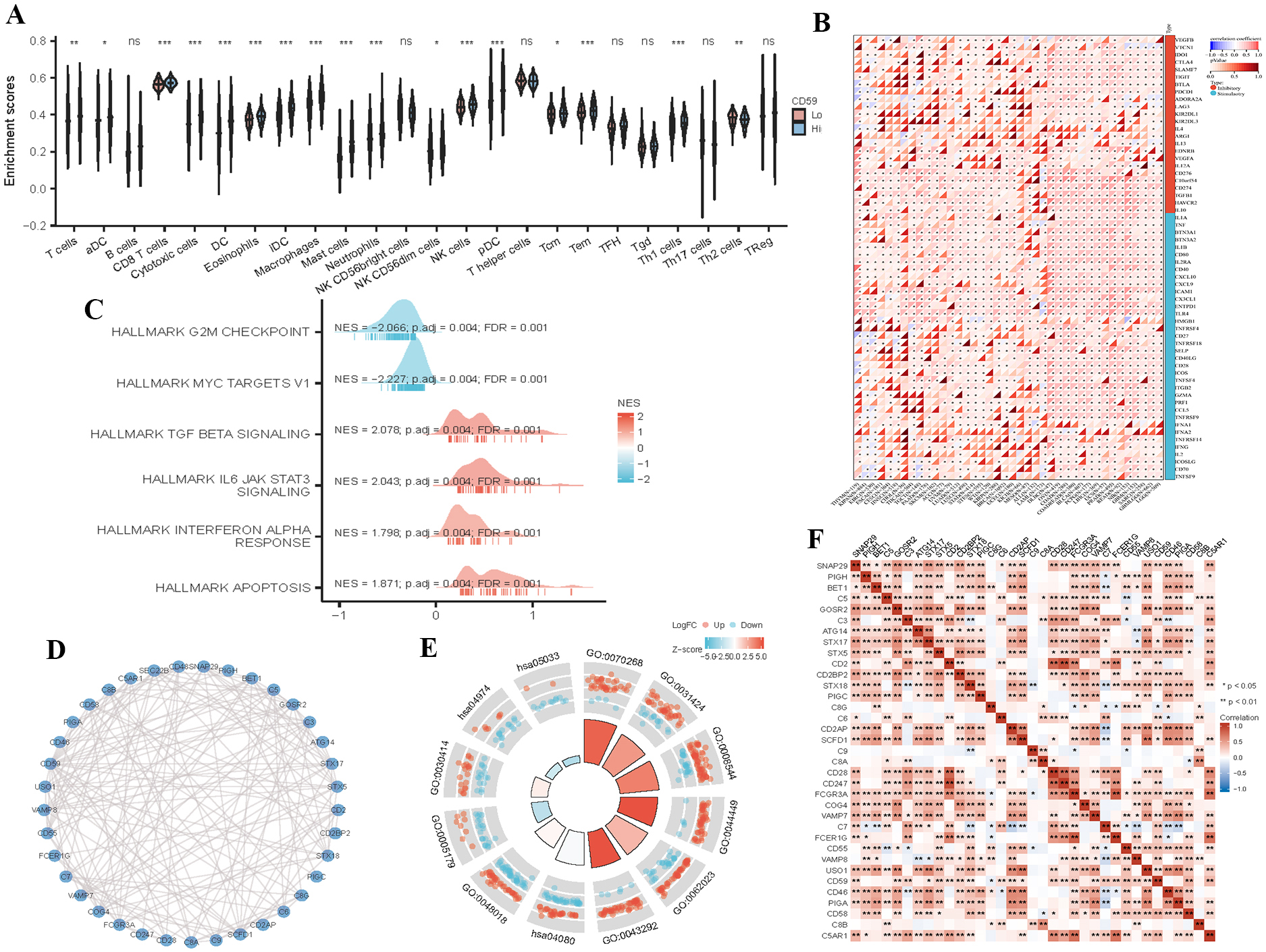
CD59 expressions in GC tissues or cell lines infected with H. pylori analyzed by Western blot. CD59 expressions in (A) GC tissues and GC tissues infected with H. pylori. (B) GES-1 cells and GES-1 cells infected with H. pylori. (C) AGS cells and AGS cells infected with H. pylori.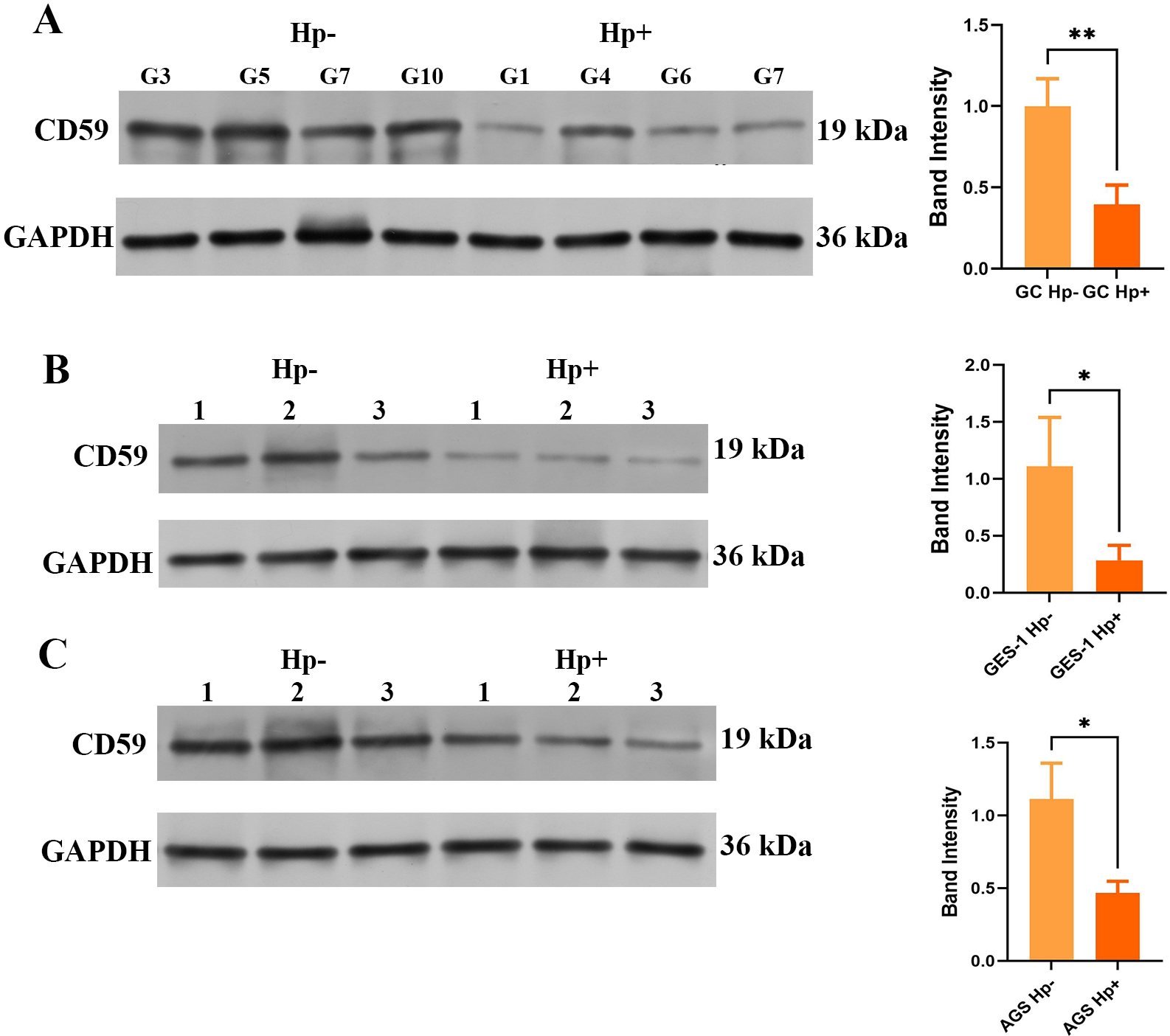
The CD59 gene expression decreased significantly in the H. pylori positive versus negative samples (Fig. 2A). To probe into the roles of CD59, we divided the H. pylori positive patients into two groups with low (group 1) or high (group 2) CD59 expression. The heat map of the differential genes was shown, and the two subgroups had different expression patterns (Fig. 2B). Meanwhile, ten genes with the most significant difference were labeled in the volcano map (Fig. 2C). The GO functions of the differential genes were analyzed, and KEGG pathway was enriched. BPs mainly involve digestive system process, lipid homeostasis and cholesterol (Fig. 2D). In terms of the CC, differential genes are engaged in the apical part of cell, apical plasma membrane, and cell projection membrane (Fig. 2D). As for MF, the differential genes participate in activities of lipid transporter, symporter, and intermembrane lipid transfer (Fig. 2D). The KEGG pathway analysis reveals that differential genes are enriched in protein/fat digestion and absorption, chemical carcinogenesis, and PPAR signaling pathway (Fig. 2D).
Bioinformatics analysis of CD59 in occurrence and development of GC
The CD59 expression decreased significantly in stomach adenocarcinoma (STAD) analyzed through TCGA (Fig. 3A). But unfortunately, there is no statistical difference of CD59 expression between unpaired and matched STAD samples (Fig. 3B and C). The relationship between CD59 and overall tumor survival was analyzed through Gene Expression Profiling Interactive Analysis 2 (GEPIA). It was found the survival rate of STAD patients with CD59 low expression is worse that with high expression (Fig. 3E).
Bioinformatics analysis of CD59 in H. pylori infection related to tumor immunity of GC
H. pylori in GC microenvironment regulated various immune cells, such as CD8
CD59 expression in H. pylori infected GC tissues or cell lines
According to bioinformatics analysis, CD59 is pivotal in H. pylori infection or GC. H. pylori infection inhibited CD59 expression in GC in vivo (Fig. 5A). In addition, the expression of CD59 decreased significantly in H. pylori infected GES-1 or AGS cells in vitro (Fig. 5B and C).
Discussion
H. pylori has been one of the most widespread bacterial infections globally, and causes chronic gastritis and increases the risk of peptic ulcer disease, gastric adenocarcinoma, and mucosa-associated lymphoid tissue lymphoma [19]. In this study, we analyzed the DEGs between H. pylori-infected GC cells relative to GC cells (GC-Hp and GC-Hp
Among these genes, CD59 was downexpressed in H. pylori-infected GC cells and attracted us. CD59 is recognized as a glycosylphosphatidylinositol-anchored membrane protein that inhibits the generation of the membrane attack complex to modulate complement stimulation [20]. At present, studies revealed that CD59 is highly expressed in various cancer cells or tissues, and mediates the action, infiltration and phenotypes of various immune cells in the tumor microenvironment [21, 22]. Thus, targeting at these biofunctions and pathways assists in elaborating the concrete mechanisms of GC. Additionally, the relationship between CD59 expression and H. pylori infection was analyzed with the GEO database. The biological functions of differential genes are enriched in protein and fat digestion/absorption, chemical carcinogenesis, and PPAR pathway. Our bioinformatics results show that the expression is closely related to various cancers. Among them, CD59 expression decreased significantly in H. pylori-infected STAD. In addition, the overall survival rate of STAD patients is closely related to the expression levels of CD59.
The expressions and functions of CD59 in immune cells were already studied, such as T lymphocytes, tumor-associated macrophages (TAM), and NK cells [23]. The CD59 molecule enhances the cytotoxicity of NK cells, so a decrease in the expression of CD59 in GC cells may weaken the CD59 mediated cytotoxicity of NK cells, thereby facilitating the escape of tumor cells from cellular immune response and leading to the occurrence of GC [24]. The functions of CD59 in H. pylori infection related to tumor immunity and GC were studied through bioinformatics. About 24 immune cells are closely related to CD59 expression such as T cells, CD8
The biological functions of CD59 are to prevent cancer cells from complement attack and to mediate the roles and phenotypes of immune cells. Nevertheless, CD59 shortage can initiate hemolytic dysfunction in autoimmune hemolytic anemia and paroxysmal nocturnal hemoglobinuria [26, 27]. Soluble and glycated CD59 is a novel biomarker for recognizing obstructive chronic lung disorders and diabetes [28, 29]. Cell experiments in vitro showed that H. pylori infection suppressed CD59 expression in GC cells and GC tissues.
This study demonstrates that H. pylori infection leads to CD59 downexpression in GC tissues and cell lines as verified in cell experiments in vitro. In the future, the function and mechanism of CD59 in H. pylori infected GC will be studied further.
Declaration of competing interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 82002111), the Medical Scientific Research Project of Jiangsu Provincial Health Commission (No. Z2019001), and the Young Scientists Foundation of Changzhou No. 2 People’s Hospital (NO. 2022K006).
Author contributions
Conception: Jianjun Wang, Hong Zhu.
Interpretation or analysis of data: Jun Sun.
Preparation of the manuscript: Hui Cao.
Revision for important intellectual content: Tingting Wen, Zi Xu.
Supervision: Xian Zhang.
Footnotes
Acknowledgments
All authors acknowledge her immense support in the grammatical review of this manuscript.