Over the past decade, cancer immunotherapy, such as immune checkpoint inhibitors (ICRs), has attained considerable progresses in clinical practice. T-cell immunoglobulin and mucin domain-containing protein 3 (Tim-3) act as next ICRs, and originally function as a co-inhibitory receptor expressed on interferon (IFN)- producing CD4 and CD8 T-cells. Furthermore, Tim-3 has also been found to express on innate immune cells and several types of tumors, signifying the pivotal role that Tim-3 plays in chronic viral infections and cancer. In addition, Tim-3 and multiple ICRs are concurrently expressed and regulated on dysfunctional or exhausted T-cells, leading to improved antitumor immune responses in preclinical or clinical cancer therapy through co-blockade of Tim-3 and other ICRs such as programmed cell death-1 (PD-1). In this review, the biological characteristics of Tim-3 and the function of Tim-3 in regulating tumorigenesis and inflammation have been summarized. The usage of a single blockade of Tim-3 or in combination with multiple immunotherapy regimens have drawn attention to antitumor potential as a target for immunotherapy.
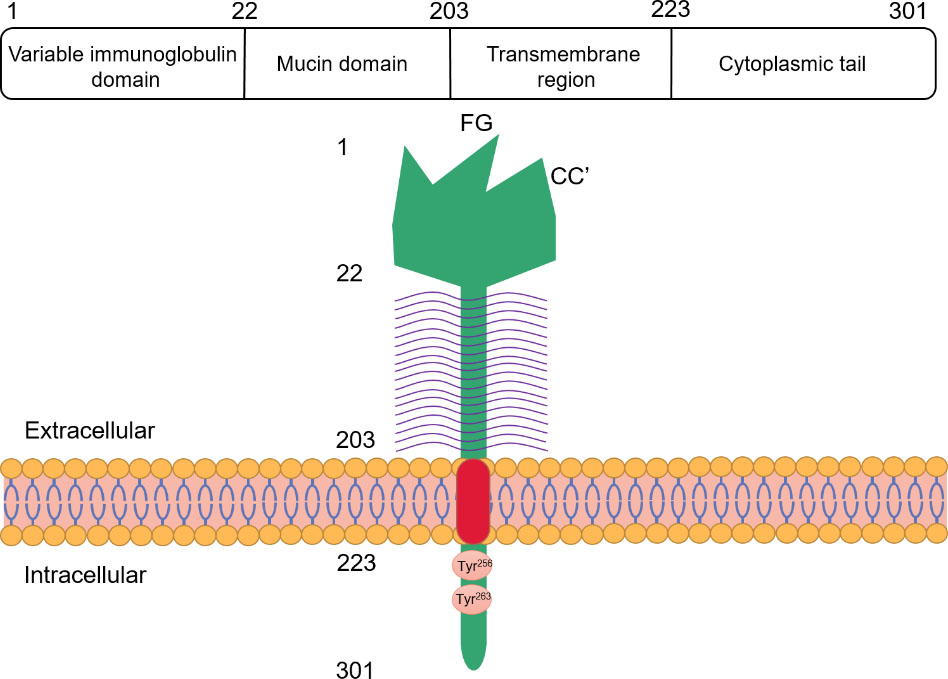
Molecular structure of Tim-3. The human Tim-3 is located on chromosome 5, with a total length of 301 amino acids. The structure of Tim-3 protein consists of four distinct regions: variable immunoglobulin domain (IgV), mucin domain, transmembrane region and cytoplasmic tail. In the IgV domain of Tim-3, due to the existence of disulfide bonds, compared with the traditional IgV domain, the distance between FG loop and CC’ loop is smaller. The mucin domain of Tim-3 contains N-linked glycosylation sites.
Introduction
Immunotherapy is being increasingly considered as a vital part of cancer therapy. It has been evidenced to enhance the human immune response to cancer cells, hinders tumor growth, and has attained remarkable success in the field of cancer therapy. The treatment by targeting immune checkpoint inhibitors (ICRs), such as programmed cell death-1 (PD-1) and cytotoxic T-lymphocyte antigen-4 (CTLA-4), has been successfully accomplished in kidney cancer, melanoma, and lung cancer. Nevertheless, due to a poor response rate of certain cancer treatments, it is necessary to seek other co-inhibitory receptors or ICRs to enhance the outcome of immunotherapy ineffective or refractory to anti-CTLA-4 and anti-PD-1 therapy. Tim-3 is a member of the T-cell immunoglobulin and mucin domain-containing protein 3 (TIM) family. Tim-3 is a co-inhibitory receptor, generally expressed in CD4 T, CD8 T, dendritic, and macrophagic cells that secrete IFN-. The interaction between Tim-3 and its ligands has been proved to induce a negative regulatory pathway of the normal immune response, causing amplified autoimmunity and loss of tolerance and rendering Tim-3 a negative regulatory molecule. These immune tolerance mechanisms are part of the latest immunotherapy, which work by blocking immune checkpoints to annul the immune response of functional inhibition, activate T-cells, and eventually promote antitumor immunity. Tim-3 is one such prospective receptor for tumor immunotherapy. Blocking Tim-3 using the checkpoint inhibitors is known to augment antitumor immunity and inhibit tumor growth by several studies. Sun et al. [1] have shown that intravenous injection of oncolytic vaccinia virus (oVV) has a synergistic antitumor immunotherapy effect with both PD-1 and Tim-3 blockade in a mouse lung cancer model, rather than blocking single PD-1 or Tim-3. Studies have shown that Tim-3 combined with PD-1 blocking may help to prevent the depletion of CD8 T cells in patients with advanced acute myeloid leukemia (AML) and other malignant hematological tumors [2]. Even though, Tim-3 has been shown to play a certain role in the occurrence and development of liver cancer [3], colon cancer [4]. and so on. In order to gain an in-depth understanding of Tim-3 as a potential immune target, the biological characteristics of Tim-3 have been reviewed initially, and its role in regulating tumorigenesis, tissue inflammation, and antitumor immunity is detailed next, followed by its usage as a target for cancer therapy.
Expression and function of Tim-3 on cancer cells
Cell type
Expression or function
Reference
Acute myelocytic leukemia
Tim-3 combined with PD-1 blocking may help to prevent CD8 T cell exhaustion in patients with advanced acute myelocytic leukemia (AML) and other malignant hematological tumors.
Tim-3 was expressed in all prostate cancer cell lines, and knock-down expression inhibited the proliferation and invasion of prostate cancer cell lines.
Tim-3 was over-expressed in bladder cancer cells, tumor infiltrating lymphocytes (TIL) and endothelial cells from patients with bladder urothelial carcinoma (BUC).
Both Gal-9 and Tim-3 are expressed in tumor cells of gastric cancer patients, which may be potential and independent prognostic factors in gastric cancer patients. Gal-9 and Tim-3 may also play an important role in the development of gastric cancer.
Otherwise known as hepatitis a virus cellular receptor-2 (Havcr-2), Tim-3 is a member of the TIM family [5]. Whereas eight TIM genes (Tim-1–Tim-8) have been found in mice, only three genes encoding Tim-1 (Havcr-1), Tim-3 (Havcr-2), and Tim-4 (Timd4) have been identified in humans [6]. Tim-3’s structure has four distinct regions, namely, variable immunoglobulin domain (IgV), mucin domain, a transmembrane region, and the cytoplasmic tail [7]. The IgV domain includes two antiparallel chains, N-linked glycosylation sites, and four cysteines [8]. As against the traditional IgV domain, the Tim-3 IgV domain consists of two disulfide bonds in the four cysteines conserved by TIM protein, which repositions the CC’ loop closer to the FG loop, forming a unique cleft in the IgV domain of the family of proteins [9] (Fig. 1). Among the TIM family, Tim-3 is the molecule with the smallest mucin domain, containing O-linked glycosylation sites, especially rich in serine, proline, and threonine [9]. A tail, mainly composed of hydrophobic amino acids and embedded in a lipid bilayer, is utilized by Tim-3 proteins to pass through the cell membrane, which can penetrate into the cell with a cytoplasmic domain of 42–77 amino acids. Tim-3 also occurs in a soluble form (sTim-3), which, as compared with the complete Tim-3 structure, lacks mucin and transmembrane domain [10].
Earlier studies recognized Tim-3 as a specific membrane marker for terminal differentiation of helper T-cell 1 (Th1) but is not expressed on helper T-cell (Th2) [11]. Tim-3 has also been indicated by recent research to be expressed on pro-inflammatory T-cell subsets of Th17 cells, although the level of expression is lower than that of Th1 cells [12, 13]. Tim-3 was later found to be not just expressed on cells of the acquired immune response. Tim-3 has also been reported to be expressed by other cell subsets, including human monocytes [14], macrophages [15], natural killer cells [16], dendritic cells [17], mouse dendritic cells [18], mast cells [19], macrophages [20], and even in certain tumor cells. These results indicate that Tim-3 not only negatively regulates inflammatory T-cell function but also essentially participates in inhibiting autoimmune response [21], besides supporting the formation of an immunosuppressive tumor environment [21]. Depending on the pedigree transcription factor T-BET, Tim-3 is strongly expressed in differentiated Th1 cells. T-BET encourages the expression of Tim-3 by directly binding to the Tim-3 promoter [22]. Second, immunosuppressive cytokine IL-27 is a T-cell depletion regulator, which stimulates suppressor gene modules that cohere to Tim-3 and other ICRs such as TIGIT (T cell immunoglobulin and ITIM domain), Lag-3 (Lymphocyte activation gene-3), and PD-1 [23]. The transcription factor NFIL3 can also be upregulated by IL-27 to directly trigger the expression of Tim-3, whereas NFIL3 and T-BET together reshape the chromatin structure of the Tim-3 promoter for transcription [24].
To date several Tim-3 ligands have been discovered. The first identified ligand of Tim-3 is Galectin-9 (Gal-9), which belongs the highly conserved galactose lectin family [25]. It suggests the inhibitory function of Tim-3, as Gal-9 particularly recognizes Tim-3 of Th1 cells and induces apoptosis, thereby hindering tissue inflammation and autoimmune disease EAE [26]. Gal-9 has been identified to be expressed in autoimmune diseases, tumor microenvironment, chronic infection tissues, and various other cells [26]. Phosphatidylserine (PtdSer) is an essential phospholipid found in the membranes of bacteria, yeast, plants, and mammalian cells [27]. It can bind human and mouse Tim-3 in the FG-CC’ loop of the IgV domain. As there is an FG-CC’ loop in the TIM family, PtdSer can also act as a ligand for other TIM family proteins [9]. Thus, PtdSer may be the key ligand of Tim-3 on macrophages or dendritic cells and their interaction could be essentially responsible for initiating inhibitory signals without prompting phagocytosis of apoptotic cells. Another ligand of Tim-3 is CEACAM-1. It is found to be expressed on T-cells and by way of the interaction of IgV domains on two molecules, binds to Tim-3 [28]. The negative regulatory function of Tim-3 has been shown to be defective in the absence of CEACAM-1, suggesting CEACAM-1 is essential for Tim-3 to function normally, and that CEACAM-1 enhances the expression on the surface of Tim-3 cells [28]. Another Tim-3 ligand is HMGB1 that influences innate immune response [29]. HMGB1 chiefly encourages the functioning of congenital cells by cohering to advanced glycation end products receptor (RAGE) and Toll-like receptor (TLR), in turn activating congenital cells and producing pro-inflammatory factors. Nevertheless, Tim-3, together with HMGB1, interferes with the above process, thus suppressing the activation of the innate immune response.
Roles of TIM-3 in cancer
Tim-3 and tumorigenesis
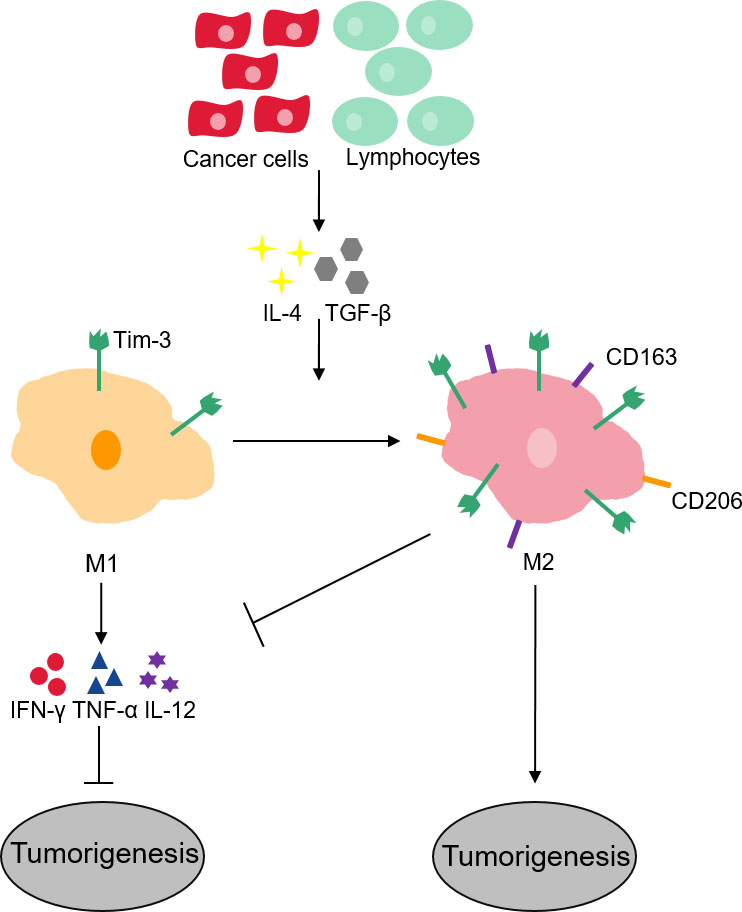
A variety of cancer cells such as myeloid leukemia [2], colon cancer [4], renal cell carcinoma [30], cervical cancer [31], lung cancer [32], prostate cancer [33], bladder urothelial carcinoma [34]. and gastric cancer [35]. have evidenced the expression of Tim-3 (Table 1). The amplified expression of Tim-3 is linked to disease progression and shorter survival in these cancers. A high proportion of Tim-3-expressing stem cells have been observed by certain investigations in human hepatocellular carcinoma tissue, supporting tumor cell growth [36], whereas anti-Tim-3 antibody or Tim-3 knockout mice suppresses tumor growth. Tim-3 receptor actually activates NF-B phosphorylation in hepatocytes, which eventually triggers IL-6 secretion and STAT3 phosphorylation [37]. The inherent function of intratumoral Tim-3 in tumorigenesis is thus confirmed, suggesting that Tim-3–mediated immune depletion in the tumor microenvironment will likely become a new target for tumor therapy. It has been proved that Tim-3 plays an important role in the occurrence and development of hepatocellular carcinoma. At the same time, Tim-3 can be used as a target to evaluate the prognosis and therapeutic effects of liver cancer. Tim-3 intervention has shown an anti-tumor effect in preclinical trials [38]. In terms of regulatory mechanism, phosphatidylserine (PtdSer) is involved in promoting the phosphorylation of Tim-3, and then competes with PI3K p110 to bind p85 to inhibit downstream Akt/mTORC1 signal, resulting in dysfunction of two NK cell subsets (conventional NK cells and liver-resident NK cells). Tim-3 promotes the growth of liver cancer by blocking the function of NK cells, which provides a new strategy for targeted therapy based on immune checkpoints [3]. Considering the interaction between Tim-3 and its ligands described hereinbefore induces T-cell apoptosis and functional inhibition, increasing evidence indicates that Tim-3 negatively regulates antitumor immunity. Therefore, Tim-3 appears to have a certain promoting effect on tumorigenesis. The total number of tumor antigen-specific CD8 T-cells increases when anti-Tim-3 and anti-PD-L1 antibodies are utilized to block Tim-3 and PD-1 signaling pathways, which considerably inhibits tumor growth. Tim-3 is a prospective biomarker for tumor immunotherapy. Tim-3’s expression in M1 macrophages is low and executes the role of phagocytosis, antigen presentation, and production of pro-inflammatory cytokines such as IFN-, TNF-, and IL-12 [39]. When anti-inflammatory cytokines such as IL-4 and TGF- are released into the environment by cancer cells and lymphocytes, they induce M1 phenotype to the M2 as they lose or reduce several protective functions in cancer, therefore inducing tumor development and progression, besides increased expression of Tim-3 and that of specific markers of M2 phenotype such as CD163 and CD206 (Fig. 2) [40]. Simultaneously, these outcomes also indicate that depleted T lymphocytes may be advantageous to tumor development. Moreover, Tim-3 has also been evidenced to promote the progress of tumor metastasis through another unknown mechanism, such as in HeLa cells, following the silencing of Tim-3 by antisense cDNA, the metastatic ability of the HeLa cell line is decreased [31].
Tim-3 macrophages and its role in tumorigenesis. In M1 macrophages with low expression of Tim-3, which can promote phagocytosis, antigen presentation and product pro-inflammatory cytokines such as IFN-, TNF-, and IL-12. And these pro-inflammatory cytokines can active anti-tumor response. Then as cancer cells and lymphocytes release anti-inflammatory cytokines into the environment, macrophages are induced to transition to tolerance profile (M2), they lose several protective functions in cancer. In M2-like macrophages with high expression of Tim-3, CD163 and CD206are conducive to tumor development and progression.
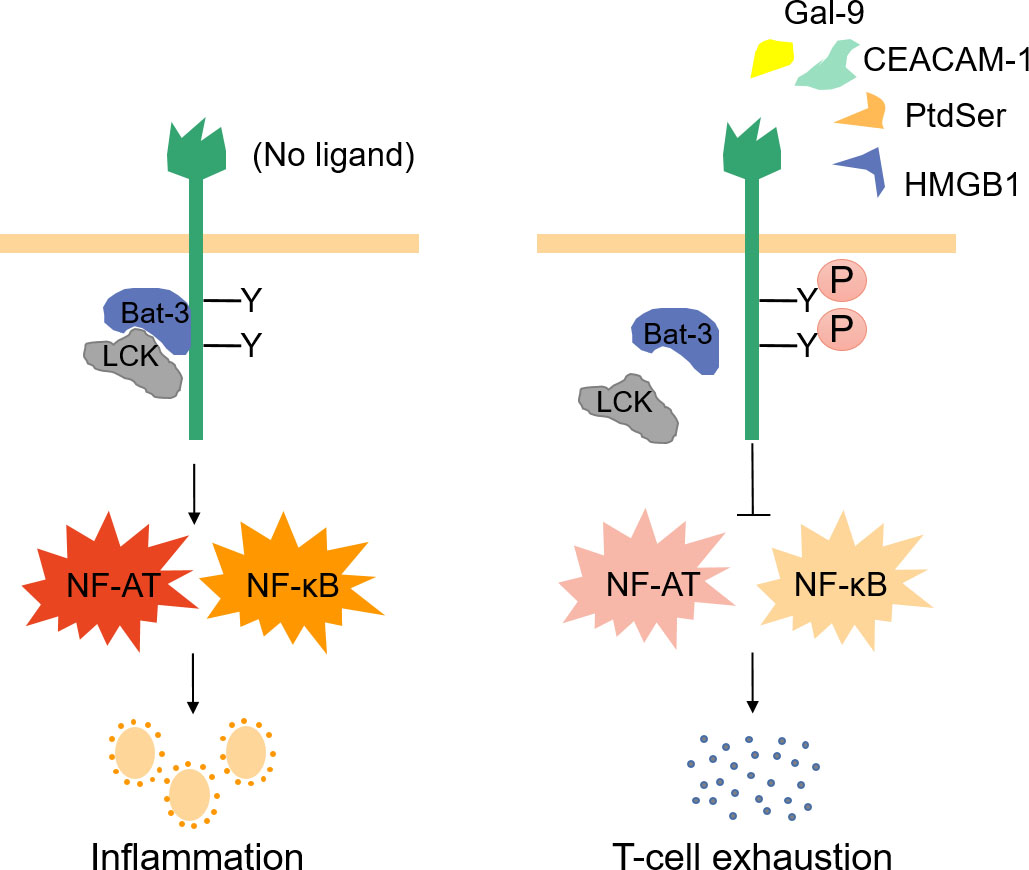
The role of Tim-3 in inflammation. In the absence of ligand interaction, molecular junction Bat-3 and Src family kinase LCK bind to the tail of Tim-3 and activate NF-AT and NF-B pathways through Tim-3 signaling pathways, thus promoting T cell activation and inflammation. Tim-3 ligand interacts with it to phosphorylate the key tyrosine residues in the tail of Tim-3, thus destroying the binding of Tim-3, Bat3 and LCK, resulting in T cell exhaustion, which is conducive to tumor development..
Tim-3 and tissue inflammation
The inflammatory response is controlled by several pathways, usually involving CD4+T helper cell 1 (Th1). Th1, an inflammatory mediator, is regulated by various factors besides immune negative receptor Tim-3 [41]. The interaction between Tim-3 and its ligands significantly regulates Th1 response and promotes T-cell tolerance, suggesting Tim-3’s essential role in the pathogenesis of human autoimmune diseases involved in the imbalance of Th1 immune response [26]. Tim-3 has been reported to terminate Th1 immunity and influence a series of inflammatory responses by way of differential expression of innate and acquired immune cells [17]. Tim-3 encourages inflammation in a naïve resting immune system. Tim-3 is primarily expressed in naïve DC cells and cooperates with TLRs to support the secretion of TNF-. In this perspective, Tim-3 is highly expressed in terminally differentiated Th1 cells following the Th1 response, which is higher than that in DCs, and upregulation of Gal-9 is induced by the production of IFN-. Simultaneously, Tim-3 interacts with Gal-9 and eventually ceases the cellular immunity of Th1 [42]. In the absence of Tim-3 and its ligand interaction, the molecular junction of human leukocyte antigen-B–associated transcript 3 (Bat-3) and Src family kinase LCK cohere to the tail of Tim-3 and trigger NF-AT and NF-B pathways via Tim-3 signaling pathways, thereby promoting T-cell activation and inflammation [43]. The interaction between Tim-3 and ligands phosphorylates the crucial tyrosine residues in the Tim-3 tail, consequently destroying the binding of Tim-3, Bat3 and LCK, terminating the NF-AT and NF-B pathways, leading to T-cell exhaustion (Fig. 3) [44]. These studies indicate that single-molecule Tim-3 performs an opposite role in innate and acquired immune systems, which not only promotes the inflammatory response but also terminates Th1 immunity. To summarize, Tim-3 may turn out to be an effective therapeutic target for peripheral and organ-specific human inflammatory diseases.
Tim-3 as a novel ICR
ICRs have a vital role in immune regulation, such as immune tolerance, immune homeostasis, and tumor immune escape. ICRs are mainly expressed on T cells on immune activation to reconcile immune response and sustain homeostasis. Lack of ICRs on T cells may cause autoimmunity diseases, whereas their expression on tumor-infiltrating T cells may make tumors to evade immune elimination [45]. In addition to two earliest best-described ICRs, namely, PD-1 and CTLA-4, several others have been successively recognized till date, which include Tim-3, Lymphocyte activation gene-3 (Lag-3) T-cell immunoglobulin and ITIM domain (TIGIT), and CD96. Of them, PD-1 and CTLA-4 were extensively studied and broadly expressed on activated T-cell, which play key roles in inhibiting T-cell–mediated immune response [46]. Therefore, both PD-1 and CTLA-4 have been practiced as therapeutic targeting for aggressive, metastatic, and resistant solid cancers.
As a next-generation ICR molecule, the role of Tim-3 in immune control has attained high popularity. Expression of Tim-3 has been evidenced by earlier studies to be largely restricted in T cells, including inflammatory IFN--producing type-1 CD4 T-cells, CD8 T-cells, NK T cells, and FoxP3 Treg cells [21, 47]; therefore, it can maintain the physiological immune balance. In addition, high expression of Tim-3 was also found on exhausted T cells and led to cancer progress and immune evasion with dysfunctional immune response [48]. Tim-3 has also been found in recent years to be expressed by macrophages [49], NK cells [16], and mast cells [19], which allows therapeutic targeting for multiple disease types through modulating immune responses.
Mechanistically, the primary intracellular signaling of Tim-3 was linked to the interaction between its cytoplasmic tail tyrosine Tyr and Tyr and Bat-3 [44], wherein Bat-3 promotes T-cell responses and autoimmunity by repressing Tim-3–mediated cell death and exhaustion. On phosphorylation of both Tyr and Tyr by the tyrosine kinase ITK following Tim-3 binding with its ligands Gal-9 and CEACAM1, Bat-3 was discharged from Tim-3 at Tyr and Tyr [28, 43], which stimulates Tim-3 to exert its inhibitory function. Rangachari et al. [44] further testified that Bat3-deficient T cells have a higher expression of co-inhibitory receptors, including Tim-3, and distinctly inhibit the autoimmune encephalomyelitis, suggesting that Bat-3 could act as an inhibitor of Tim-3–mediated T-cell exhaustion. Another prospective inhibitor of Tim-3 is FYN, which can competitively bind Tyr and Tyr of Tim-3 with Bat-3 to trigger Tim-3–mediated immune inhibitory signaling [50]. Also, Tim-3 can be recruited to the immune synapse on CD8 T-cell activation, which spoils a stable synapse formation, and interacts with receptor phosphatases CD45 and CD148 [51], signifying a different mechanism of Tim-3–mediated immune dysfunction and exhaustion. Therefore, Tim-3 performs the immunological inhibitory function, a novel ICR through multiple intracellular signaling mechanisms.
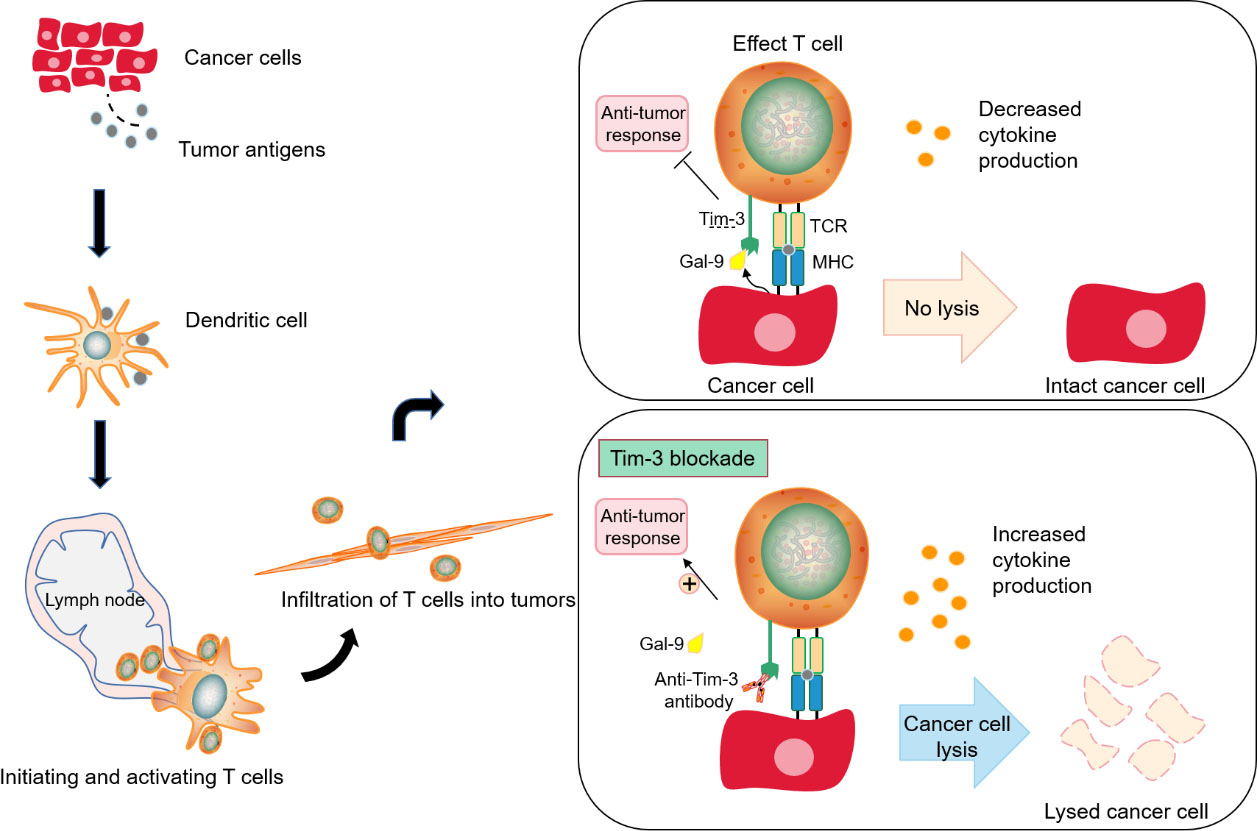
Blocking Tim-3 to promote anti-tumor immunotherapy. The production of immunity to cancer is a cyclic process, from the production of tumor antigen to the capture and transmission of tumor antigen by dendritic cells (antigen presenting cells, APCs) to T cells, which are bound by the specific receptor TCR of T cells, and interact directly to kill cancer cells, thus releasing new related antigens. However, if the co-inhibitory receptor TIM-3 on T cells binds to the ligands of inhibitory receptors produced by cancer cells (A), cytokines will decrease, resulting in a down-regulation of T cell function, which essentially forms a negative feedback cycle that suppresses anti-tumor immunity. At this time, blocking Tim-3, can restore the function of T cell effect and lysed tumor cells and enhance anti-tumor immunity.
Nevertheless, unlike classic ICRs such as PD-1, CTLA-4, and TIGIT, Tim-3 lacks a definable inhibitory signaling motif, and encloses five tyrosines in its cytoplasmic tail at the intracellular region, indicating that Tim-3 may play either immunosuppressive or stimulatory function by way of incompletely elucidated signaling mechanism [45]. This argument was supported by at least studies. Lee et al. observed that ectopic expression of Tim-3 in T-cells promotes activation of TCR signaling pathways, including the intensification of the level of transcriptional reporters and endogenous cytokine production with short-term stimulation [50]. Avery et al. evidenced that Tim-3 co-stimulation hastens short-lived effector T-cells by way of enhanced AKT-mTOR signal pathway [52]. Therefore, exhaustive investigations to describe the roles of Tim-3 for either co-inhibitory or co-stimulatory receptors in human diseases, including cancers, are imperative.
TIM-3 as a target in cancer immunotherapy
A pivotal emerging hallmark of cancer is evasion of immunological destruction, in particular by T cell dysfunction or T cell exhaustion. Aiming at therapeutic targeting, the breakthrough drugs have been increasingly developed including anti-ICRs mAb, CAR-T, and oncolytic virus that are in clinical trials or approved for treating certain types of human cancer.
Tim-3 can act as an attractive clinical target for checkpoint-targeted cancer immunotherapy. Consistent with the key regulatory effects of innate and acquired immune responses, blocking Tim-3 plays a positive role in restoring anti-tumor immunity (Fig. 4). However, Tim-3 may play a more complex role in regulating anti-tumor response. For instance, Tim-3 blockade by anti-Tim3 antibody reduced both population of CD4 CD25 Foxp3 Treg and Tim-3 Treg cells, drived antitumor immune response and suppressed tumor growth in head and neck cancer with mice xenograft [53, 54]. Tim-3 target is an effective target for tumor immunotherapy, and its expression is related to the poor prognosis of tumor and drug resistance of PD-1. Anti-Tim-3 antibody has a synergistic effect with CTLA-4, PD-1 and PD-L1 antibodies, and can treat patients with drug resistance to PD-1 and PD-L1. However, single Tim-3 blockade by antibody does not achieve a substantial therapeutic benefit, and the principle of the limitations of single blocking Tim-3 is not known.
The increasing data showed that there exist a common interaction between Tim-3 and other ICRs to regulate T cell function, such as frequent Tim-3 overexpression on tumor-infiltrating CD8 PD-1 T cells in various types of cancers [55, 56, 57]. Thus, it provides the basis of combination therapy with Tim-3 blockade and PD-1 blockade. The study showed that targeting both Tim-3 and PD-1 with their antibodies reversed T cell exhaustion and exerted a synergistic effect in inhibiting tumor growth and restoring antitumor immunity [56]. In addition, the combination blockade of Tim-3 and PD-1 reversed tumor-induced T cell exhaustion, and improved cancer antigen specific CD8 T cell responses in melanoma [57, 58]. Co-blockade of Tim-3 and PD-1 also obtained stronger antitumor immune responses in colon cancer and AML mouse models than that of PD-1 blockade alone [2, 59, 60]. Moreover, co-targeting of Tim-3 and CTLA-4 with blocking antibodies was showed to be well-tolerated and to be more effective in T cell IFN--mediated antitumor immunity and inhibition of established tumor growth compared with single blockade [59].
Clinical trials with anti-Tim-3 drugs in cancer
Drug name
Developer
Co-interventions
Phase
Cancer type
NCT number
Sym023
SymphogenA/S
Monotherapy
I
Metastatic cancer, Solid tumor and lymphoma
NCT03489343
Sym023
SymphogenA/S
Sym021 (Anti-PD1) and Sym022 (Anti-LAG3)
I
Metastatic Cancer, Solid Tumor and Lymphoma
NCT03311412
MBG453
Novartis Pharmaceuticals
PDR001 (Anti-PD1), SRS
I
Glioblastoma Multiforme
NCT03961971
MBG453
Novartis Pharmaceuticals
Azacitidine
III
Myelodysplastic Syndromes and Leukemia, Myelomonocytic, Chronic
NCT04266301
MBG453
Novartis Pharmaceuticals
Hypomethylating agents
II
Myelodysplastic Syndromes
NCT03946670
MBG453
Novartis Pharmaceuticals
HDM201
I
Acute Myeloid Leukemia (AML) and High-risk Myelodysplastic Syndrome (MDS)
NCT03940352
MBG453
Novartis Pharmaceuticals
PDR001 (Anti-PD1) and Decitabine
I, II
Advanced Malignancies
NCT02608268
INCAGN02390
Incyte
INCAGN02385 (Anti-LAG3), INCMGA00012 (Anti-PD1)
I/II
Melanoma
NCT04370704
INCAGN02390
Incyte
Monotherapy
I
Cervical Cancer, Gastric Cancer, Stomach Cancer, Gastroesophageal Junction Cancer
Adult Primary Liver Cancer (PLC), Advanced Adult PLC and Localized Unresectable Adult PLC
NCT03680508
RO7121661
Roche
Monotherapy
I
Solid Tumors, Metastatic Melanoma, NSCLC, SCLC and ESCC
NCT03708328
LY3321367
Eli Lilly and Company
LY3300054 (Anti-PDL1)
I
Solid Tumor
NCT03099109
BGB-A425
BeiGene
Tislelizumab (Anti-PD1)
I, II
Locally Advanced or Metastatic Solid Tumors
NCT03744468
LY3415244
Eli Lilly and Company
Monotherapy
I
Solid Tumor
NCT03752177
To be noted, Tim-3 expression is limited on terminally differentiated IFN--producing T cells and intratumoral Treg cells, whereas CTLA-4 and PD-1 are broadly expressed on activated-T cells, and expression of CTLA-4 exists on all Treg cells [45]. Moreover, Tim-3-deficient mice didn’t develop experimental allergic asthma [61], whereas both PD-1-deficient and CTLA-4-deficient mice develop autoimmune disease [62, 63]. Thus, Tim-3 blockade is likely to avoid risk of toxic effects that occurred in PD-1 or CTLA-4 blockade, and the advantage of co-blockade of Tim-3 and PD-1 or CTLA-4 may be safer and more effective than that of co-blockade of PD-1 and CTLA-4 in targeting intratumoral T cells.
Recently, combinational strategy of ICR blockade therapy and oncolytic virus attracted more and more attention, and achieved satisfactory results in clinical trials for several solid tumors through the hypothesis that oncolytic virus can turn cold (such as refractory PD-1 Tim3) cancers into hot (PD-1 Tim3) ones allowing ICR blockade antibody to treat better [64]. It can be verified by Sun et al. [1] that systemically administered oncolytic vaccinia virus exerts synergetic antitumor efficacy with co-blockade of PD-1 and Tim-3 in refractory lung cancer through direct cell killing effect, recruitment of TILs and enhanced expression of PD-1 and Tim-3. In addition, we combine oncolytic adenovirus and Tim-3 blocking antibody to conduct therapeutic experiments, and found this combination could efficiently suppress tumor cell growth in liver cancer (unpublished data), indicating possible prospect for other cold solid cancer therapy.
Recent studies showed that high tumor-infiltrating CD39 CD8 T cells has reduced secretion of TNF- and IFN- along with enhanced expression of Tim-3 that decides poor prognosis in renal carcinoma [65], indicating that the ectonucleotidase CD39 may be acted as a novel combinational therapy target with anti-Tim-3 antibodies. Actually, it was early reported that co-blockade of Tim-3 with a blocking antibody and CD39 by the small molecule inhibitor POM-1 significantly inhibit melanoma growth in mouse xenograft model [66]. Moreover, some studies also have tried to use shRNA/siRNA to interfere Tim-3 expression, and showed that siTim-3 could efficiently suppress tumor growth and enhanced antitumor immune effect [39, 67], suggesting an alternative therapeutic approach targeting Tim-3. Hepatocellular carcinoma (HCC) is still one of the types of tumors with the worst prognosis, as a cancer with few systematic treatment, sorafenib is approved to be used in the treatment of liver cancer but can not eradicate liver cancer. Recent studies have shown that it is safe and effective to use cabozantinib, ramucirumab or metronomic capecitabine in the treatment of liver cancer in patients with HCC who are ineffective in first-line sorafinib failure [68, 69]. In the clinical treatment of hepatocellular carcinoma, it is expected that Tim-3 combined with chemotherapeutic drugs can further improve the therapeutic efficiency of hepatocellular carcinoma.
From bench to bedside, it is indispensable to further investigate the clinical effect of anti-Tim3 blocking antibodies due to its positive role of Tim-3 blockade in cancer therapy, especially the co-blockade of Tim-3 and PD-1 with the antibodies or oncolytic virus. Up to date, there are several ongoing and planned chinical trials with anti-Tim-3 blocking antibodies alone or in combination with anti-PD-1, PD-L1 or LAG-3 antibodies et al (Table 2). However, a single anti-Tim-3 mAb agent didn’t strongly elicit activating immune responses and significant tumor regression. It might be caused by which Tim-3 is co-expressed with other ICRs such as PD-1 on CD4 T and CD8 T cells, and its effect of blocking Tim-3 alone or PD-1 alone is limited. In addition, as immunosuppressive cytokine, IL-27 might play pivotal roles in driving ICRs expression including Tim-3, PD-1, TIGIT, and LAG-3 on CD4 or CD8 T cells [23, 24]. Thus, most clinical trials were performed by co-blockade of Tim-3 and PD-1 using their blocking antibodies. TSR-022, the first reported humanized anti-Tim3 blocking antibody by Tesaro company [70], was used in a phase I trial for patients with advanced solid tumors by single agent treatment and had no dose-limiting toxicity [71]. Furthermore, TSR-022 in combination with TSR-042 (anti-PD-1) exerted increased clinical effect in patients with colorectal cancer, post-PD-1 NSCLC and melanoma without dose-escalated toxicity [72]. Another two developed Tim-3 blocking antibodies LY3321367 [73] and MGB453 [74] derived from Eli Lilly and Company and Novartis, respectively, were proved to be interim safe, efficient and well-tolerated when combined with anti-PD-1 in a phase I trial in advanced cancers. More development of anti-Tim3 blocking antibodies, such as M6903 [75], are ongoing for pre-clinical test in worldwide (Table 2). These data emphasize the advantage of anti-Tim3 antibodies on single safety, superior efficacy and favorable pharmacokinetics with co-blockade of PD-1 pathway in clinical trials.
Conclusion
The complex biological characteristic endows the diverse functions of Tim-3 linked to tumorigenesis, immune regulation, chronic viral infections, and inflammation. In addition to its expression on T cells, Tim-3 is expressed by dendritic cells, macrophages, mast cells, NK cells, and myeloid-derived suppressor cells.
Furthermore, the role of Tim-3 in these non-T immune cells needs to be investigated. Besides, Tim-3 has no inhibitory signaling motifs in its structure, unlike other ICRs, provoking the debate whether Tim-3 functions as an inhibitory receptor or a stimulatory receptor. Another concern is that raised plasma sTim-3 level is linked to patients with membranous nephropathy [76], GVHD, and diverse malignancies, indicating that sTim-3 may serve as a diagnostic biomarker. Furthermore, sTim-3 may also function as a molecular sink to hinder the interaction of Tim-3 and its ligands. Nevertheless, the exact function of sTim-3 is yet to be studied.
Remarkably, blockade of the Tim-3 pathway by anti-Tim3 antibodies indicated its therapeutic potential in cancer, especially noteworthy, is its co-blockade with other ICRs such as PD-1 during the clinic trials. Yet, additional efforts may be needed to further investigate the molecular mechanism of Tim-3 function in diverse contexts for successfully developing the novel agent targeting Tim-3.
Footnotes
Acknowledgments
This review has been supported by the Natural Science Foundation of Zhejiang Province (LY18C070002); National Natural Science Foundation of China (81803069); Zhejiang Medical Technology Plan Project (2019RC007, 2019KY007, 2021KY047), Public Welfare Technology Project of Zhejiang Province (LGF21H160033) and the 521 Talent Project of Zhejiang Sci-Tech University.
Conflict of interest
The authors declare that they have no conflict of interest.
Authors’ contributions
Wang YG and Huang F contributed to the conception and design of the paper, literature review, critical revision, and approval of the final version; Wang YG, Cao YT, Liu HH, He XL and Li Q searched the literature and wrote the paper.
References
1.
SunF.GuoZ.S.GregoryA.D.ShapiroS.D.XiaoG. and QuZ., Dual but not single PD-1 or TIM-3 blockade enhances oncolytic virotherapy in refractory lung cancer, J Immunother Cancer8 (2020), e000294.
2.
ZhouQ.MungerM.E.VeenstraR.G.WeigelB.J.HirashimaM.MunnD.H.MurphyW.J.AzumaM.AndersonA.C.KuchrooV.K. and BlazarB.R., Coexpression of Tim-3 and PD-1 identifies a CD8+ T-cell exhaustion phenotype in mice with disseminated acute myelogenous leukemia, Blood117 (2011), 4501–4510.
3.
TanS.XuY.WangZ.WangT.DuX.SongX.GuoX.PengJ.ZhangJ.LiangY.LuJ.PengJ.GaoC.WuZ.LiC.LiN.GaoL.LiangX. and MaC., Tim-3 hampers tumor surveillance of liver-resident and conventional NK cells by disrupting pi3k signaling, Cancer Res80 (2020), 1130–1142.
4.
ZhouE.HuangQ.WangJ.FangC.YangL.ZhuM.ChenJ.ChenL. and DongM., Up-regulation of Tim-3 is associated with poor prognosis of patients with colon cancer, Int J Clin Exp Pathol8 (2015), 8018–8027.
5.
McIntireJ.J.UmetsuS.E.AkbariO.PotterM.KuchrooV.K.BarshG.S.FreemanG.J.UmetsuD.T. and DeKruyffR.H., Identification of Tapr (an airway hyperreactivity regulatory locus) and the linked Tim gene family, Nat Immunol2 (2001), 1109–1116.
6.
KuchrooV.K.UmetsuD.T.DeKruyffR.H. and FreemanG.J., The TIM gene family: Emerging roles in immunity and disease, Nat Rev Immunol3 (2003), 454–462.
7.
Ocana-GuzmanR., L. Torre-Bouscoulet and Sada-OvalleI., TIM-3 regulates distinct functions in macrophages, Front Immunol7 (2016), 229.
8.
DasM.ZhuC. and KuchrooV.K., Tim-3 and its role in regulating anti-tumor immunity, Immunol Rev276 (2017), 97–111.
9.
CaoE.ZangX.RamagopalU.A.MukhopadhayaA.FedorovA.FedorovE.ZencheckW.D.LaryJ.W.ColeJ.L.DengH.XiaoH.DilorenzoT.P.AllisonJ.P.NathensonS.G. and AlmoS.C., T cell immunoglobulin mucin-3 crystal structure reveals a galectin-9-independent ligand-binding surface, Immunity26 (2007), 311–321.
10.
SabatosC.A.ChakravartiS.ChaE.SchubartA.Sanchez-FueyoA.ZhengX.X.CoyleA.J.StromT.B.FreemanG.J. and KuchrooV.K., Interaction of Tim-3 and Tim-3 ligand regulates T helper type 1 responses and induction of peripheral tolerance, Nat Immunol4 (2003), 1102–1110.
11.
MonneyL.SabatosC.A.GagliaJ.L.RyuA.WaldnerH.ChernovaT.ManningS.GreenfieldE.A.CoyleA.J.SobelR.A.FreemanG.J. and KuchrooV.K., Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease, Nature415 (2002), 536–541.
12.
HastingsW.D.AndersonD.E.KassamN.KoguchiK.GreenfieldE.A.KentS.C.ZhengX.X.StromT.B.HaflerD.A. and KuchrooV.K., TIM-3 is expressed on activated human CD4(+) T cells and regulates Th1 and Th17 cytokines, Eur J Immunol39 (2009), 2492–2501.
13.
NakaeS.IwakuraY.SutoH. and GalliS.J., Phenotypic differences between Th1 and Th17 cells and negative regulation of Th1 cell differentiation by IL-17, J Leukocyte Biol81 (2007), 1258–1268.
14.
KhademiM.IllesZ.GielenA.W.MartaM.TakazawaN.Baecher-AllanC.BrundinL.HannerzJ.MartinC.HarrisR.A.HaflerD.A.KuchrooV.K.OlssonT.PiehlF. and WallstromE., T Cell Ig- and mucin-domain-containing molecule-3 (TIM-3) and TIM-1 molecules are differentially expressed on human Th1 and Th2 cells and in cerebrospinal fluid-derived mononuclear cells in multiple sclerosis, J Immunol172 (2004), 7169–7176.
15.
WangH.W.ZhuX.L.QinL.M.QianH.J. and WangY., Microglia activity modulated by T cell Ig and mucin domain protein 3 (Tim-3), Cell Immunol293 (2015), 49–58.
16.
NdhlovuL.C.Lopez-VergesS.BarbourJ.D.JonesR.B.JhaA.R.LongB.R.SchoefflerE.C.FujitaT.NixonD.F. and LanierL.L., Tim-3 marks human natural killer cell maturation and suppresses cell-mediated cytotoxicity, Blood119 (2012), 3734–3743.
17.
AndersonA.C.AndersonD.E.BregoliL.HastingsW.D.KassamN.LeiC.ChandwaskarR.KarmanJ.SuE.W.HirashimaM.BruceJ.N.KaneL.P.KuchrooV.K. and HaflerD.A., Promotion of tissue inflammation by the immune receptor Tim-3 expressed on innate immune cells, Science318 (2007), 1141–1143.
18.
OikawaT.KamimuraY.AkibaH.YagitaH.OkumuraK.TakahashiH.ZeniyaM.TajiriH. and AzumaM., Preferential involvement of Tim-3 in the regulation of hepatic CD8+ T cells in murine acute graft-versus-host disease, J Immunol177 (2006), 4281–4287.
19.
NakaeS.IikuraM.SutoH.AkibaH.UmetsuD.T.DekruyffR.H.SaitoH. and GalliS.J., TIM-1 and TIM-3 enhancement of Th2 cytokine production by mast cells, Blood110 (2007), 2565–2568.
20.
Frisancho-KissS.NylandJ.F.DavisS.E.BarrettM.A.GatewoodS.J.NjokuD.B.CihakovaD.SilbergeldE.K.RoseN.R. and FairweatherD., Cutting edge: T cell Ig mucin-3 reduces inflammatory heart disease by increasing CTLA-4 during innate immunity, J Immunol176 (2006), 6411–6415.
21.
YanJ.ZhangY.ZhangJ.P.LiangJ.LiL. and ZhengL., Tim-3 expression defines regulatory T cells in human tumors, PLoS One8 (2013), e58006.
22.
AndersonA.C.LordG.M.DardalhonV.LeeD.H.Sabatos-PeytonC.A.GlimcherL.H. and KuchrooV.K., T-bet, a Th1 transcription factor regulates the expression of Tim-3, Eur J Immunol40 (2010), 859–866.
23.
ChiharaN.MadiA.KondoT.ZhangH.AcharyaN.SingerM.NymanJ.MarjanovicN.D.KowalczykM.S.WangC.KurtulusS.LawT.EtminanY.NevinJ.BuckleyC.D.BurkettP.R.BuenrostroJ.D.Rozenblatt-RosenO.AndersonA.C.RegevA. and KuchrooV.K., Induction and transcriptional regulation of the co-inhibitory gene module in T cells, Nature558 (2018), 454–459.
24.
ZhuC.SakuishiK.XiaoS.SunZ.ZaghouaniS.GuG.WangC.TanD.J.WuC.RangachariM.PertelT.JinH.T.AhmedR.AndersonA.C. and KuchrooV.K., An IL-27/NFIL3 signalling axis drives Tim-3 and IL-10 expression and T-cell dysfunction, Nat Commun6 (2015), 6072.
25.
WadaJ. and KanwarY.S., Identification and characterization of galectin-9, a novel beta-galactoside-binding mammalian lectin, J Biol Chem272 (1997), 6078–6086.
26.
ZhuC.AndersonA.C.SchubartA.XiongH.ImitolaJ.KhouryS.J.ZhengX.X.StromT.B. and KuchrooV.K., The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity, Nat Immunol6 (2005), 1245–1252.
27.
VanceJ.E., Molecular and cell biology of phosphatidylserine and phosphatidylethanolamine metabolism, Prog Nucleic Acid Res Mol Biol75 (2003), 69–111.
28.
HuangY.H.ZhuC.KondoY.AndersonA.C.GandhiA.RussellA.DouganS.K.PetersenB.S.MelumE.PertelT.ClaytonK.L.RaabM.ChenQ.BeaucheminN.YazakiP.J.PyzikM.OstrowskiM.A.GlickmanJ.N.RuddC.E.PloeghH.L.FrankeA.PetskoG.A.KuchrooV.K. and BlumbergR.S., CEACAM1 regulates TIM-3-mediated tolerance and exhaustion, Nature517 (2015), 386–390.
29.
ChibaS.BaghdadiM.AkibaH.YoshiyamaH.KinoshitaI.Dosaka-AkitaH.FujiokaY.OhbaY.GormanJ.V.ColganJ.D.HirashimaM.UedeT.TakaokaA.YagitaH. and JinushiM., Tumor-infiltrating DCs suppress nucleic acid-mediated innate immune responses through interactions between the receptor TIM-3 and the alarmin HMGB1, Nature Immunology13 (2012), 832–842.
30.
YuanJ.JiangB.ZhaoH. and HuangQ., Prognostic implication of TIM-3 in clear cell renal cell carcinoma, Neoplasma61 (2014), 35–40.
31.
CaoY.ZhouX.HuangX.LiQ.GaoL.JiangL.HuangM. and ZhouJ., Tim-3 expression in cervical cancer promotes tumor metastasis, PLoS One8 (2013), e53834.
32.
ZhuangX.ZhangX.XiaX.ZhangC.LiangX.GaoL.ZhangX. and MaC., Ectopic expression of TIM-3 in lung cancers: A potential independent prognostic factor for patients with NSCLC, Am J Clin Pathol137 (2012), 978–985.
33.
PiaoY.R.PiaoL.Z.ZhuL.H.JinZ.H. and DongX.Z., Prognostic value of T cell immunoglobulin mucin-3 in prostate cancer, Asian Pac J Cancer Prev14 (2013), 3897–3901.
34.
YangM.YuQ.LiuJ.FuW.CaoY.YuL.ShaoS.WangX.NiuH. and WangY., T-cell immunoglobulin mucin-3 expression in bladder urothelial carcinoma: Clinicopathologic correlations and association with survival, J Surg Oncol112 (2015), 430–435.
35.
JiangJ.JinM.S.KongF.CaoD.MaH.X.JiaZ.WangY.P.SuoJ. and CaoX., Decreased galectin-9 and increased Tim-3 expression are related to poor prognosis in gastric cancer, PLoS One8 (2013), e81799.
36.
LiH.WuK.TaoK.ChenL.ZhengQ.LuX.LiuJ.ShiL.LiuC.WangG. and ZouW., Tim-3/galectin-9 signaling pathway mediates T-cell dysfunction and predicts poor prognosis in patients with hepatitis B virus-associated hepatocellular carcinoma, Hepatology56 (2012), 1342–1351.
37.
ZhangH.SongY.YangH.LiuZ.GaoL.LiangX. and MaC., Tumor cell-intrinsic Tim-3 promotes liver cancer via NF-kappaB/IL-6/STAT3 axis, Oncogene37 (2018), 2456–2468.
38.
LiuF.LiuY. and ChenZ., Tim-3 expression and its role in hepatocellular carcinoma, J Hematol Oncol11 (2018), 126.
39.
YanW.LiuX.MaH.ZhangH.SongX.GaoL.LiangX. and MaC., Tim-3 fosters HCC development by enhancing TGF-beta-mediated alternative activation of macrophages, Gut64 (2015), 1593–1604.
40.
SolinasG.GermanoG.MantovaniA. and AllavenaP., Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation, J Leukoc Biol86 (2009), 1065–1073.
41.
HastingsW.D.AndersonD.E.KassamN.KoguchiK.GreenfieldE.A.KentS.C.ZhengX.X.StromT.B.HaflerD.A. and KuchrooV.K., TIM-3 is expressed on activated human CD4+ T cells and regulates Th1 and Th17 cytokines, Eur J Immunol39 (2009), 2492–2501.
42.
AsakuraH.KashioY.NakamuraK.SekiM.DaiS.ShiratoY.AbedinM.J.YoshidaN.NishiN.ImaizumiT.SaitaN.ToyamaY.TakashimaH.NakamuraT.OhkawaM. and HirashimaM., Selective eosinophil adhesion to fibroblast via IFN-gamma-induced galectin-9, J Immunol169 (2002), 5912–5918.
43.
P.S. van de WeyerMuehlfeitM.KloseC.BonventreJ.V.WalzG. and KuehnE.W., A highly conserved tyrosine of Tim-3 is phosphorylated upon stimulation by its ligand galectin-9, Biochem Biophys Res Commun351 (2006), 571–576.
44.
RangachariM.ZhuC.SakuishiK.XiaoS.KarmanJ.ChenA.AnginM.WakehamA.GreenfieldE.A.SobelR.A.OkadaH.McKinnonP.J.MakT.W.AddoM.M.AndersonA.C. and KuchrooV.K., Bat3 promotes T cell responses and autoimmunity by repressing Tim-3-mediated cell death and exhaustion, Nat Med18 (2012), 1394–1400.
45.
WolfY.AndersonA.C. and KuchrooV.K., TIM3 comes of age as an inhibitory receptor, Nat Rev Immunol20 (2020), 173–185.
46.
RotteA., Combination of CTLA-4 and PD-1 blockers for treatment of cancer, J Exp Clin Cancer Res38 (2019), 255.
47.
SakuishiK.NgiowS.F.SullivanJ.M.TengM.W.KuchrooV.K.SmythM.J. and AndersonA.C., TIM3(+) FOXP3(+) regulatory T cells are tissue-specific promoters of T-cell dysfunction in cancer, Oncoimmunology2 (2013), e23849.
48.
TangR.RangachariM. and KuchrooV.K., Tim-3: A co-receptor with diverse roles in T cell exhaustion and tolerance, Semin Immunol42 (2019), 101302.
49.
JiangX.ZhouT.XiaoY.YuJ.DouS.ChenG.WangR.XiaoH.HouC.WangW.ShiQ.FengJ.MaY.ShenB.LiY. and HanG., Tim-3 promotes tumor-promoting M2 macrophage polarization by binding to STAT1 and suppressing the STAT1-miR-155 signaling axis, Oncoimmunology5 (2016), e1211219.
50.
LeeJ.SuE.W.ZhuC.HainlineS.PhuahJ.MorocoJ.A.SmithgallT.E.KuchrooV.K. and KaneL.P., Phosphotyrosine-dependent coupling of Tim-3 to T-cell receptor signaling pathways, Mol Cell Biol31 (2011), 3963–3974.
51.
ClaytonK.L.HaalandM.S.Douglas-VailM.B.MujibS.ChewG.M.NdhlovuL.C. and OstrowskiM.A., T cell Ig and mucin domain-containing protein 3 is recruited to the immune synapse, disrupts stable synapse formation, and associates with receptor phosphatases, J Immunol192 (2014), 782–791.
52.
AveryL.FildermanJ., A.L. Szymczak-Workman and KaneL.P., Tim-3 co-stimulation promotes short-lived effector T cells, restricts memory precursors, and is dispensable for T cell exhaustion, Proc Natl Acad Sci U S A115 (2018), 2455–2460.
53.
LiuJ.F.MaS.R.MaoL.BuL.L.YuG.T.LiY.C.HuangC.F.DengW.W.KulkarniA.B.ZhangW.F. and SunZ.J., T-cell immunoglobulin mucin 3 blockade drives an antitumor immune response in head and neck cancer, Mol Oncol11 (2017), 235–247.
54.
LiuJ.F.WuL.YangL.L.DengW.W.MaoL.WuH.ZhangW.F. and SunZ.J., Blockade of TIM3 relieves immunosuppression through reducing regulatory T cells in head and neck cancer, J Exp Clin Cancer Res37 (2018), 44.
55.
JinH.T.AndersonA.C.TanW.G.WestE.E.HaS.J.ArakiK.FreemanG.J.KuchrooV.K. and AhmedR., Cooperation of Tim-3 and PD-1 in CD8 T-cell exhaustion during chronic viral infection, Proc Natl Acad Sci USA107 (2010), 14733–14738.
56.
SakuishiK.ApetohL.SullivanJ.M.BlazarB.R.KuchrooV.K. and AndersonA.C., Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity, J Exp Med207 (2010), 2187–2194.
57.
FourcadeJ.SunZ.PaglianoO.ChauvinJ.M.SanderC.JanjicB.TarhiniA.A.TawbiH.A.KirkwoodJ.M.MoschosS.WangH.GuillaumeP.LuescherI.F.KriegA.AndersonA.C.KuchrooV.K. and ZarourH.M., PD-1 and Tim-3 regulate the expansion of tumor antigen-specific CD8(+) T cells induced by melanoma vaccines, Cancer Res74 (2014), 1045–1055.
58.
FourcadeJ.SunZ.BenallaouaM.GuillaumeP.LuescherI.F.SanderC.KirkwoodJ.M.KuchrooV. and ZarourH.M., Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients, J Exp Med207 (2010), 2175–2186.
59.
NgiowS.F., B. von ScheidtAkibaH.YagitaH.TengM.W. and SmythM.J., Anti-TIM3 antibody promotes T cell IFN-gamma-mediated antitumor immunity and suppresses established tumors, Cancer Res71 (2011), 3540–3551.
60.
LiuJ.ZhangS.HuY.YangZ.LiJ.LiuX.DengL.WangY.ZhangX.JiangT. and LuX., Targeting PD-1 and Tim-3 Pathways to Reverse CD8 T-Cell Exhaustion and Enhance Ex Vivo T-Cell Responses to Autologous Dendritic/Tumor Vaccines, J Immunother39 (2016), 171–180.
61.
BarlowJ.L.WongS.H.BallantyneS.J.JolinH.E. and McKenzieA.N., Tim1 and Tim3 are not essential for experimental allergic asthma, Clin Exp Allergy41 (2011), 1012–1021.
62.
WaterhouseP.PenningerJ.M.TimmsE.WakehamA.ShahinianA.LeeK.P.ThompsonC.B.GriesserH. and MakT.W., Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4, Science270 (1995), 985–988.
63.
NishimuraH.NoseM.HiaiH.MinatoN. and HonjoT., Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor, Immunity11 (1999), 141–151.
64.
Twumasi-BoatengK.PettigrewJ.L.KwokY.Y.E.BellJ.C. and NelsonB.H., Oncolytic viruses as engineering platforms for combination immunotherapy, Nat Rev Cancer18 (2018), 419–432.
65.
QiY.XiaY.LinZ.QuY.QiY.ChenY.ZhouQ.ZengH.WangJ.ChangY.BaiQ.WangY.ZhuY.XuL.ChenL.KongY.ZhangW.DaiB.LiuL.GuoJ. and XuJ., Tumor-infiltrating CD39(+)CD8(+) T cells determine poor prognosis and immune evasion in clear cell renal cell carcinoma patients, Cancer Immunol Immunother69 (2020), 1565–1576.
66.
Sade-FeldmanM.YizhakK.BjorgaardS.L.RayJ.P.de BoerC.G.JenkinsR.W.LiebD.J.ChenJ.H.FrederickD.T.Barzily-RokniM.FreemanS.S.ReubenA.HooverP.J.VillaniA.C.IvanovaE.PortellA.LizotteP.H.ArefA.R.ElianeJ.P.HammondM.R.VitzthumH.BlackmonS.M.LiB.GopalakrishnanV.ReddyS.M.CooperZ.A.PaweletzC.P.BarbieD.A.Stemmer-RachamimovA.FlahertyK.T.WargoJ.A.BolandG.M.SullivanR.J.GetzG. and HacohenN., Defining T cell states associated with response to checkpoint immunotherapy in melanoma, Cell175 (2018), 998–1013, e1020.
67.
YuM.LuB.LiuY.MeY.WangL. and ZhangP., Tim-3 is upregulated in human colorectal carcinoma and associated with tumor progression, Mol Med Rep15 (2017), 689–695.
68.
IelasiL.SansoneV.GranitoA.BeneventoF., S. De Lorenzo and TovoliF., An update of treatments of hepatocellular carcinoma in patients refractory to sorafenib, Drugs Today (Barc)54 (2018), 615–627.
69.
GranitoA.MarinelliS.TerziE.PiscagliaF.RenzulliM.VenerandiL.BeneventoF. and BolondiL., Metronomic capecitabine as second-line treatment in hepatocellular carcinoma after sorafenib failure, Dig Liver Dis47 (2015), 518–522.
70.
MurtazaA.LakenH., J. Da Silva CorreiaMcNeeleyP.AltobellL.ZhangJ.VancutsemP.WilcoxenK. and JenkinsD., Discovery of TSR-022, a novel, potent anti-human TIM-3 therapeutic antibody, Eur J Cancer69 (2016), S102.
71.
WeissG.J.LukeJ.J.FalchookG.ErogluZ.WangJ.HamiltonE.HechtJ.R.LoRussoP.EderJ.P.HughesL.WangJ.RunningK.McEachernK.BobilevD. and RibasA., A phase 1 study of TSR-022, an anti-TIM-3 monoclonal antibody, in patients (pts) with advanced solid tumors, J Immunother Cancer5 (2017), 86.
72.
DavarD.BoasbergP.ErogluZ.FalchookG.GainorJ.HamiltonE.HechtJ.R.LukeJ.PishvaianM.RibasA.WangJ.McEachernK.WaszakA.LuS.LiY.WangY. and LoRussoP., A phase 1 study of TSR-022, an anti-TIM-3 monoclonal antibody, in combination with TSR-042 (anti-PD-1) in patients with colorectal cancer and post-PD-1 NSCLC and melanoma, J Immunother Cancer6 (2018), 115.
73.
HardingJ.PatnaikA.MorenoV.SteinM.JankowskaA.M.MendizabalN.LiuZ.KoneruM. and CalvoE., A phase Ia/Ib study of an anti-TIM-3 antibody (LY3321367) monotherapy or in combination with an anti-PD-L1 antibody (LY3300054): Interim safety, efficacy, and pharmacokinetic findings in advanced cancers, Journal of Clinical Oncology37 (2019), 12.
74.
Giuseppe CuriglianoH.G.MachN.DoiT.TaiW.M.D.FordeP.SarantopoulosJ.BedardP.L.LinC.C.HodiS.WilgenhofS.SantoroA.Sabatos-PeytonC.LongmireT.WanK.NikolopoulosP.ManentiL. and NaingA., Phase (Ph) I/II study of MBG453 spartalizumab (PDR001) in patients (pts) with advanced malignancies [Abstract CT183], Cancer Res79 (2019), CT183.
75.
ZhangD.JiangF.ZaynagetdinovR.HuangH.SoodV.D.WangH.ZhaoX.JenkinsM.H.JiQ.WangY.NannemannD.P.MusilD.WesolowskiJ.PaolettiA.BartholomewT.DernerM.G.AnQ.IfflandC. and HalleJ.P., Identification and characterization of M6903, an antagonistic anti-TIM-3 monoclonal antibody, Oncoimmunology9 (2020), 1744921.
76.
ChenM.WangL.WangY.ZhouX.LiuX.ChenH.HuangB. and HuZ., Soluble Tim3 detection by time-resolved fluorescence immunoassay and its application in membranous nephropathy, J Clin Lab Anal34 (2020), e23248.