The N-myc downstream-regulated gene 1 (NDRG1) has been discovered as a significant gene in the progression of cancers. However, the regulatory mechanism of NDRG1 remained obscure in prostate cancer (PCa).
METHODS:
The miR-96-5p and NDRG1 expression levels were evaluated in PCa cell lines, prostate tissues, and validated public databases by real-time PCR, western blot analysis, and immunohistochemistry. The function of miR-96-5p and NDRG1 were investigated by wound healing and transwell assays in vitro, and mouse xenograft assay in vivo. The candidate pathway regulated by NDRG1 was conducted by the next-generation gene sequencing technique. Immunofluorescence and luciferase assay was used to detect the relation between miR-96-5p, NDRG1, and NF-B pathway.
RESULTS:
Overexpressing NDRG1 suppresses the migration, invasion, and epithelial-mesenchymal transition (EMT) in vitro, and inhibits metastasis in vivo. Moreover, miR-96-5p contributes to NDRG1 deficiency and promotes PCa cell migration and invasion. Furthermore, NDRG1 loss activates the NF-B pathway, which stimulates p65 and IKBa phosphorylation and induces EMT in PCa.
CONCLUSIONS:
MiR-96-5p promotes the migration and invasion of PCa by targeting NDRG1 and regulating the NF-B pathway.
Prostate cancer (PCa) was the second common malignant tumor in males. Based on the GLOBOCAN 2020 estimates of cancer, there were 1,414,259 new cases of PCa worldwide. Moreover, there were also 375,304 patients who died of PCa [1]. Most of the patients were in an advanced stage at the time of first diagnosis with lymph nodes or distance metastasis especially in eastern Asian countries [2]. Although several treatments have been improved recently, the prognosis of PCa remains not well. Therefore, it is necessary to look for reliable biomarkers that play a crucial role in tumor growth and metastasis to distinguish early stage tumors.
N-myc downstream-regulated gene-1 (NDRG1) is located on chromosome 8q24.3. containing 16 exons and 15 introns. It is an important member of the NDRG family [3, 4]. Epithelial-mesenchymal transition (EMT) is known as a crucial phenotypic conversion. In the process of EMT, cells can change from epithelial state to mesenchymal state in a highly plastic and dynamic way, modify the adhesion molecules expressed by cells, and make them have migration and invasive behavior [5, 6]. Some previous studies have demonstrated that NDRG1 potentially inhibited cancer cell migration, invasion, and EMT by affecting the indispensable molecules involved in metastasis [7, 8]. Similar studies by LC Tu and RS Jin et al. suggested that NDRG1 could bind to -catenin and E-cadherin directly, indicating that these proteins formed a complex. Under the stimulation of WNT3a, ectopic overexpression of NDRG1 maintained -catenin on the plasma membrane of colon and PCa cells, which blocked the activation of the wnt pathway [9, 10]. In addition, RX Xi et al. have recently found an effective way to inhibit TNF- and LYRIC induced EMT through NDRG1 [11]. NDRG1 also modulated EMT by regulating the level of related protein expression. Elevating E-cadherin and inhibiting vimentin, N-cadherin, slug, and snail-1 have been established by JC Lee et al. in oral squamous cancer cells [12]. It has also been suggested thatNDRG1 potentially attenuates EMT by inhibiting the expression of Smad2 and suppressing the latter related protein in nasopharyngeal cancer [13]. Though several molecular pathways illustrated the function of NDRG1 have been partially elucidated, the underlying mechanism responsible for NDRG1 and EMT in PCa is still needed further exploration.
MicroRNA (miRNA) is a small endogenous non-coding RNA that binds to a specific sequence of target gene 3 ’untranslated region (3’UTR) and affects the post-transcriptional regulation of the target gene, which might result in mRNA degradation or translation inhibition [14]. The dysregulation of miRNA in cancer is widely confirmed, and several types of research have shown that the level of miRNA expression wasrelated to tumor metastasis [15, 16]. MiR-96 is a member of the miR-183 cluster and has been demonstrated to act as an oncogene or tumor suppressor in tumors [17, 18, 19, 20, 21]. Furthermore, recent literature has identified that miR-96 also modulated EMT by regulating the level of related protein expression. Notably, CF HE et al. showed that miR-96 regulated the protein of EMT-related gene FOXQ1, which could be modulated by TGF-1 in bladder cancer [22]. A study by HY Wang et al. implicated that miR-96-5p promoted proliferation and EMT by directly regulating FOXF2 expression in oral squamous cell carcinoma [23]. Moreover, the circPTPRA and RASSF8 suppressed EMT and metastasis of non-small cell lung carcinomas also by competitive sponging miR-96-5p [24]. MiR-96-5p wasmarkedly upregulated in PCa compared to normal tissue and it acted as an oncogene in some previous studies [25, 26, 27, 28]. However, the mechanisms of miR-96-5p as a tumor promoter in PCa have not been fully elucidated.
The NF-B pathway has been studied for many years, and its significant role in immunity inflammation, and other physiological processes has been fully confirmed. Previous studies showed that EMT was potently administrated by several signaling pathways, such as TGF-, Wnt, and ErbB [29, 30]. While accumulating studies have indicated that the NF-B signaling pathway also played a crucial role in inducing and maintaining EMT in a variety of tumors [31, 32, 33, 34]. Furthermore, some evidence has illustrated that activation of the NF-B pathway was also related to the metastasis of PCa [16, 35].
Materials and methods
Clinical samples and cell lines
Twenty paired PCa and adjacent normal prostate tissue were collected during surgery at the second hospital of Tianjin Medical University in 2019. RWPE-1, LNCaP, C4-2, PC3, and DU145 cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). RWPE-1 cells were grown in keratinocyte-SFM (Invitrogen). LNCaP, C4-2, PC3, and DU145 were cultured in RPMI1640 medium containing 10% fetal bovine serum (FBS) and penicillinstreptomycin (100 U/mL) (Solarbio, Beijing, China). All cell lines were incubated under a humidified atmosphere at 37C with 5% CO2. This study was approved by the Human Ethics Committee.
Western blot
After cleaning with cold PBS twice, the tissues or cells were collected and then extracted the whole cell lysate with RIPA buffer (Solarbio, Beijing, China concentration was measured by a detection kit (Solarbio, Beijing, China) according to the manufacturer’s guidance. The same amount of protein in each sample was placed in 10% gel by SDS-PAGE and then transferred to the nitrocellulose membrane. 5% non-fat milk or 5% BSA (Solarbio, Beijing, China)was utilized to block the membrane, then incubated with primary antibodies anti- (NDRG1, diluted 1:10000, Ki-67, diluted 1:1000, Abcam; p65, p-p65, IkBa, p-IkBa, diluted 1:1000, Cell Signaling Technology; E-cadherin, N-cadherin diluted 1:1000, GAPDH, diluted 1:2000, vimentin, diluted 1:10000 Proteintech). After incubation with the corresponding secondary peroxidase-conjugated antibody, the protein bands were displayed with chemiluminescence dissolvent (Solarbio, Beijing, China).
qRT-PCR analysis
Total RNA was extracted by utilizing TRIzol reagent (Invitrogen) following the manufacturer’s guidance. Then equal amounts of 1-5g total RNA was transcribed to cDNA according to the protocol supplied with HiFiScript cDNA Synthesis Kit (Cwbio, Taizhou, China). The mRNA expression was measured by SYBR Green qPCR master mix. The expression levels of all mRNA were normalized to GAPDH or U6. All assays were conducted three times independently. The primers for qRT-PCR were listed as followed: NDRG1 Forward: 5’-GTCCTTATCAACGTGAACCCTT-3’, Reverse: 5’-GCATTGGTCGCTCAATCTCCA-3’; GAPDH Forward: 5’-GCTCTCTGCTCCTCCTGTTC-3’, Reverse: 5’-ACGACCAAATCCGTTGACTC-3’; miR-96-5p Forward: 5’-CCTCGATTTGGCACTAGCAC-3’, Reverse: 5’-TATGGTTGTTCTGCTCTCTGTCTC-3’; U6 Forward: 5’-CGCTTCGGCAGCATATAC-3’, Reverse: 5’-TTCACGAATTTGCGTGTCATC-3’.
Immunofluorescence
Each group of cells was seeded on 24-well plates using glass coverslips (Beijing Solarbio), fixed with 4% paraformaldehyde, infiltrated with 0.5% Triton X-100. Then 5% BSA was used to block cells for 1 hour and incubated with the primary antibody. NDRG1, (diluted 1:200, Abcam) p-P65 (diluted 1:1600, CST) at 4C overnight, followed by incubation with FITC-labeled or TRITC-labeled secondary antibody (diluted 1:200, Proteintech, China) for 2 h. Finally, the cells were stained with DAPI, and images were detected under a confocal microscope (FV500; Olympus).
Immunohistochemical analysis
Tissue microarray was purchased from Alenabio, Xi’an (PR803d). The six tissue with lymph node metastasis were also collected from the second hospital of Tianjin Medical University. Standard immunohistochemistry (IHC) protocols were applied with specific antibodies following previously reported [36].
Luciferase reporter assay
Luciferase reporter assay was conducted with the Dual-Luciferase Reporter Assay kit (Promega, USA) pmirGLO; pmirGLO-WT and pmirGLO-MUT for NDRG1-3’UTR were cotransfected into 293T cell lines, together with miR-96-5p or miR-183-5p mimics, by using Lipoferctamine 2000 (Invitrogen, USA). After transfection in 48 h, Renilla luciferase activity acts as a standard of the relative luciferase activity. Three independent assays were performed.
Cell transfection
Cells were transfected with the control, siRNA, miRNA mimics/inhibitor were purchased from GenePharma (Shanghai, China) using Lipofectamine 2000 (Thermo Fisher) following the manufacturer’s guidance. siRNAs for NDRG1 were defined as siNDRG1#1 and siNDRG1#2, siNDRG1#3 respectively. All siRNA are listed as followed: si1: NDRG1-homo-688; si2: NDRG1-homo-835; si3: NDRG1-homo-956. To overexpress NDRG1, full-length NDRG1 was cloned into a modified LV18 lentiviral vector (GenePharma, Shanghai, China), while an empty vector served as a control was screened with puromycin. Luciferase express LV11 lentiviral vector (GenePharma, Shanghai, China) were transfected and screened with neomycin.
Wound healing
LNCaP, C4-2, PC3, and DU145 cells were added into 6-well plates with a density of 6 10 cells per well. When the cells are fully attached, the 2 mm scratch will divide the well into three equal parts. Next, the cells were cultured in a serum-free medium for 48 hours. The cells were detected and photographed at 0 h, 24 h, and 48 h, respectively. The scratch healing rate was calculated by ImageJ software.
Migration and invasion assays
Cell migration and invasion abilities were identified by transwell chambers which consist of an 8-mm membrane filter covered with or without Matrigel (BD Biosciences). Cells were cultured with serum-free medium for 24 h before use. Cells (1–2 10) in serum-free medium were seeded to the upper chambers. Lower chambers were supplied with medium with 10% FBS. After incubation for 24–48 h, The cells at the bottom of the chamber were fixed with 4% paraformaldehyde, infiltrated with 0.5% Triton X-100, stained with DAPI, and counted by a fluorescence microscope.
Animal experiment
Animal experiments have been approved by the Ethics Committee of Tianjin Medical University. BALB/c nude male mice (5–6 weeks old) were raised in a room under sterile conditions. The DU145 cells with high expression of NDRG1 were directly subcutaneously injected to establish the model of subcutaneous xenograft. ( 5 mice/group, 3.0 10 cells/100 l). The cells were added to Matrigel (BD Biosciences) with a culture medium (1:1 diluted). D-luciferin/D-PBS at a concentration of 15 mg/ml were given through intraperitoneal injections with 10 l/g for imaging after six weeks of feeding and then all primary tumors were carried out. The two mice with suspected metastatic foci were kept feeding for eight weeks. Primary tumors and suspected metastatic foci were excised, embedded, and fixed. H&E staining and immunohistochemistry analyses were conducted.
Statistical analysis
SPSS v.21.0 software (USA) and GraphPad Prism Software 7.0 (USA) were used for data assessment. Mean SD was used to show experimental outcomes. Student’s -test or one-way ANOVA was used to reveal the differences between groups. values 0.05 were considered statistically significant.
Results
NDRG1 expression is decreased in PCa and affected the prognosis of patients
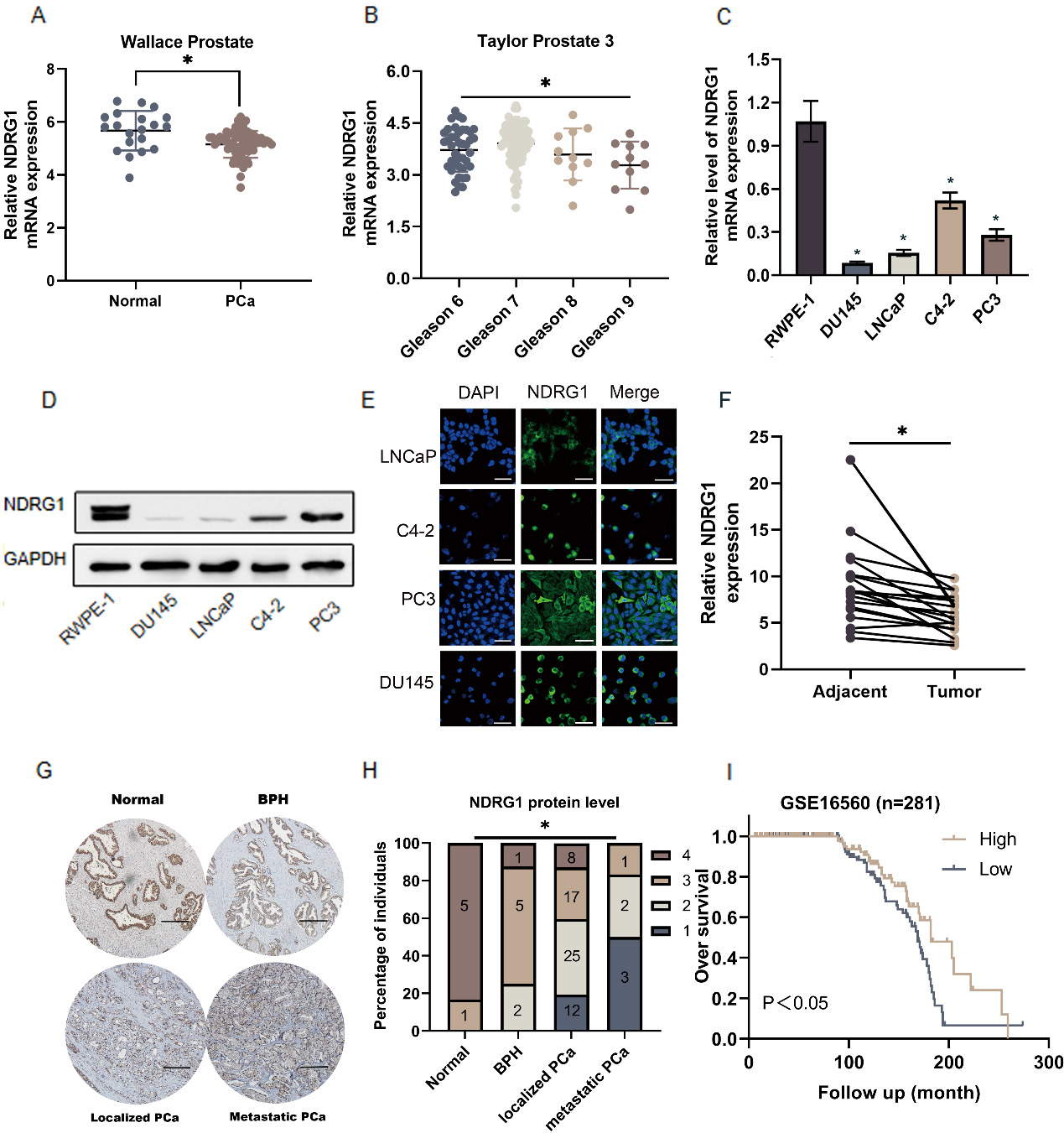
To investigate the clinical significance of NDRG1 in PCa, the mRNA sequencing dataset of PCa from the Oncomine database (https://www.oncomine.org/resource/login.html) was evaluated. The dataset showed that NDRG1 expression was decreased in PCa compared with the normal tissues and showed a declined trend with Gleason score increased (Fig. 1A and B). The expression of NDRG1 was also down-regulated in mRNA and protein levels in PCa cells (DU145 LNCaP C4-2 PC3) compared with normal prostate epithelial cells (RWPE-1) (Fig. 1C and D). Most of the proteins are located in the cytoplasm (Fig. 1E). Our study further measured the expression levels of NDRG1 in 20 paired PCa patients and an 80-point tissue chip combined with six metastatic specimens. The NDRG1 expression level in PCa was also decreased. There was a tendency of gradual decrease of NDRG1 level in normal tissue, benign prostatic hyperplasia (BPH) tissue, localized PCa and metastatic PCa tissue judged by immunohistochemistry (Fig. 1F–H, Fig. S1). Furthermore, the patients with a high level of NDRG1 had higher overall survival than the low according to the dates from GSE16560 ( 281) (https://www.ncbi.nlm.nih.gov/).
NDRG1 expression is decreased in prostate cancer (PCa) and affected the prognosis of patients. a–b NDRG1 expression was decreased in PCa tissues compared with the normal tissues and showed a declined trend with Gleason score increased by analyzing the Oncomine PCa mRNA sequencing dataset, 0.05. c Real-time PCR analysis of NDRG1 expression levels in the normal prostate epithelial cell (RWPE-1), PCa cell lines (LNCaP, C4-2, PC3, DU145), 0.05. d Western blot analysis of each cell line. e NDRG1 location and relative level in each PCa cell lines by immunofluorescence, scale bar 25 m. f 20 paired PCa tissues compared with the matched adjacent normal tissues as assessed by real-time PCR, 0.05. g–h Immunohistochemistry analysis of NDRG1 in 80-point tissue chip combined with six metastatic specimens including normal prostate tissue, benign prostatic hypertrophy (BPH), localized PCa and metastatic PCa. The NDRG1 staining intensity was categorized into four groups: 1 (no or poor staining), 2 (weak staining), 3 (intermediate staining), and 4 (strong staining), scale bar 250 m. i Kaplan-Meier survival analyses of PCa patients with high or low NDRG1 expression based on GSE16560 dataset ( 281).
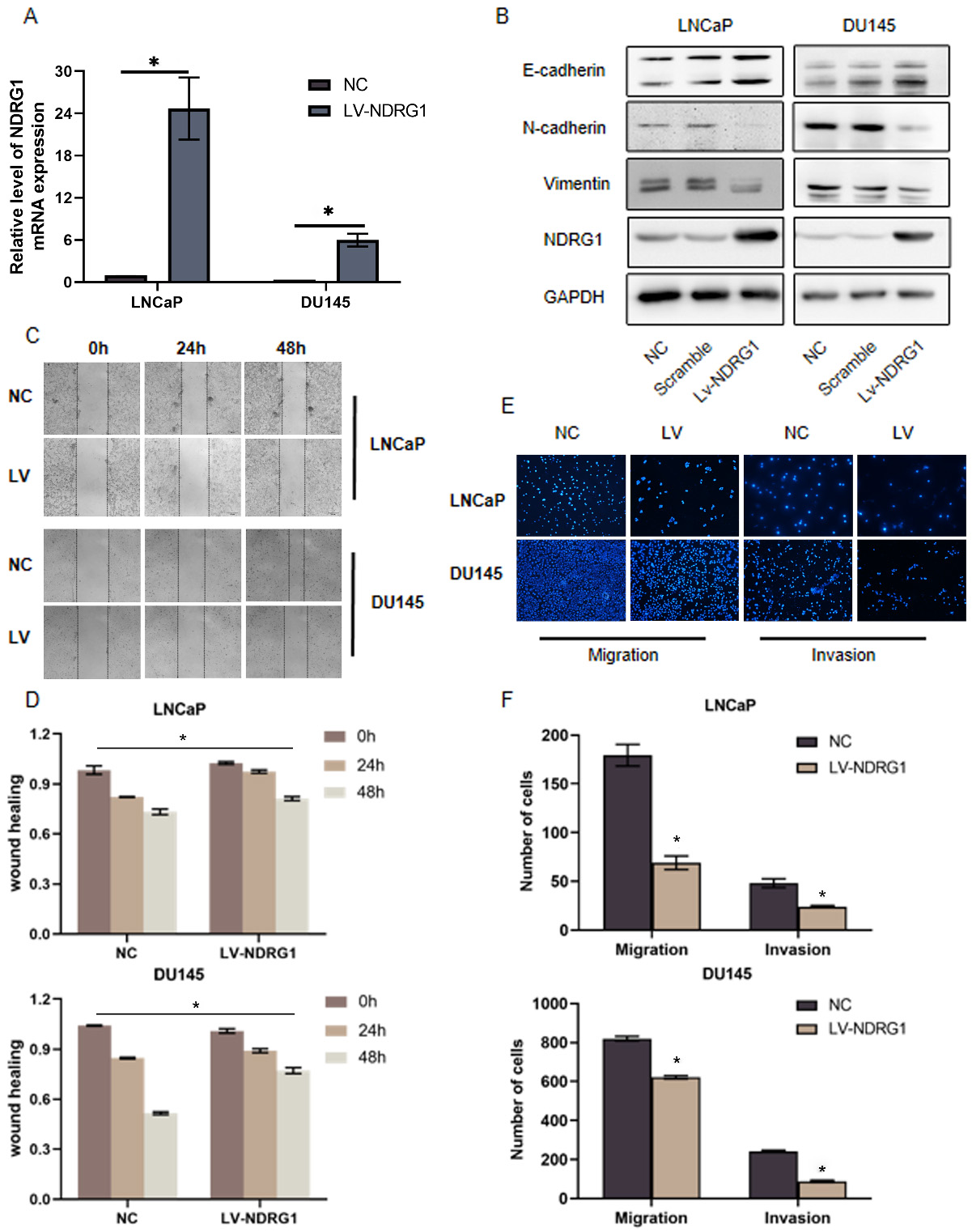
Overexpression NDRG1 suppresses EMT, migration, and invasion in PCa cells. a Real-time PCR validate the effect of overexpression NDRG1 in two cell lines (LNCaP, DU145), 0.05. b Overexpression of NDRG1 increased E-cadherin expression and decreased vimentin and N-cadherin expression in LNCaP and DU145 cells. c–d The migration of prostate cells were detected by wound healing assays after overexpressing NDRG1, 0.05. e–f The migration and invasion of prostate cells were assessed by transwell assays after overexpressing NDRG1, 0.05.
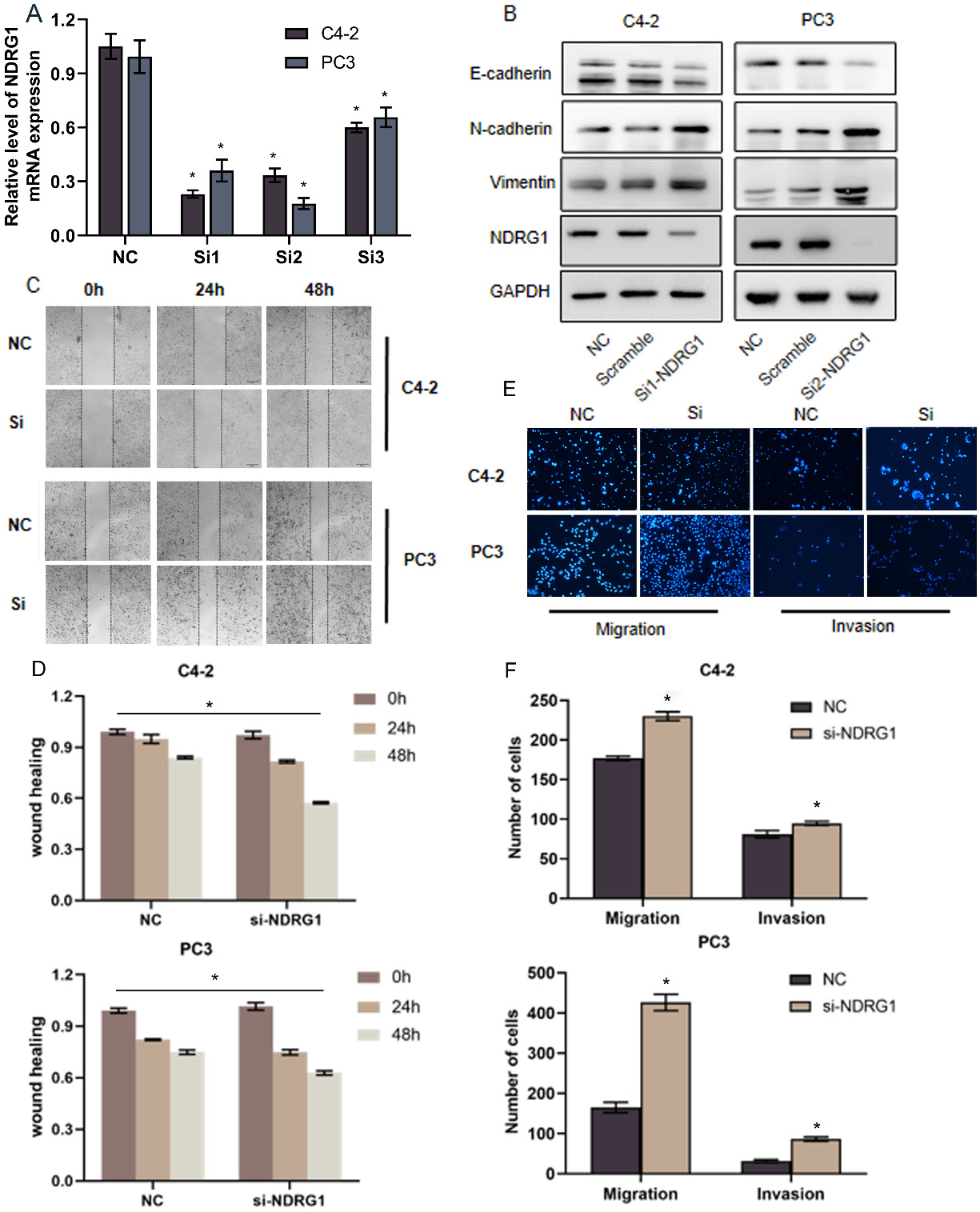
Silencing NDRG1 promotes EMT, migration, and invasion in PCa cells. a Real-time PCR validate the effect of downregulating NDRG1 in two cell lines (C4-2, PC3) by transfect with three siRNA, 0.05. b Silencing NDRG1 decreased E-cadherin expression and increased Vimentin and N-cadherin expression in C4-2 and PC3 cells. c–f c–d The migration of prostate cells were detected by wound healing assays after down-regulating NDRG1, 0.05. e–f The migration and invasion of prostate cells were assessed by transwell assays after down-regulating NDRG1, 0.05.
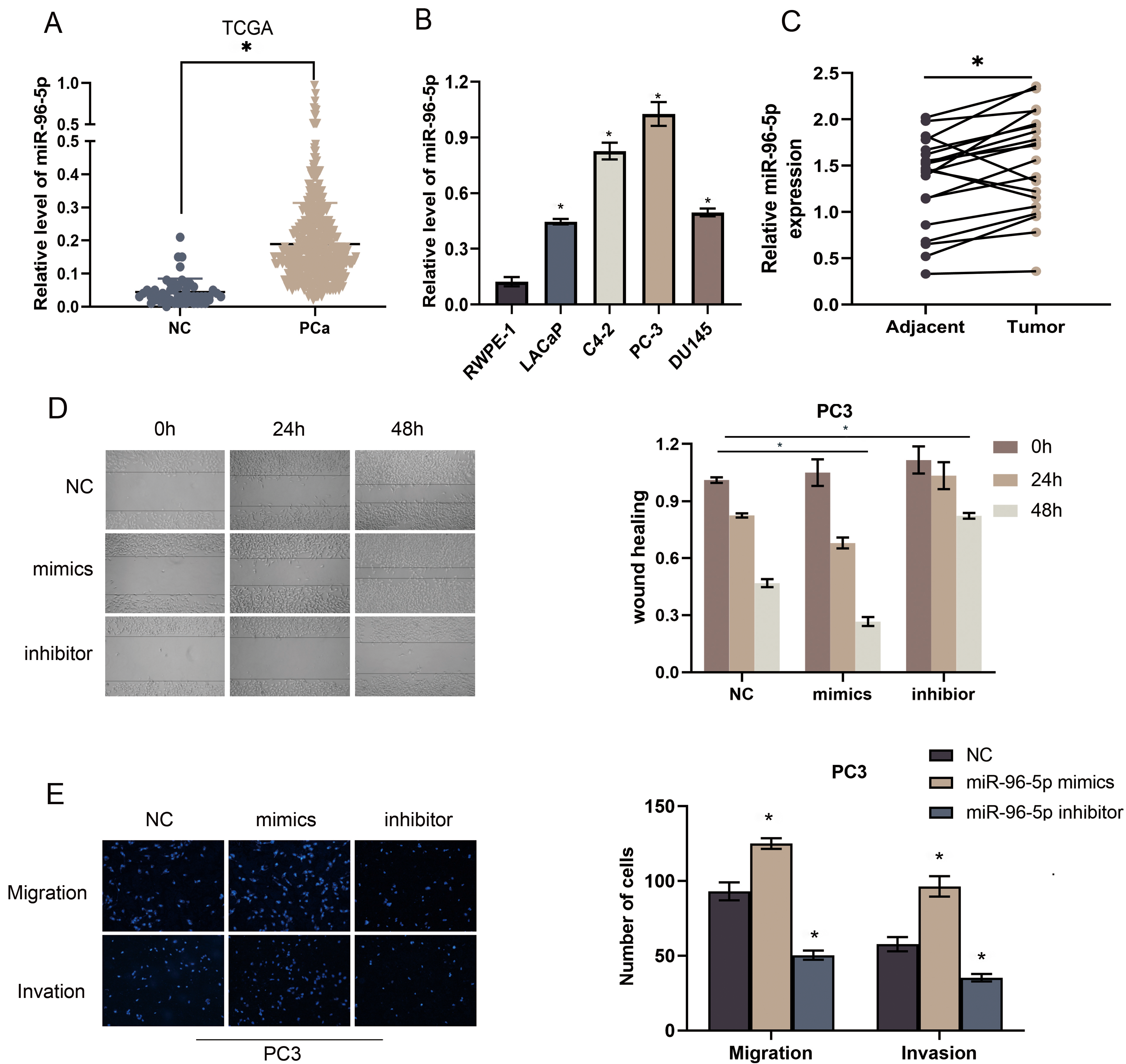
MiR-96-5p expression is increased in PCa and promotes EMT in PCa cells. a MiR-96-5p expression in PCa based on TCGA, 0.05. b Real-time PCR analysis of miR-96-5p expression levels in the normal prostate epithelial cell (RWPE-1), PCa cell lines (LNCaP, C4-2, PC3, DU145), 0.05. c Real-time PCR analysis of miR-96-5p expression levels in adjacent tumor tissues and tumor tissues, 0.05. d–e Overexpression of miR-96-5p enhanced while silencing miR-96-5p suppressed invasion and migration abilities in the PC3 cell line, 0.05.
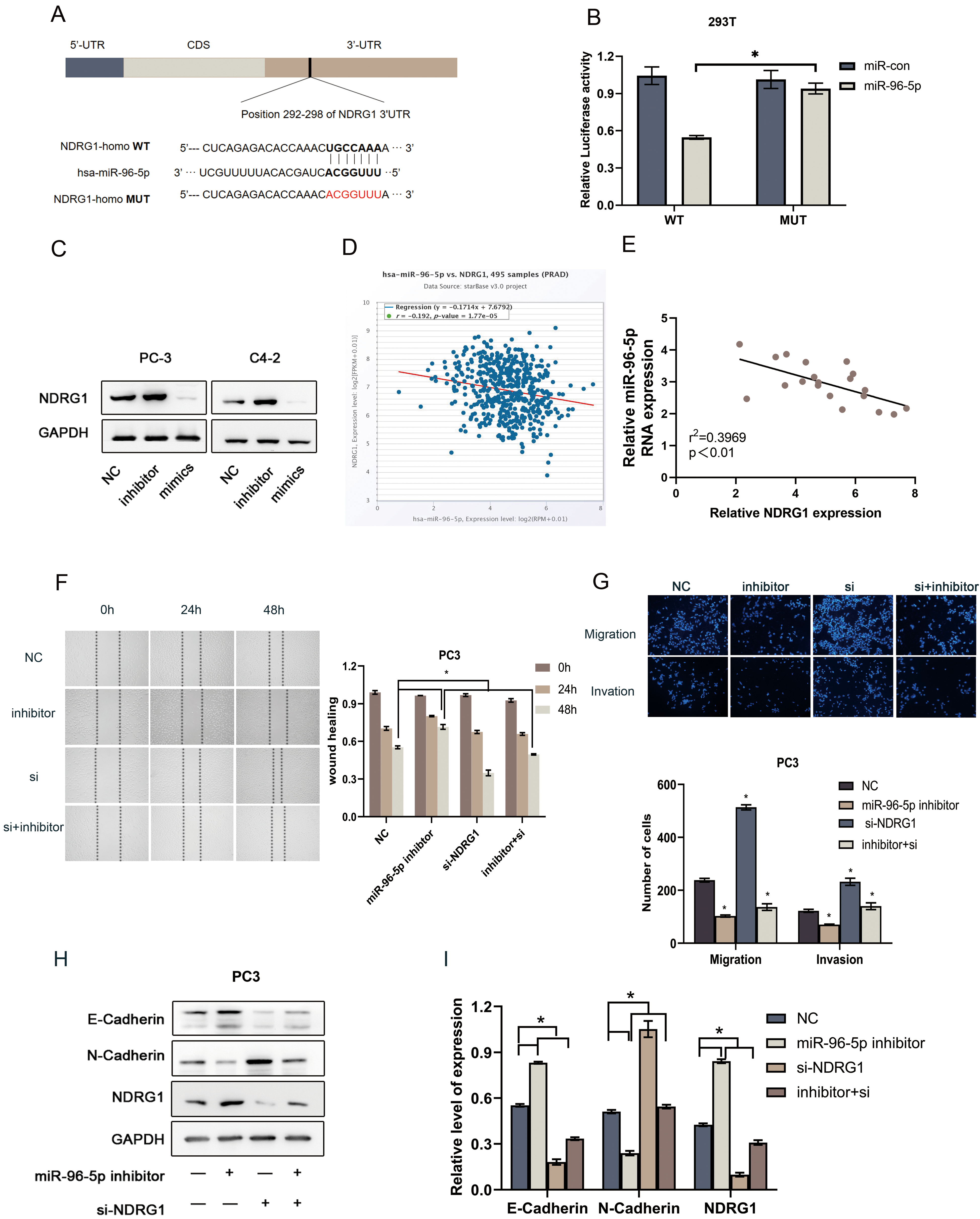
NDRG1 is directly targeted by miR-96-5p and important for miR-96-5p mediated EMT. a MiR-96-5p-binding sequence in NDRG1 3’UTR. A mutation was generated in NDRG1 3’UTR in the complementary site for miR-96-5p binding. b The luciferase reporter assay was used to validate the relationship between miR-96-5p and NDRG1 in 293T cells, 0.05. c Downregulation of miR-96-5p increased NDRG1 expression, while overexpression of miR-96-5p decreased NDRG1 expression. d–e Spearman’s correlation analysis of the TCGA database and clinical samples showed that miR-96-5p expression was negatively correlated with NDRG1 mRNA level in PCa tissues. f–g The ability of migration and invasion in PC3 cells transfected with miR-96-5p inhibitor, siNDRG1 or co-transfection compared with control group. hI Western blot of indicating proteins changed in PC3 cells, 0.05.
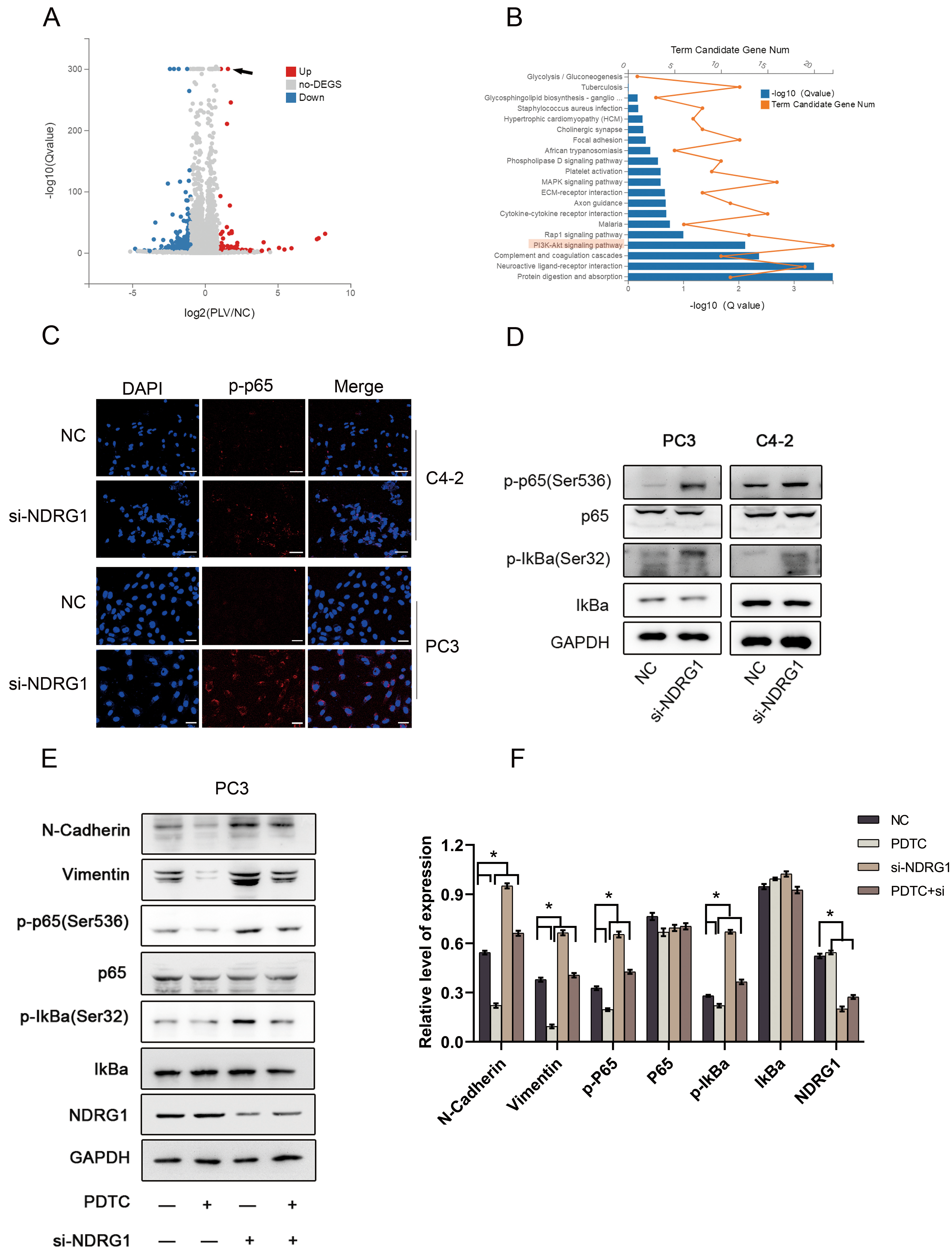
Down-regulating NDRG1 activates NF-B signaling pathway promoting EMT in PCa cells. a Volcano plot of genes changed between treatment group and control group in DU145 cells (the red arrow refers to overexpressed NDRG1). b Go enrichment analysis of downstream pathway changes gene number c. Immunofluorescence show that knocking down the expression of NDRG1 could increase the number of phosphorylated p65 (p-p65) and increase its nuclear translocation in C4-2 and PC3 cell, scale bar 25 m. d Western blot analysis of NF-B pathway proteins in C42 and PC3 cells when suppressed NDRG1 level. e–f Western blot use to assess the effects of PDTC or NDRG1 on the protein levels of NF-B pathway proteins in PC3 cells, 0.05.
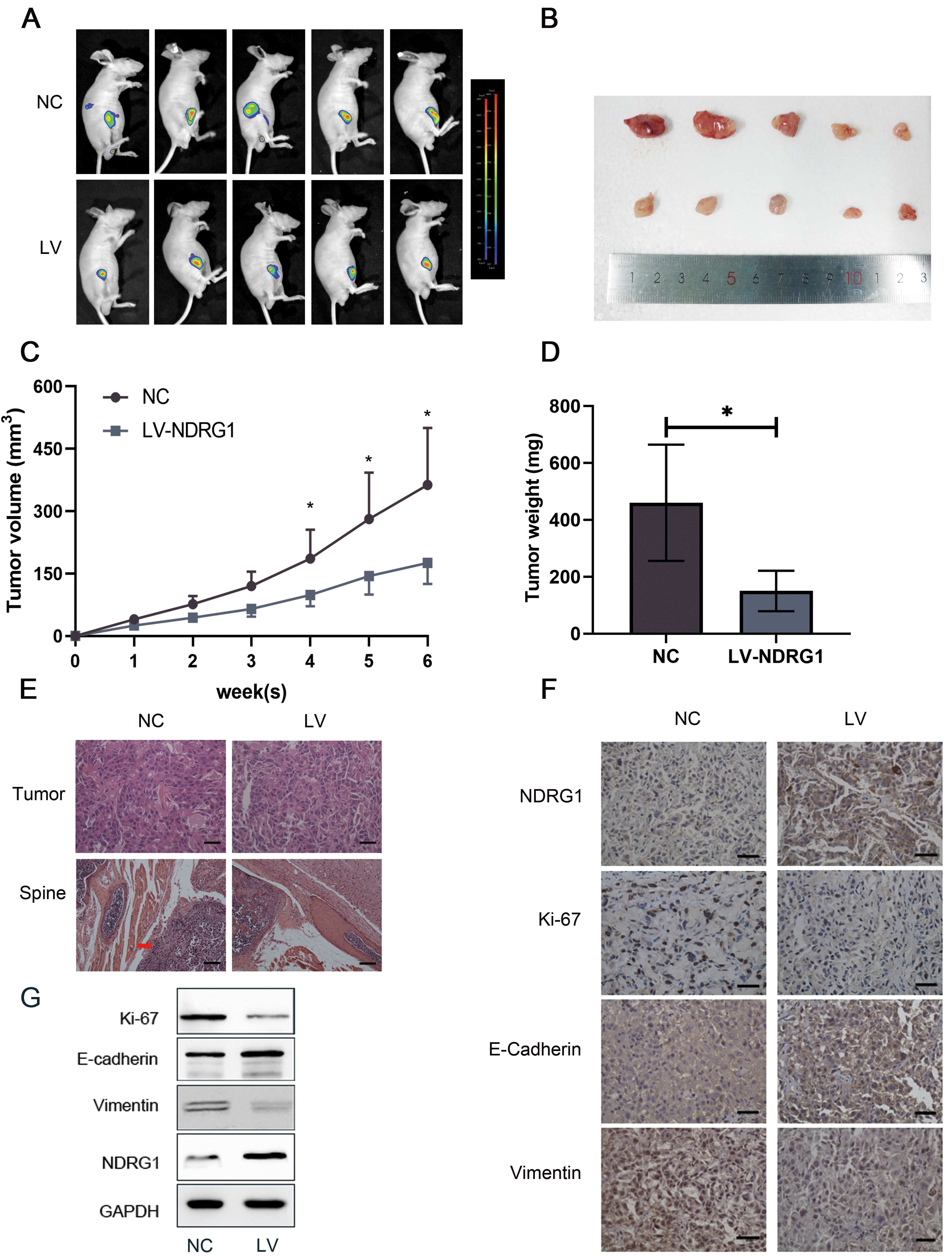
Elevating NDRG1 inhibits PCa metastasis in vivo. a Images of the tumors in the subcutaneous mouse model. b Images of subcutaneous tumors formed by the DU145 cells. c–d Growth curves and weight analyses of subcutaneous tumors formed by DU145 cells, 0.05. e H&E staining show tumor in situ and one distant metastasis lesion near the spine, red arrow indicated the metastatic tumor area, scale bar 25 m (tumor), 100 m (spine). f IHC staining of tumor in situ, scale bar 25 m g Western blot of proteins changed in tumor tissues.
NDRG1 suppresses EMT, migration, and invasion in PCa cells
To determine the function of NDRG1 in human PCa cells, we transfected lentivirus to overexpress NDRG1 in two cell lines (LNCaP DU145) with relatively low expression of NDRG1 (Fig. 2A) and siRNA to silenced NDRG1 in relatively high expression of NDRG1 cell lines (C4-2 PC3). To achieve a better-silenced effect, we selected three siRNA to identified in two cell lines (Fig. 3A). The C42 and PC3 were knocked out with siRNA1 and siRNA2, respectively. Relative to control cells, NDRG1-overexpressing cells was significantly inhibited migration and NDRG1 knockdown cells significantly promoted migration in the wound healing assay (Figs 2C and D, 3C and D). Transwell assays were also performed and the result indicated a consistent trend in migration. The overexpression group showed a strong ability to inhibit invasion and the knocking down group showed a high ability to promote invasion especially in the PC3 cell line (Figs 2E and F, 3E and F). Furthermore, western blot analysis implicated that upregulating NDRG1 increased the expression of E-cadherin and decreased vimentin and N-cadherin in LNCaP and DU145 cells (Fig. 2B); Conversely, silencing NDRG1 had an opposite effect on these EMT markers (Fig. 3B). These results demonstrate that NDRG1 suppresses migration, invasion, and EMT in PCa cells in vitro.
miR-96-5p expression is increased in PCa and promote EMT in PCa cells
The miR-96-5p expression was analyzed by miRNA sequencing dataset of PCa from The Cancer Genome Atlas (TCGA) (https://www.cancer.gov/tcga). The data suggested that miR-96-5p was significantly up-regulated in PCa compared with adjacent normal tissues (Fig. 4A). We further investigated the expression levels of miR-96-5p in different cell lines. Compared with the RWPE1 cell line, the expression of miR-96-5p was significantly increased (Fig. 4B) In the tumor tissues we found the same trend (Fig. 4C). Then, transfection of PC3 cells was performed with mimic and inhibitor of miR-96-5p. The wound-healing assays revealed that Overexpression or knocking down of miR-96-5p can promote or inhibit migration, respectively (Fig. 4D). Transwell assays further confirmed that miR-96-5p could promote the migration and invasion of PCa (Fig. 4E).
NDRG1 is directly targeted by miR-96-5p and important for miR-96-5p mediated EMT
Our previous study found that miR-182-5p couldtarget the expression of NDRG1 [37]. To further explored the possible mechanism that contributed to NDRG1 downregulation in PCa, we used bioinformatics databases TargetScanHuman 7.2 (http://www.targetscan.org) and Starbase 2.0 (https://starbase.sysu.edu.cn/starbase2/) to predict potential miRNA targets in NDRG1 mRNA. Interestingly, we found miR-96-5p and miR-183-5p may both have the opportunity to combine with NDRG1 mRNA 3’-UTR binding site. Moreover, miR-183 96 182 as a cluster had been considered to play a significant role in tumor cell survival, proliferation, and migration. Further sequence analysis indicated the presence of only one putative binding site for miR-96-5p and miR-183-5p in the NDRG1 3’-UTR. Luciferase assay was carried out detecting the changes of miR-96-5p (Fig. 5B) and miR-183-5p (Fig. S2) transiently transfected with WT NDRG1 3’-UTR and mutated NDRG1 3’-UTR in 293T cells. As shown in Fig. 2, luciferase activity was remarkably repressed only in WT NDRG1 3’-UTR. While there was no significant change in fluorescence value when mutated NDRG1 3’-UTR co-transfected with miR-96-5p in 293T cells. To further verified the regulatory relationship, western blot was conducted in different treatment groups in PC3 cells. Figure 5C showed that miR-96-5p inhibitor could up-regulate NDRG1 expression while miR-96-5p mimics down-regulate NDRG1 expression. Moreover, spearman’s correlation analysis of the TCGA database demonstrated that miR-96-5p mRNA level was negatively correlated with NDRG1 mRNA level in PCa tissues (0.192, 0.001; Fig. 5D) by using the starbase database. Clinical samples were also demonstrated that the expression of mRNA level was negatively correlated with each other( 0.397, 0.01; Fig. 5E). There were significant differences in the ability of migration and invasion between the control group and the group transfected with miR-96-5p inhibitoror siNDRG1 in PC3 cells(Fig. 5F and G). Western blot of indicating proteins changed also illustrated that NDRG1 was crucial to miR-96-5p mediated EMT(Fig. 5F and G).
To further assessed the mechanism of NDRG1 promoting the metastasis of PCa, the next generation sequence was used to compared overexpressed NDRG1 cells in DU145 with normal cells (Fig. 6A). By comparing the go enrichment analysis of downstream pathway changes, we found that the PI3K-AKT pathway has significantly modified (Fig. 6B). It has been confirmed that NF-B could be activated as a downstream pathway of pAKT in many researches So we further explored the relationship between NDRG1 and NF-B pathway. We analyzed the subcellular localization of phosphorylated p65 (p-p65) in PC3 and C4-2 cells using immunofluorescence. Knocking down the expression of NDRG1 could increase the number of p-p65 and promoteits nuclear translocation (Fig. 6C). Western blot also demonstrated that downregulating NDRG1 increased the expression of p-p65 and phosphorylated IkBa (p-IkBa) in C4-2 and PC3 cells (Fig. 6D). To further verify whether NDRG1 can mediate EMT through the NF-B pathway, we added Pyrrolidinedithiocarbamate ammonium (PDTC) with 1.0 ng/ml, an inhibitor of NF-B pathway. When PDTC was added in PC3 cells, the content of p-p65 and p-IkBa was induced as well as the expression of vimentin and N-cadherin. However, there was an increased level ofp-p65 and p-IkBa when down-regulating NDRG1 Moreover, inhibition of NF-B signaling by PTDC impaired the stimulatory effect of NDRG1 down-expression on EMT in PCa cells (Fig. 6E and F). The above data indicate that down-regulating NDRG1 activates NF-B signaling pathway promoting EMT in PCa cells.
Elevating NDRG1 inhibits PCa metastasis in vivo
To determine the function of NDRG1 on the metastasis of PCa in vivo, we used lentivirus to overexpress NDRG1 in luciferase-labeled vector DU145 cells and established the xenograft model by directly subcutaneous injection of tumor cells into nude mice. As shown in Fig. 7A, compared with the control group, the metastatic ability of the overexpression group was lower. After feeding for 6 weeks, there were two mice in the control group had distant metastases, while no suspicious lesions were found in the treatment group under the imaging system. We also discovered that the tumor volume and weight were significant differences between the two groups. Upregulating NDRG1 inhibited tumor proliferation at the same time (Fig. 7B–D). H&E staining showed the tumor in situ and one typical distant metastasis lesion near the spine. The red arrow indicated the metastatic tumor area (Fig. 7E). Furthermore, the immunohistochemistry of tumor tissue and western blot demonstrated that up-regulating NDRG1 dramatically reduced the level of ki-67 and vimentin, while increased E-cadherin (Fig. 7F and G). Consequently, these findings demonstrate that elevating NDRG1 inhibits PCa proliferation and metastasis in vivo.
Discussion
The main findings of our study display novel insights that miR-96-5p induced NDRG1 deficiency and activated NF-B signal pathway, which further promoted EMT of PCa. Here, we observed that NDRG1 expression was decreased in PCa tissues, and high expression of miR-96-5p correlated with PCa cell migration and invasion. Our outcomes further indicated that miR-96-5p via directly targeting NDRG1 promoted EMT in PCa cells, leading to the development of PCa metastasis. Therefore, our study uncovered novel insights that miR-96-5p promotes EMT through NDRG1 and NF-B signal pathway, elucidating the tumor suppressor gene of NDRG1 and oncogenic function of miR-96-5p in PCa.
The role of NDRG1 in inhibiting the progression of metastasis has been described in different cancer researches [38, 39, 40]. Our previous study also partly elucidated the mechanism of decreased level of NDRG1 in PCa and its possible ways of regulating expression [36, 37]. In fact, available evidence has indicated that NDRG1 might act as a tumor suppressor in several PCa cell lines through different molecular mechanisms [9, 11, 41]. However, the mechanism of NDRG1 regulating migration and invasion is still partly unknown. In our study, we illustrated that inhibiting the activation of the NF-B pathway could change the EMT markers induced by NDRG1 depletion, andmiR-96-5p inhibitors overexpression also reversed the effect of NDRG1 down-regulation to the EMT markers. We also demonstrated that overexpression of NDRG1 inhibited the proliferation and metastasis of PCa in vivo. Higher miR-96-5p expression was also accompanied with lower NDRG1 expression in PCa tissues, which further illustrates the negative relation between the two molecules in PCa progression. More importantly, these demonstrate that miR-96-5p regulates NDRG1 expression and NDRG1 regulates EMT by modulating NF-B activation, in part, state that NDRG1 suppressive function in cell migration, invasion, and EMT mediate by miR-96-5p and NF-B.
MiR-183 cluster is a significant gene located on the short arm of chromosome 7 (7q32.2). Three mature miRNAs (miR-96, -182, and -183) were generated from a single polycistronic transcript. The expression of the miR-183 cluster has been documented in several cancersMost studies assumed that the miR-183 cluster has shownan oncogenic function in cancers [42, 43, 44], while some have suggested inhibition of these effects [45, 46]. In our previous study, we illustrated that miR-182-5p induced NDRG1 deficiency and promoted proliferation and metastasis in PCa cells [37]. We further explored the effect of miR-96-5p and miR-183-5p on NDRG1 in this study. Luciferase activity was remarkably repressed only in miR-96-5p. Several previous studies have suggested that miR-96-5p was up-regulated in different kinds of tumors and a high level of miR-96-5p promoted cancer cell migration and invasion via different mechanisms and also predicted poor survival. MK Siu et al. illustrated that miR-96-5p could target the tumor suppressor ETV6, which downregulated the expression of EMT markers in PCa [27]. Moreover, TGF- could regulate the expression of miR-96 through Smad-dependent transcription. MiR-96 also promoted bone metastasis in PCa [26]. Notably, a study by MD Long et al. showed that miR-96 controlled the AR signaling pathway and promoted PCa progression by adjusting the RAR network [47]. In our study, these findings reveal that miR-96-5p is elevated in PCa and promoted EMT by regulating NDRG1 expression to control NF-B pathway activity.
A large number of studies have illustrated that the NF-B signal was significantly activated in several kinds of human tumors, which is closely associated with tumor progression and metastasis [31, 48]. In glioma cancer, the activation of the NF-B signal played a significant role in promoting cell migration and invasion [49, 50]. In colorectal cancer, DCLK1 promoted EMT via the PI3K/Akt/NF-B pathway [32]. In addition, a study by LL Mei et al. has demonstrated that the level of miR-145-5p was induced by inhibiting the Sp1/NF-B pathway, which limited the migration and invasion of esophageal squamous cancer cells [33]. Accumulating evidence has illustrated that NF-B signaling played a crucial role in promoting invasion and metastasis of PCa [16, 51]. ZQ Shang et al. showed that lncRNA-PCAT1 induced CRPC progression by positively regulating AKT/NF-B signaling [52]. Notably, JW Zhang et al. discovered that Migration and invasion enhancer 1 was an NF-B induced gene, which enhanced the proliferation and invasion of human PCacells [53]. In the present study, we reveal that miR-96-5p activates the NF-B pathway by directly targeting NDRG1, which promotes EMT in PCa cells. Moreover, we also demonstrate that NF-B signaling activity did play a significant role in the invasion and migration of PCa cells.
Conclusions
In summary we demonstrate that NDRG1 loss is correlated with cell metastasis and poor prognosis of PCa patients. During the development of PCa, the abnormally high expression of miR-96-5p can promote this biological process, inhibit the expression of NDRG1, activate the NF-B pathway, and promote cell EMT. Based on our findings, we propose a new molecular mechanism in which miR-96-5p promotes EMT by regulating NDRG1 expression to control NF-B pathway activation in PCa. These findings provide a new understanding of the progression of PCa and further guide the clinical practices.
Author contributions
Conception: Ranlu Liu, Jing Li.
Interpretation or analysis of data: Zhenpeng Lian, Taihao Chang, Shenfei Ma, Jing Li.
Preparation of the manuscript: Zhenpeng Lian, Hongtuan Zhang, Xiaoming Wang.
Revision for important intellectual content: Zhenpeng Lian, Ranlu Liu, Taihao Chang.
Supervision: Ranlu Liu.
Supplementary data
The supplementary files are available to download from http://dx.doi.org/10.3233/CBM-210072.
sj-docx-1-cbm-10.3233_CBM-210072.docx - Supplemental material
The authors thank professor E Du and Yan Liu for guidance on the experiment. We also thank the whole project team who worked on this study.
This work was supported by the Fund program: the Key Project of Tianjin Municipal Science and Technology Commission (NO.19ZXDBSY00050), Tianjin Municipal Health Bureau (NO.ZC20116), the Project of Henan Science and Technology Department (NO.182102310344) and Henan Health Commission (NO.2018020487).
Conflict of interest
The authors report no conflicts of interest in this work.
References
1.
SungH.FerlayJ.SiegelR.L.LaversanneM.SoerjomataramI.JemalA. and BrayF., Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries, CA Cancer J Clin71 (2021), 209–249.
2.
ZhangK.BangmaC.H. and RoobolM.J., Prostate cancer screening in Europe and Asia, Asian J Urol4 (2017), 86–95.
3.
FangB.A.KovacevicZ.ParkK.C.KalinowskiD.S.JanssonP.J.LaneD.J.SahniS. and RichardsonD.R., Molecular functions of the iron-regulated metastasis suppressor, NDRG1, and its potential as a molecular target for cancer therapy, Biochim Biophys Acta1845 (2014), 1–19.
4.
QuX.ZhaiY.WeiH.ZhangC.XingG.YuY. and HeF., Characterization and expression of three novel differentiation-related genes belong to the human NDRG gene family, Mol Cell Biochem229 (2002), 35–44.
5.
NietoM.A.HuangR.Y.JacksonR.A. and ThieryJ.P., Emt: 2016, Cell166 (2016), 21–45.
6.
ThieryJ.P.AcloqueH.HuangR.Y. and NietoM.A., Epithelial-mesenchymal transitions in development and disease, Cell139 (2009), 871–890.
7.
LiuW.YueF.ZhengM.MerlotA.BaeD.H.HuangM.LaneD.JanssonP.LuiG.Y.RichardsonV.SahniS.KalinowskiD.KovacevicZ. and RichardsonD.R., The proto-oncogene c-Src and its downstream signaling pathways are inhibited by the metastasis suppressor, NDRG1, Oncotarget6 (2015), 8851–8874.
8.
WangpuX.LuJ.XiR.YueF.SahniS.ParkK.C.MenezesS.HuangM.L.ZhengM.KovacevicZ. and RichardsonD.R., Targeting the Metastasis Suppressor, N-Myc Downstream Regulated Gene-1, with Novel Di-2-Pyridylketone Thiosemicarbazones: Suppression of Tumor Cell Migration and Cell-Collagen Adhesion by Inhibiting Focal Adhesion Kinase/Paxillin Signaling, Mol Pharmacol89 (2016), 521–540.
9.
JinR.LiuW.MenezesS.YueF.ZhengM.KovacevicZ. and RichardsonD.R., The metastasis suppressor NDRG1 modulates the phosphorylation and nuclear translocation of beta-catenin through mechanisms involving FRAT1 and PAK4, J Cell Sci127 (2014), 3116–3130.
10.
TuL.C.YanX.HoodL. and LinB., Proteomics analysis of the interactome of N-myc downstream regulated gene 1 and its interactions with the androgen response program in prostate cancer cells, Mol Cell Proteomics6 (2007), 575–588.
11.
XiR.PunI.H.MenezesS.V.FouaniL.KalinowskiD.S.HuangM.L.ZhangX.RichardsonD.R. and KovacevicZ., Novel Thiosemicarbazones Inhibit Lysine-Rich Carcinoembryonic Antigen-Related Cell Adhesion Molecule 1 (CEACAM1) Coisolated (LYRIC) and the LYRIC-Induced Epithelial-Mesenchymal Transition via Upregulation of N-Myc Downstream-Regulated Gene 1 (NDRG1), Mol Pharmacol91 (2017), 499–517.
12.
LeeJ.C.ChungL.C.ChenY.J.FengT.H. and JuangH.H., N-myc downstream-regulated gene 1 downregulates cell proliferation, invasiveness, and tumorigenesis in human oral squamous cell carcinoma, Cancer Lett355 (2014), 242–252.
13.
HuZ.Y.XieW.B.YangF.XiaoL.W.WangX.Y.ChenS.Y. and LiZ.G., NDRG1 attenuates epithelial-mesenchymal transition of nasopharyngeal cancer cells via blocking Smad2 signaling, Biochim Biophys Acta1852 (2015), 1876–1886.
14.
BartelD.P., MicroRNAs: Target recognition and regulatory functions, Cell136 (2009), 215–233.
15.
LiJ.YeD.ShenP.LiuX.ZhouP.ZhuG.XuY.FuY.LiX.SunJ.XuJ. and ZhangQ., Mir-20a-5p induced WTX deficiency promotes gastric cancer progressions through regulating PI3K/AKT signaling pathway, J Exp Clin Cancer Res39 (2020), 212.
16.
RenD.YangQ.DaiY.GuoW.DuH.SongL. and PengX., Oncogenic miR-210-3p promotes prostate cancer cell EMT and bone metastasis via NF-kappaB signaling pathway, Mol Cancer16 (2017), 117.
17.
AndersonO. and Guttilla ReedI.K., Regulation of cell growth and migration by miR-96 and miR-183 in a breast cancer model of epithelial-mesenchymal transition, PLoS One15 (2020), e0233187.
18.
HuangX.LvW.ZhangJ.H. and LuD.L., miR96 functions as a tumor suppressor gene by targeting NUAK1 in pancreatic cancer, Int J Mol Med34 (2014), 1599–1605.
19.
LiuZ.M.WuZ.Y.LiW.H.WangL.Q.WanJ.N. and ZhongY., MiR-96-5p promotes the proliferation, invasion and metastasis of papillary thyroid carcinoma through down-regulating CCDC67, Eur Rev Med Pharmacol Sci23 (2019), 3421–3430.
20.
RessA.L.StiegelbauerV.WinterE.SchwarzenbacherD.KiesslichT.LaxS.JahnS.DeutschA.BauernhoferT.LingH.SamoniggH.GergerA.HoeflerG. and PichlerM., MiR-96-5p influences cellular growth and is associated with poor survival in colorectal cancer patients, Mol Carcinog54 (2015), 1442–1450.
21.
YaoQ.PeiY.ZhangX. and XieB., microRNA-96 acts as a tumor suppressor gene in human osteosarcoma via target regulation of EZRIN, Life Sci203 (2018), 1–11.
22.
HeC.ZhangQ.GuR.LouY. and LiuW., miR-96 regulates migration and invasion of bladder cancer through epithelial-mesenchymal transition in response to transforming growth factor-beta1, J Cell Biochem119 (2018), 7807–7817.
23.
WangH.MaN.LiW. and WangZ., MicroRNA-96-5p promotes proliferation, invasion and EMT of oral carcinoma cells by directly targeting FOXF2, Biol Open9 (2020).
24.
WeiS.ZhengY.JiangY.LiX.GengJ.ShenY.LiQ.WangX.ZhaoC.ChenY.QianZ.ZhouJ. and LiW., The circRNA circPTPRA suppresses epithelial-mesenchymal transitioning and metastasis of NSCLC cells by sponging miR-96-5p, EBioMedicine44 (2019), 182–193.
25.
PudovaE.A.KrasnovG.S.NyushkoK.M.KobelyatskayaA.A.SavvateevaM.V.PoloznikovA.A.DolotkazinD.R.KliminaK.M.GuvatovaZ.G.SimanovskyS.A.GladyshN.S.TokarevA.T.MelnikovaN.V.DmitrievA.A.AlekseevB.Y.KaprinA.D.KiselevaM.V.SnezhkinaA.V. and KudryavtsevaA.V., miRNAs expression signature potentially associated with lymphatic dissemination in locally advanced prostate cancer, BMC Med Genomics13 (2020), 129.
26.
SiuM.K.TsaiY.C.ChangY.S.YinJ.J.SuauF.ChenW.Y. and LiuY.N., Transforming growth factor-beta promotes prostate bone metastasis through induction of microRNA-96 and activation of the mTOR pathway, Oncogene34 (2015), 4767–4776.
27.
TsaiY.C.ChenW.Y.SiuM.K.TsaiH.Y.YinJ.J.HuangJ. and LiuY.N., Epidermal growth factor receptor signaling promotes metastatic prostate cancer through microRNA-96-mediated downregulation of the tumor suppressor ETV6, Cancer Lett384 (2017), 1–8.
28.
VossG.HaflidadottirB.S.JaremoH.PerssonM.Catela IvkovicT.WikstromP. and CederY., Regulation of cell-cell adhesion in prostate cancer cells by microRNA-96 through upregulation of E-Cadherin and EpCAM, Carcinogenesis41 (2020), 865–874.
29.
MiettinenP.J.EbnerR.LopezA.R. and DerynckR., TGF-beta induced transdifferentiation of mammary epithelial cells to mesenchymal cells: Involvement of type I receptors, J Cell Biol127 (1994), 2021–2036.
30.
WangM.RenD.GuoW.HuangS.WangZ.LiQ.DuH.SongL. and PengX., N-cadherin promotes epithelial-mesenchymal transition and cancer stem cell-like traits via ErbB signaling in prostate cancer cells, Int J Oncol48 (2016), 595–606.
31.
HoeselB. and SchmidJ.A., The complexity of NF-kappaB signaling in inflammation and cancer, Mol Cancer12 (2013), 86.
32.
LiuW.WangS.SunQ.YangZ.LiuM. and TangH., DCLK1 promotes epithelial-mesenchymal transition via the PI3K/Akt/NF-kappaB pathway in colorectal cancer, Int J Cancer142 (2018), 2068–2079.
33.
MeiL.L.WangW.J.QiuY.T.XieX.F.BaiJ. and ShiZ.Z., miR-145-5p Suppresses Tumor Cell Migration, Invasion and Epithelial to Mesenchymal Transition by Regulating the Sp1/NF-kappaB Signaling Pathway in Esophageal Squamous Cell Carcinoma, Int J Mol Sci18 (2017).
34.
WangJ.TianL.KhanM.N.ZhangL.ChenQ.ZhaoY.YanQ.FuL. and LiuJ., Ginsenoside Rg3 sensitizes hypoxic lung cancer cells to cisplatin via blocking of NF-kappaB mediated epithelial-mesenchymal transition and stemness, Cancer Lett415 (2018), 73–85.
35.
ZhangJ.KuangY.WangY.XuQ. and RenQ., Notch-4 silencing inhibits prostate cancer growth and EMT via the NF-kappaB pathway, Apoptosis22 (2017), 877–884.
36.
LiY.PanP.QiaoP. and LiuR., Downregulation of N-myc downstream regulated gene 1 caused by the methylation of CpG islands of NDRG1 promoter promotes proliferation and invasion of prostate cancer cells, Int J Oncol47 (2015), 1001–1008.
37.
LiuR.LiJ.TengZ.ZhangZ. and XuY., Overexpressed microRNA-182 promotes proliferation and invasion in prostate cancer PC-3 cells by down-regulating N-myc downstream regulated gene 1 (NDRG1), PLoS One8 (2013), e68982.
38.
BandyopadhyayS.PaiS.K.HirotaS.HosobeS.TakanoY.SaitoK.PiquemalD.CommesT.WatabeM.GrossS.C.WangY.RanS. and WatabeK., Role of the putative tumor metastasis suppressor gene Drg-1 in breast cancer progression, Oncogene23 (2004), 5675–5681.
39.
MatsugakiT.ZenmyoM.HiraokaK.FukushimaN.ShodaT.KomiyaS.OnoM.KuwanoM. and NagataK., N-myc downstream-regulated gene 1/Cap43 expression promotes cell differentiation of human osteosarcoma cells, Oncol Rep24 (2010), 721–725.
40.
MiL.ZhuF.YangX.LuJ.ZhengY.ZhaoQ.WenX.LuA.WangM.ZhengM.JiJ. and SunJ., The metastatic suppressor NDRG1 inhibits EMT, migration and invasion through interaction and promotion of caveolin-1 ubiquitylation in human colorectal cancer cells, Oncogene36 (2017), 4323–4335.
41.
BandyopadhyayS.WangY.ZhanR.PaiS.K.WatabeM.IiizumiM.FurutaE.MohintaS.LiuW.HirotaS.HosobeS.TsukadaT.MiuraK.TakanoY.SaitoK.CommesT.PiquemalD.HaiT. and WatabeK., The tumor metastasis suppressor gene Drg-1 down-regulates the expression of activating transcription factor 3 in prostate cancer, Cancer Res66 (2006), 11983–11990.
42.
YuanJ.DongR.LiuF.ZhanL.LiuY.WeiJ. and WangN., The miR-183/182/96 cluster functions as a potential carcinogenic factor and prognostic factor in kidney renal clear cell carcinoma, Exp Ther Med17 (2019), 2457–2464.
43.
ZhangQ.RenW.HuangB.YiL. and ZhuH., MicroRNA-183/182/96 cooperatively regulates the proliferation of colon cancer cells, Mol Med Rep12 (2015), 668–674.
44.
ZhangW.QianP.ZhangX.ZhangM.WangH.WuM.KongX.TanS.DingK.PerryJ.K.WuZ.CaoY.LobieP.E. and ZhuT., Autocrine/Paracrine Human Growth Hormone-stimulated MicroRNA 96-182-183 Cluster Promotes Epithelial-Mesenchymal Transition and Invasion in Breast Cancer, J Biol Chem290 (2015), 13812–13829.
45.
GilamA.CondeJ.Weissglas-VolkovD.OlivaN.FriedmanE.ArtziN. and ShomronN., Local microRNA delivery targets Palladin and prevents metastatic breast cancer, Nat Commun7 (2016), 12868.
46.
KongW.Q.BaiR.LiuT.CaiC.L.LiuM.LiX. and TangH., MicroRNA-182 targets cAMP-responsive element-binding protein 1 and suppresses cell growth in human gastric adenocarcinoma, FEBS J279 (2012), 1252–1260.
47.
LongM.D.SinghP.K.RussellJ.R.LlimosG.RosarioS.RizviA.van den BergP.R.KirkJ.Sucheston-CampbellL.E.SmiragliaD.J. and CampbellM.J., The miR-96 and RARgamma signaling axis governs androgen signaling and prostate cancer progression, Oncogene38 (2019), 421–444.
48.
DiDonatoJ.A.MercurioF. and KarinM., NF-kappaB and the link between inflammation and cancer, Immunol Rev246 (2012), 379–400.
49.
JiangL.LinC.SongL.WuJ.ChenB.YingZ.FangL.YanX.HeM.LiJ. and LiM., MicroRNA-30e* promotes human glioma cell invasiveness in an orthotopic xenotransplantation model by disrupting the NF-kappaB/IkappaBalpha negative feedback loop, J Clin Invest122 (2012), 33–47.
50.
JiangL.WuJ.YangY.LiuL.SongL.LiJ. and LiM., Bmi-1 promotes the aggressiveness of glioma via activating the NF-kappaB/MMP-9 signaling pathway, BMC Cancer12 (2012), 406.
51.
ChenP.C.ChengH.C. and TangC.H., CCN3 promotes prostate cancer bone metastasis by modulating the tumor-bone microenvironment through RANKL-dependent pathway, Carcinogenesis34 (2013), 1669–1679.
52.
ShangZ.YuJ.SunL.TianJ.ZhuS.ZhangB.DongQ.JiangN.Flores-MoralesA.ChangC. and NiuY., LncRNA PCAT1 activates AKT and NF-kappaB signaling in castration-resistant prostate cancer by regulating the PHLPP/FKBP51/IKKalpha complex, Nucleic Acids Res47 (2019), 4211–4225.
53.
ChangK.S.TsuiK.H.LinY.H.HouC.P.FengT.H. and JuangH.H., Migration and Invasion Enhancer 1 Is an NF-kB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo, Cancers (Basel)11 (2019).