
Abstract
BACKGROUNDS:
Anaplastic thyroid cancer/ATC is highly lethal malignancy without reliable chemotherapeutic drug. Resveratrol possesses anti-ATC activities but encounters resistance in some cases due to certain unknown reason(s).
OBJECTIVE:
Because signal transducer and activator of transcription/STAT3 signaling is critical for ATC cell survival and the main molecular target of resveratrol, its roles in determining the fates of resveratrol-treated ATC cells were investigated here.
METHODS:
Human THJ-11T, THJ-16 and THJ-21T ATC cell lines were treated by 100
RESULTS:
Resveratrol inhibited proliferation, p-STAT3 nuclear translocation as well as LIF and STAT3 expression of THJ-16T and THJ-21T but not THJ-21T cells which showed LIF upregulation and more frequent p-STAT3 nuclear translocation. AG490 significantly prevent p-STAT3 nuclear translocation, and reversed the resveratrol tolerance of THJ-11T cells. Immonohistochemical staining revealed 14.3% (4/28) of LIF and 3.6% (1/28) of p-STAT3 detection in noncancerous ATC-surrounding tissues, which increased to 89.5% (17/19) and 52.6% (10/19) respectively among ATC specimens. The correlative analysis indicated the relevance of LIF expression and STAT3 activation (
CONCLUSIONS:
The status of STAT3 activation and LIF expression are closely correlated with the therapeutic effect of resveratrol on ATCs. Frequent LIF upregulation and STAT3 activation are the unfavorable factors of ATCs and the potential targets of anti-ATC therapy.
Introduction
Thyroid cancer is the most common endocrine malignancy and its incidence keeps increasing [1]. According to the standards of World Health Organization, thyroid cancer can be divided into differentiated and undifferentiated thyroid cancer [2]. Differentiated thyroid cancer has a favorable prognosis and long survival period, while ATC, though accounts for less than 2% of the total thyroid cancer cases, has the highest mortality rate and the patients usually die within one year of diagnosis [3]. Surgery and adjuvant chemotherapy are the standard management of ATCs, but the prognosis is very poor because of the highly aggressive growth and metastasis tendency of the tumors [4, 5, 6]. All-trans retinoic acid has been used to promote ATC cell differentiation for improving iodine uptake and therefore radiosensitivity, while the outcomes are not promising and even show the opposite effects as we have reported recently [7]. Therefore, it would be of clinical values to explore safer and more reliable agent(s) against ATCs.
Resveratrol, a polyphenolic compound commonly found in natural plants, has multiple biological activities, including anti-inflammatory and anti-cancer effects [8]. More importantly, resveratrol in the anticancer dose has little toxic effects on normal cells [9]. Our previous results show that all of three ATC cell lines (THJ-11T, THJ-16T and THJ-21T) so far checked are resistant to RA treatment [10], but two of them (THJ-16T and THJ-21T) are sensitive to resveratrol, suggesting the better therapeutic effects of resveratrol on ATCs. However, THJ-11T cells are not sensitive to resveratrol. This suggests the necessity to investigate the underlying reason(s) leading to differential response of ATC cells to resveratrol for the personalized anti-ATC therapy.
STAT3 pathway is known to play an important role in cancer formation and progression [11, 12]. It regulates cancer stem-cell properties in anaplastic thyroid cancer cells and confers on drug resistance of anaplastic thyroid cancer [13]. STAT3 pathway can be activated by certain cytokines including LIF that has been known to play promoting roles in thyroid carcinogenesis [14, 15]. Because STAT3 pathway is the main molecular target of resveratrol [16, 17], we speculate that the statuses of STAT3 signaling and its upstream stimulators such as LIF may be related to the resveratrol sensitivity of ATC cells. Our study thus aims to address this issue using the ATC cell lines with different resveratrol sensitivity.
Materials and methods
Thyroid cancer cell lines and treatment
Human anaplastic thyroid cancer THJ-11T, THJ-16T and THJ-21T cell lines [18] were kindly provided by Dr. Liu, Institute of Cancer Stem Cell, Dalian Medical University. They were cultured in 1640 medium with L-glutamine (Hyclone, Logan, UT, USA), supplemented with 10% fetal bovine serum (Gibco, Grand island, NY, USA) for THJ-11T and THJ-21T, and with 5% fetal bovine serum for THJ-16T. An amount of 5
Evaluation of cell proliferation and death
To elucidate the response of ATC cells to resveratrol treatment, HE staining, viable/nonviable cell counting (Automated Cell Counter, Bio-Rad, Singapore), 3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT) cell proliferation assay and a modification of the terminal deoxynucleotide transferase (TdT)-mediated dUTP-biotin nick end-labeling method (TUNEL; Roche Inc., Basel, Switzerland) were performed on coverslips bearing THJ-11T, THJ-16T and THJ-21T cells without and with resveratrol treatment by the methods described elsewhere [10].
Western blot analyses
Western blotting was performed using antibodies against
Immunocytochemical staining
Immunocytochemical staining was performed on the coverslips obtained from each of the experimental groups. The antibodies against p-STAT3 (Santa Cruz. Inc., USA; 1:80), LIF (Santa Cruz. Inc., USA): 1:80, NF-
Inhibition of STAT3 activation with AG490
AG490 (Sigma-Aldrich St. Louis, MO, USA) is a JAK2-specific inhibitor and therefore selectively inhibits STAT3 phosphorylation. This reagent was dissolved in DMSO to a stock concentration of 80 mM and was diluted to the final concentration of 80
Tissue microarray based immunohistochemical staining
The archived paraffin blocks of 28 cases of noncancerous tumor-surrounding thyroid tissues and 19 anaplastic thyroid cancers were used to construct the tissue microarrays in the densities of 56 spots/cm
Statistical analyses
Each experiment was conducted for three times, and the data obtained were analyzed together. The results of the MTT cell proliferation assay and cell counting were evaluated with ANOVA and the independent-samples
Results
Differential resveratrol sensitivities of ATC cells
Human THJ-11T, THJ-16T and THJ-21T ATC cell lines were treated with 100
Suppressed STAT3 signaling in resveratrol-sensitive THJ-16T and THJ-21T but not in resveratrol-resistant THJ-11T cells. Cellular morphology (H/E) of resveratrol-treated THJ-11T, THJ-16T and THJ-21T cells. N, normally cultured cells; Res, 100 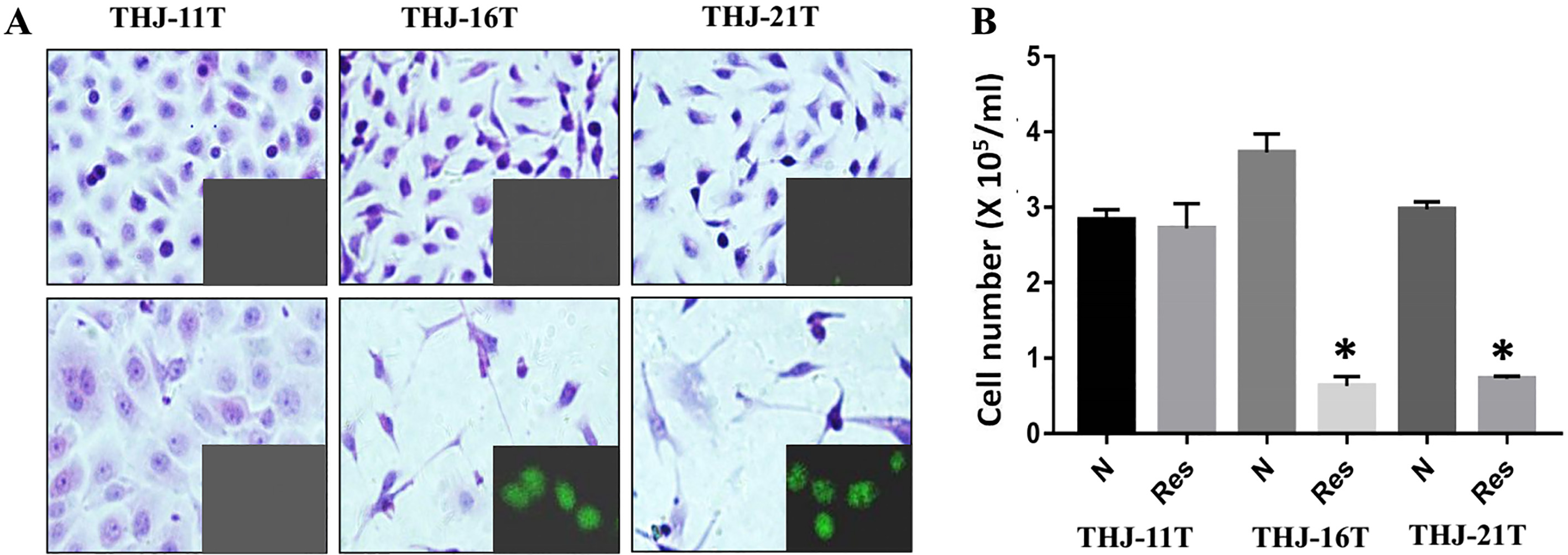
Evaluation of NF-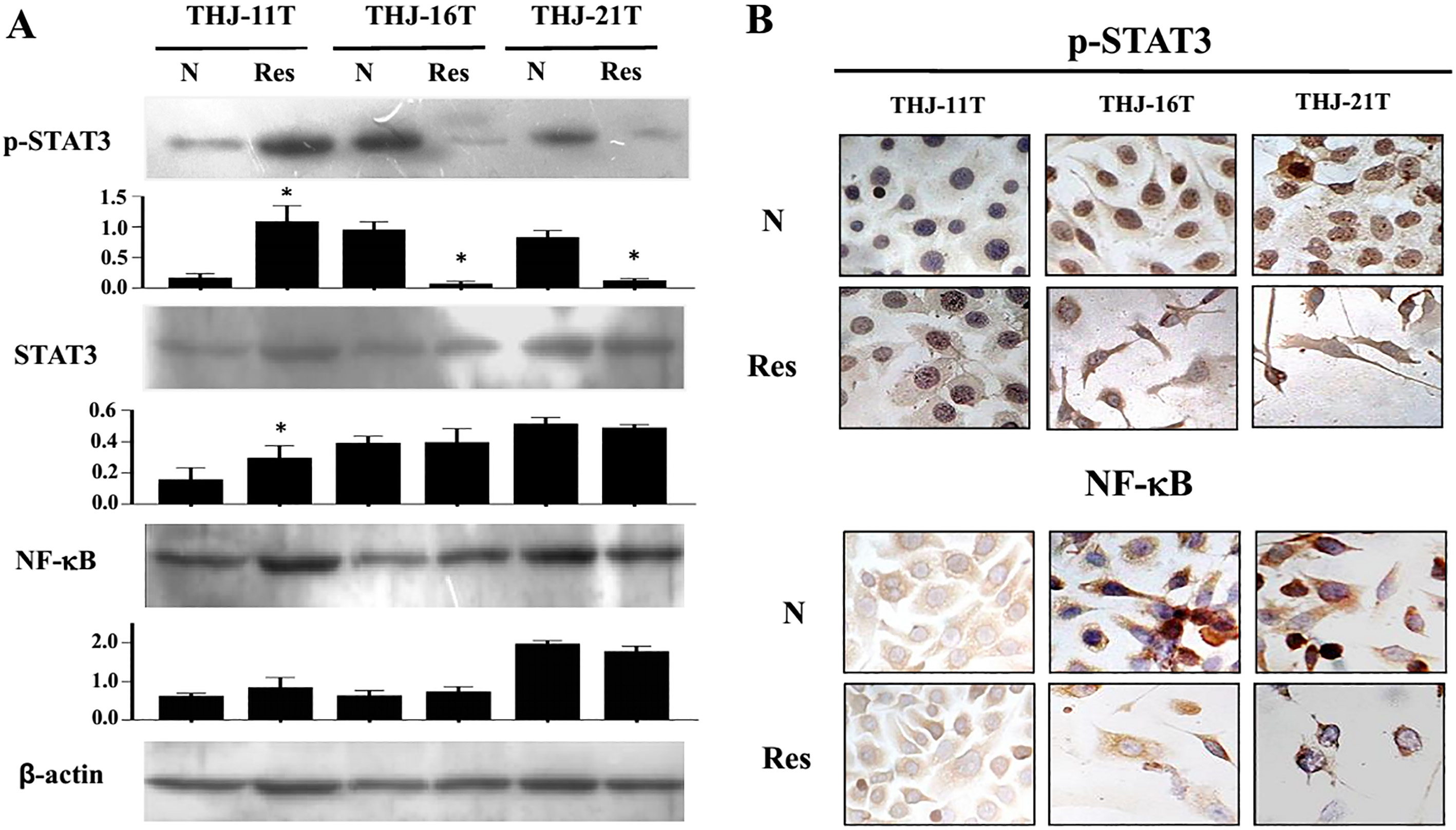
The results of Western blotting revealed that the levels of p-STAT3 but not NF-
Negative correlation of LIF expression with resveratrol sensitivity
LIF is the main activator of NF-
Evaluation of LIF expression in THJ-11T, THJ-16T and THJ-21T cells without (N) and with 48 hour 100 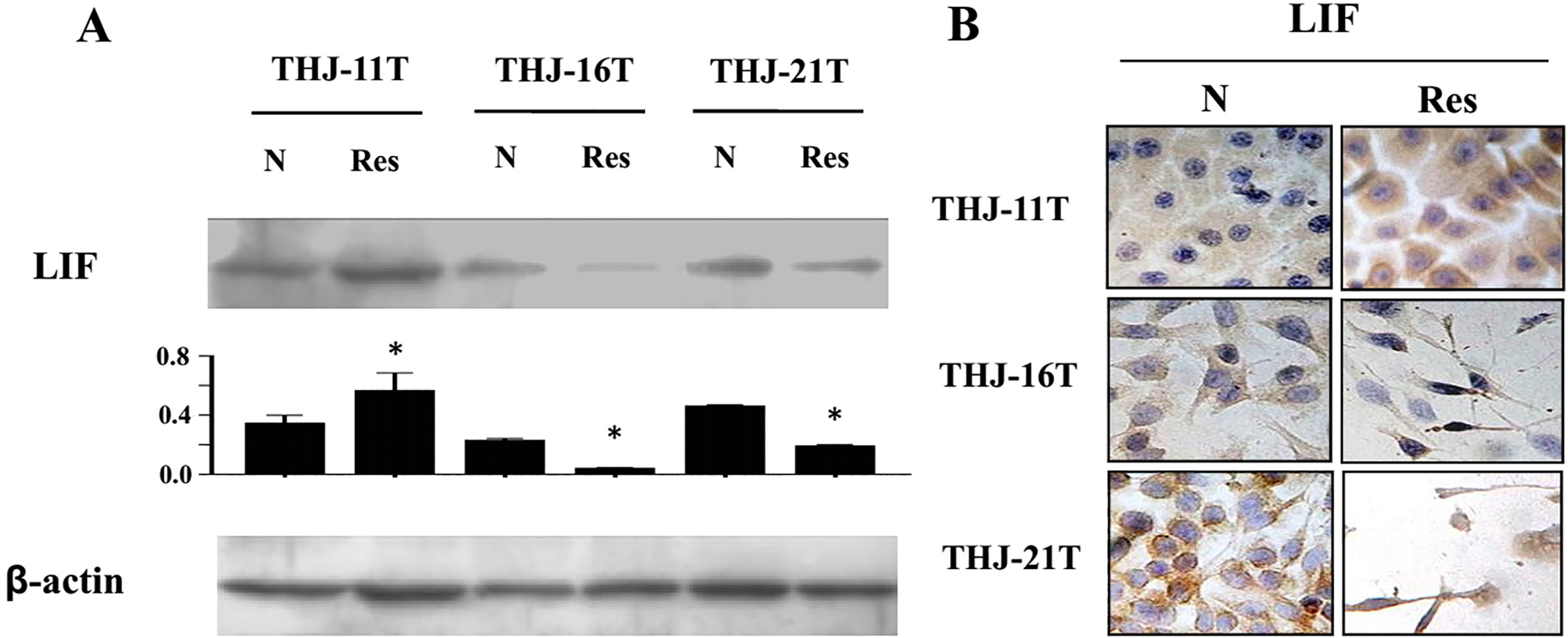
AG490 suppressed STAT3 activation and improved resveratrol sensitivity of THJ-11T cells. Immunocytochemical evaluation of p-STAT3, LIF, NF-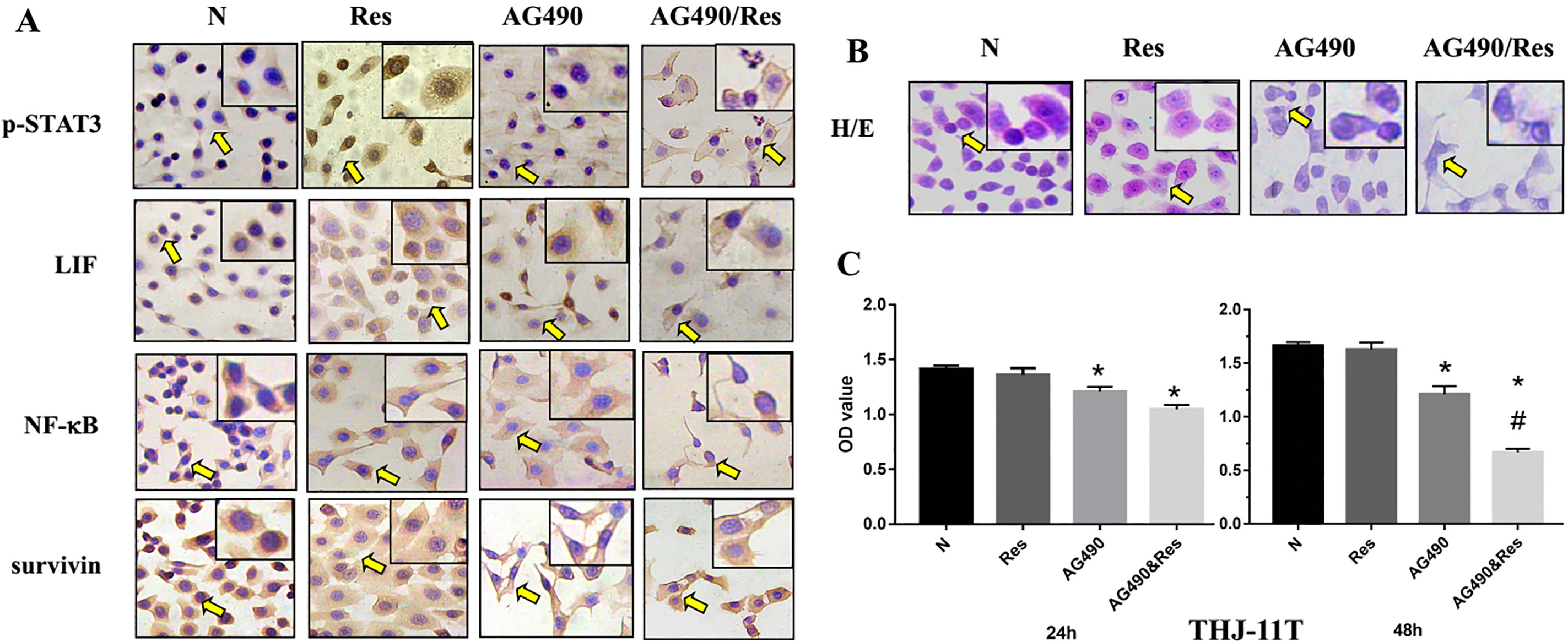
To investigate the relationship between STAT3 pathway status and resveratrol sensitivity in ATC cells, 80
Correlative analysis of p-STAT3 nuclear translocation and LIF expression in noncancerous thyroid tissues and the poorly differentiated/anaplastic thyroid cancers
Correlative analysis of p-STAT3 nuclear translocation and LIF expression in noncancerous thyroid tissues and the poorly differentiated/anaplastic thyroid cancers
#, Significant difference comparing with the data of nancancerous thyriod tissues (
Tissue microarray-based immunohistochemical profiling of p-STAT3 nuclear transloication and LIF expression in anaplastic thyroid cancers and their noncancerous counterparts. A. Immunohistochemical illustration (X 20) of p-STAT3 distribution patterns and LIF level in a case of anaplastic thyroid cancer (ATC) in the form of 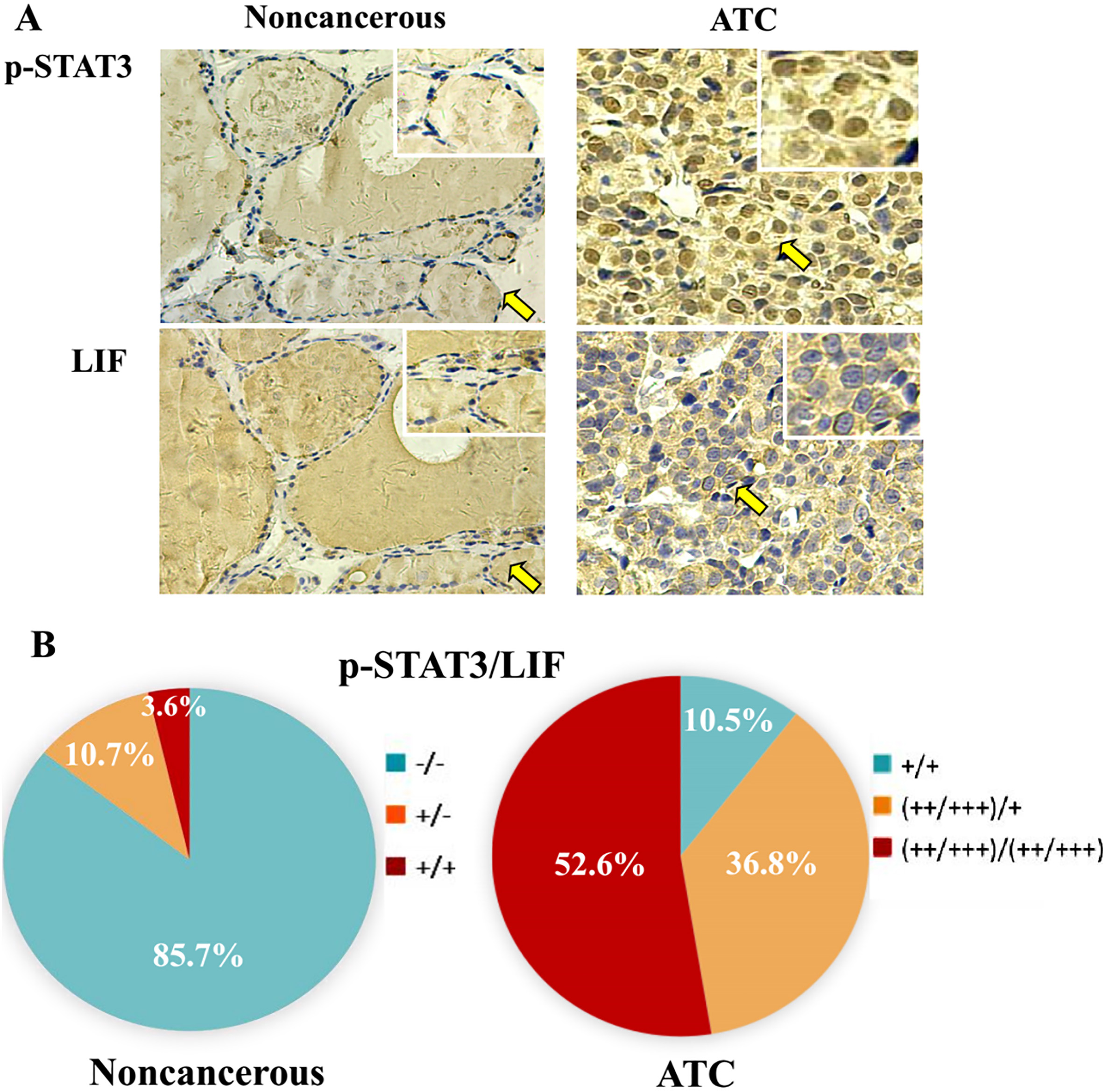
LIF-oriented immunohistochemical staining (Table 1 and Fig. 5) showed that 85.6% (24/28) of noncancerous thyroid tissues is negative in LIF expression and the remaining 14.4% (4/28) expressed LIF at a low level (
Correlation of LIF expression with STAT3 activation in ATC tissues
The positive rates of LIF and p-STAT3 in the epithelial cells of the noncancerous thyroid tissues adjacent to cancer were 14.4% and 3.6% respectively, although strong background was observed in the lumens of thyroid follicles; the positive rates of LIF and p-STAT3 in ATC tissues were 100% with higher levels of their production. The correlation analysis (Pearson) indicated the close correlation of LIF expression with the p-STAT3 nuclear translocation by showing 0.825 (
Discussion
Anaplastic thyroid cancer is the most lethal thyroid malignancy without effective chemotherapeutic remedy [4]. Resveratrol, a natural-occuring polyphenol compound, has been known as a potential chemopreventive or chemotherapeutic drug because of its extremely low toxicity to normal tissues [8], multifaceted biological activities [20] and broad anticancer spectrum [9, 21, 22]. Our recent results show that resveratrol effectively reduces the risk of carcinogen-induced rat thyroid cancer formation [23], inhibits the growth of retinoic acid-resistantg human ATC cell lines via increasing oxidative stress [24] and reverses retinoic acid-resistance of ATC cells by erasing methylation in CRABP2 promoter region [25]. Nevertheless, there are still some ATC cell lines such as THJ-11T are not sensitive to resveratrol, suggesting the necessity to investigate the internal factors leading to their resveratrol resistance for personalized therapy.
Resveratrol exerts multiple biological effects on cancer cells, of which inhibition of STAT3 activation is considered as a critical molecular event [16, 26, 27]. The activated STAT3 signaling pathway is also important for ATC cells because it regulates stem cell attributes of ATC [13] and inhibition of STAT3 activation delays the thyroid cancer formation in obese mice [28]. STAT3 pathway can be activated by IL-6-like cytokines and mediates drug resistance of thyroid cancers [2]. Given the above evidence, we supposed that the status of STAT3 signaling might be related with the anti-ATC efficacy of resveratrol. In view of the above reasons, the statuses of STAT3 pathway in resveratrol-sensitive THJ-16T and THJ-21T and -resistant THJ-11T ATC cells before and after drug treatment were analyzed. The results showed that after 48 hours after 100
AG490, a synthetic derivative of benzomethylene malonitrile, has been used as a specific inhibitor of JAK/STAT3 [31]. AG490 can inhibit the proliferation of human medullary thyroid cancers and improve their radiosensitivity [32]. To elucidate the relevance of the activated STAT3 signaling with the resveratrol resistance, THJ-11T cells were treated by 80
Because of the critical roles of STAT3 signaling in determing the response of ATC cells to resveratrol, it is necessary to investigate the upstream activator of STAT3 in THJ-11T cells before and after resveratrol treatment. Leukemia inhibitory factor/LIF is a famous member in IL-6 family, which specifically binds to gp130 receptor and activates STAT3 signaling pathway [33, 34]. We therefore checked the in vitro and in vivo expression patterns of LIF in ATCs and analyzed its correlation with activation of STAT3 pathway. In similar patterns with STAT3 alteration, 100
Taken together, our in vitro and in vivo analyses demonstrate that the status of STAT3 signaling in resveratrol-treated ATC cells is a critical element in determining the therapeutic outcome. Frequent LIF expression and its close correlation with STAT3 activation are evidenced in ATC cells and surgical specimens, which suggest a poor prognosis in ATC patients. In this context, LIF and/or IL-6 targeting therapy may improve the outcome of anti-ATC chemotherapy.
Footnotes
Acknowledgments
The authors express gratitude to Dr. Quentin Liu at the Institute of Cancer Stem Cell, Dalian Medical University, for providing the three human anaplastic thyroid cancer cell lines. This work was supported by the grants from the National Natural Science Foundation of China (No. 81272786 and 81450016 to Dr. J Liu), the Program Fund for Liaoning Provincial Department of Education Key Laboratory (LF2017002 to Dr. J Liu), and the fund from Natural Science Foundation of Liaoning Province (No. 2019-ZD-0650 to Dr. PN Li).