Triple-negative breast cancer (TNBC) is associated with an aggressive phenotype and poor prognosis, and the lack of druggable markers leads to the unavailability of targeted therapies. Thus, there is an urgent need to identify potential targets for triple-negative breast cancer.
OBJECTIVE:
In this study, we aimed to explore the expression of LAPTM4B and p27 in triple-negative breast cancer, and its clinical significance.
METHODS:
We analyzed the expression and association of LAPTM4B and p27 in human breast cancer databases. To analyze the role of LAPTM4B in the aggressiveness of the human triple-negative breast cancer, the expressions of LAPTM4B were knocked down in MDA-MB-231 and HCC1187 cell lines. Then, cell proliferation, migration and apoptosis were assessed in vitro. Furthermore, the immunohistochemistry examinations of LAPTM4B and p27 expression were performed using surgical specimens from 188 primary triple-negative breast cancer patients.
RESULTS:
Through analyses of several independent breast cancer cohorts, we found the correlation of the LAPTM4B and p27 expression. Remarkably, the knockdown of LAPTM4B restored p27 expression and inhibited the aggressiveness of breast cancer cells. Meanwhile, the knockdown of p27 relieved the suppression of cell migration. Consistent with the analyses of human breast cancer cohorts, the immunohistochemistry results showed that the expression levels of LAPTM4B and p27 were correlated in 188 triple-negative breast cancer samples ( 0.019). We also validated that the higher LAPTM4B expression, the lower p27 expression ( 0.0001), and the LAPTM4B/p27- subgroup ( 0.0001) were poor prognostic indicators, as well as the higher histologic grade ( 0.0001). In the multivariate Cox regression, p27 expression was considered as an independent predictor of survival ( 0.001).
CONCLUSIONS:
The overexpression of LAPTM4B and the loss of p27 expression are correlated. Meanwhile, the up-regulated expression of LAPTM4B together with the down-regulated expression of p27 could classified a group of breast cancer patients with poor prognosis, consequently considered as a potentially prognostic marker and candidate target for therapeutic intervention of triple-negative breast cancer.
The thorough understanding of molecular subtypes presented in breast cancer is critical to select the optimal treatment for each patient. Therefore, monitoring the valuable prognostic markers is of great importance [1]. Current clinical classification system of breast cancer often relies on the immunocytochemistry examinations of estrogen receptor (ER), progesterone receptor (PR), and HER2 protein expression [2, 3, 4, 5]. Triple-negative breast cancer is characterized by the lack of ER, PR, and HER2 protein expression. Triple-negative breast cancer is strongly associated with great invasion and metastasis potential as well as short survival, and the lack of targetable markers leading to the unavailability of targeted therapies [6]. Hence, to explore candidate targets and to gain an improved understanding of the possible pathogenesis of triple-negtive breast cancer are urgent requirements.
Lysosomal protein transmembrane 4 (LAPTM4B) gene, located in the region of chromosome 8q22.1, is composed of seven exons and six introns. LAPTM4B contains four transmembrane regions at amino acid residues 117 to 133, 163 to 179, 200 to 216, and 243 to 259 [7]. LAPTM4B encodes two isoforms, LAPTM4B-35 and LAPTM4B-24 [8]. In recent years, an increasing number of studies have found that the overexpression of LAPTM4B is associated with tumorigenesis and metastatic progression in hepatocellular carcinoma (HCC) [9], breast cancer [10, 11], ovarian carcinoma [12] and gastric cancer [13]. LAPTM4B has been reported to promote the growth of cancer cells through activating the AKT signaling pathway [8]. Yang et al. [9] found that the overexpression of LAPTM4B-35 in hepatocellular carcinoma cells could activate the AKT pathway along with the inactivation of FOXO4 (forkhead box protein O4, the transcription factor of p27), leading to the reduction in the cyclin-dependent-kinase inhibitory protein p27. p27, also known as KIP1, is encoded by the CDKN1B gene, and restrains cyclin-dependent kinases (CDKs) and cell cycle progression in G1 phase [14]. The reduction of p27 expression has been associated with poor prognosis and reduced patient survival in head and neck squamous cell carcinoma [15], lung cancer and prostate cancer [14, 16]. However, the correlation between LAPTM4B and p27 has not been fully investigated in breast cancer, as well as the prognostic significance of LAPTM4B and p27 expression.
In this study, we analyzed LAPTM4B gene expression across several independent breast cancer cohorts. Our results revealed a remarkable overexpression of LAPTM4B and a negative correlation between LAPTM4B and p27 expression in triple-negative breast cancer. Furthermore, we demonstrated that the knockdown of LAPTM4B increased p27 expression and impaired the growth and migration of human triple-negative breast cancer cells MDA-MB-231 and HCC1187 in vitro. To understand the clinical significance of LAPTM4B and p27 expression, we performed a retrospective study in 188 triple-negative breast cancer patients. Consistently, our clinical data confirmed that the overexpression of LAPTM4B was accompanied by the loss of p27 expression, which was regarded as a prognostic predictor of survival, as well as a candidate druggable target for triple negative breast cancer.
Materials and methods
Samples and clinicopathological data
A total of 188 surgically resected primary triple-negative breast cancer tumors, six benign breast tumors and six normal breast tissues were collected at the Second Hospital of Dalian Medical University between January 2009 and January 2013. The inclusion criteria and follow-up were performed as previously described [5]. All patients provided informed consents. The experimental protocol was approved by the Dalian Medical University Ethics Committee, conformed with The Code of Ethics of the World Medical Association (Declaration of Helsinki), printed in the British Medical Journal (18 July 1964).
Immunohistochemistry
All clinical samples were fixed in neutral formalin before embedded in paraffin. The expression of LAPTM4B and p27 was assessed following a two-step method. Rabbit anti-LAPTM4B polyclonal antibody (1: 100) and rabbit anti-p27 polyclonal antibody (1: 200) were purchased from Abclonal (DF12416) and Proteintech (AF6324), China. The DAB kit was purchased from Zhongshan Goldenbridge Biotechnology Company (Beijing, China). Negative controls were detected by replacing the primary antibodies with normal goat serum. Three experienced pathologists blinded to patient details evaluated all samples. The positive staining was defined as samples displaying clear brown granules in the cytoplasm and nucleus, respectively. The percentage and the staining intensity of positive cells were evaluated as previously described [5].
Cell culture and siRNA transfection
The MDA-MB-231 and HCC1187 cell lines were purchased from the American Type Culture Collection (ATCC) and cultured in DMEM medium (Gibco, USA) and RPMI-1640 medium (Gibco, USA) supplemented with 10% fetal bovine serum (Gibco, USA), respectively. Cells were incubated at 37C in a 5% CO2-containing atmosphere. All plates were purchased from NEST, China. Gene-specific and negative control oligonucleotides (siRNA, GenePharma, China) were transfected using the LipofectamineTM RNAi MAX protocol from Invitrogen. The effectiveness of siRNA knockdown was determined by western blot.
Immunoblotting and antibodies
Total proteins of cell lines were lysed with cold RIPA buffer (pH 7.4, 150 mM NaCl, 25 mM Tris, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate) supplemented with protease inhibitor (Roche). Subsequently, the lysates were measured for protein concentration using BCA kit (Tiangen, China). Equal amounts of proteins (40 ug per conditio) were separated by SDS-PAGE and then transferred to nitrocellulose (NC) membranes (Bio-RAD). After blocking with 5% non-fat milk, and the NC membranes were incubated with primary antibodies overnight at 4C, including LAPTM4B (Abclonal, China), p27 (Proteintech, China), Tubulin (Abclonal, China). After washes (TBS-T), the NC membrane was incubated with fluorophore-conjugated secondary antibodies for one hour at room temperature. The NC membrane was washed again and visualized with the Odyssey Scanner (Li-Cor, Bioscience, Lincoln, NE, USA).
Wound-healing assay and apoptosis assay
Cells were proliferated in 12-well plate (NEST, China) and scratch wound was made by 200 l pipette tip after starving overnight in the serum-free medium. The cells were washed twice with PBS to remove cell debris. Wound was observed after 0 and 24 h in the serum-free medium. The widths of scratch wound were measured under microscope (Leica). Cultured cells were treated with siControl or siLAPTM4B for 48 h, and then analyzed by the Annexin V Apoptosis Detection Kit (Biosea Biotechnology, Beijing, China) with a FACSCalibur flow cytometer.
Statistical analysis
The TCGA data were downloaded from www. cbioportal.com (cBioPortal). Two-tailed Student’s t-test and analysis of variance (ANOVA) were carried out to compare between two groups and among three groups, respectively. The Fisher’s exact test was used to analyze the relationship between LAPTM4B and p27 expression. Survival data were analyzed using univariate analysis. The significant variables ( 0.05) from the univariate regression were entered into a forward conditional Cox regression to identify the independent variables. We evaluated the capability of the combined groups or each individual protein to predict prognosis by measuring the area under receiver operator characteristic (ROC) curves. Kaplan-Meier curve and the hazard ratio were evaluated using Prism 6. The receiver operator characteristic (ROC) curves were evaluated using SPSS 17. 0.05 was considered statistically significant.
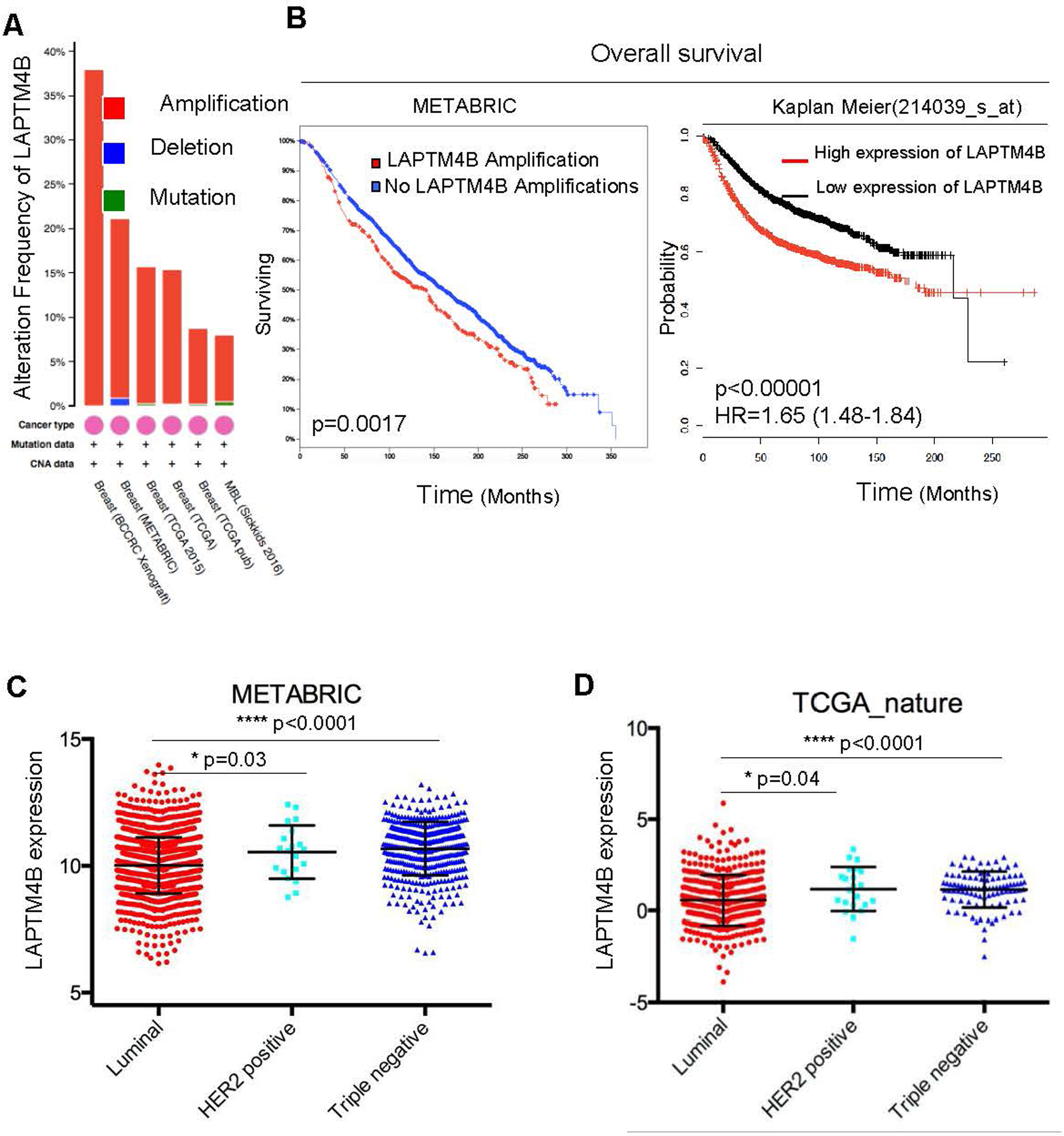
LAPTM4B is overexpressed in triple-negative breast cancer and predicts poor prognosis. (A) The data is analyzed by cBioPortal. (B) Samples were divided into two groups with and without LAPTM4B gene amplifications. The overall survival curve was analyzed by cBioPortal (left). Samples were divided into top 50% ‘LAPTM4B high’ and bottom 50% ‘LAPTM4B low’ groups based on the mRNA expression of LAPTM4B. Kaplan–Meier curve of overall survival was analyzed by the online analytical system (http://kmplot.com) (right). (C and D) The LAPTM4B mRNA expression among three subtypes of breast cancer. In the dot plots, every dot indicated an individual patient. Black lines in each group indicate median with interquartile range.
Relevance of LAPTM4B and p27 expression with clinicopathological features
Variable
Patients
LAPTM4B
value
p27
value
Negative
Positive
Negative
Positive
Age(year) – no. (%)
50
94(50%)
41(22)
53(28)
0.465
50(27)
44(23)
0.884
50
94(50%)
46(24)
48(26)
51(27)
43(23)
No. of metastatic axillary nodes – no. (%)
0
42(22%)
16(8)
26(14)
0.098
22(12)
20(10)
0.115
1–3
90(48%)
49(26)
41(22)
42(22)
48(26)
3
56(30%)
22(12)
34(18)
36(19)
20(11)
Diameter of primary tumor – no. (%)
30 mm
104(55%)
48(26)
56(30)
0.900
51(27)
53(28)
0.152
30 mm
84(45%)
38(20)
46(24)
50(27)
34(18)
Histologic grade – no. (%)
1
29(15%)
11(6)
18(10)
0.635
15(8)
14(8)
0.926
2
116(62%)
54(29)
62(33)
63(34)
53(28)
3
43(23%)
21(11)
22(11)
22(11)
21(11)
TNM staging
I
36(19%)
18(10)
18(10)
0.0194
20(11)
16(9)
0.446
II
78(41%)
41(22)
37(19)
39(21)
39(21)
III
50(27%)
23(12)
27(14)
24(12)
26(14)
IV
24(13%)
4(2)
20(11)
16(8)
8(4)
Correlation between the expression of LAPTM4B and p27
LAPTM4B
p27
Fisher’s exact test
Negative ( 87)
Positive ( 101)
-value
Negative ( 102)
39 (21%)
63 (33%)
0.019
Positive ( 86)
48 (26%)
38 (20%)
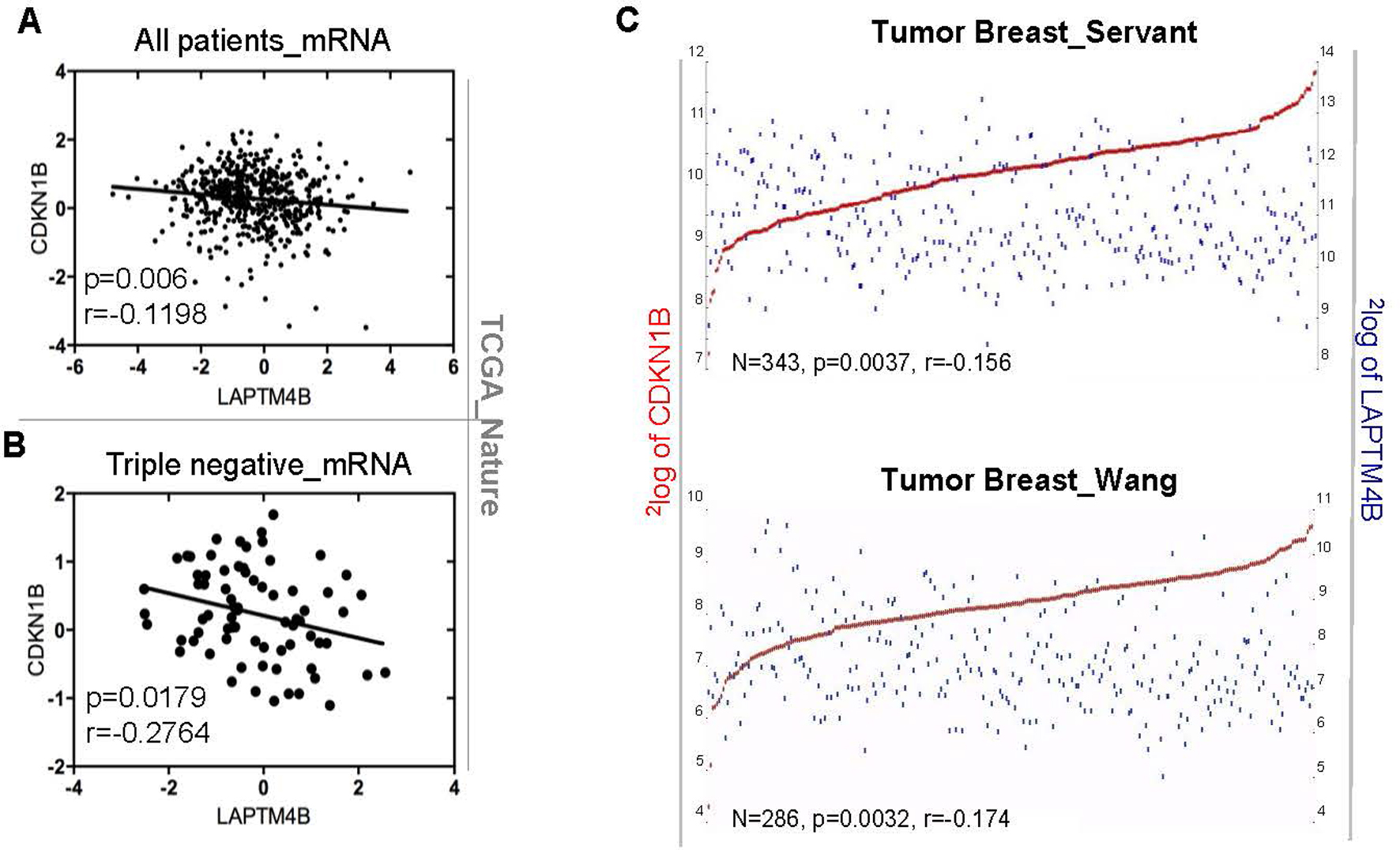
Correlation of LAPTM4B and p27 expression. (A and B) The linear regression of LAPTM4B and CDKN1B mRNA expression levels in all breast cancer patients or TNBC was analyzed across the TCGA database. (C) The correlation of LAPTM4B and CDKN1B was analyzed by the online analytical system R2 (https://hgserver1.amc.nl/cgi-bin/r2/main.cgi).
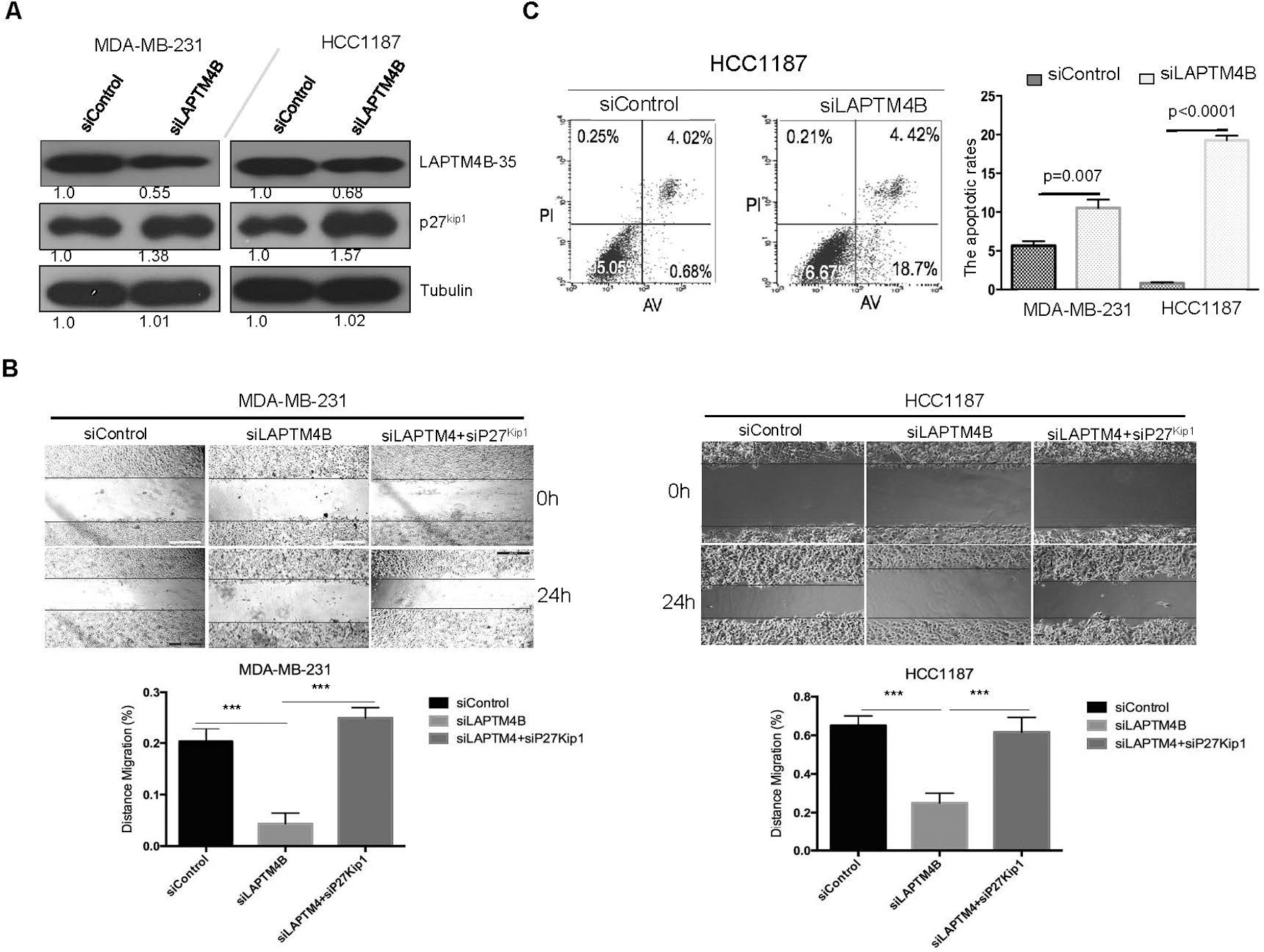
Knockdown of LAPTM4B restores p27 expression and inhibits breast cancer cell aggressiveness. (A) Immunoblotting analyses of proteins as indicated in siControl and siLAPTM4B transduced MDA-MB-231 and HCC1187 cell lines. (B) The migration capacity of MDA-MB-231 and HCC1187 was assessed by wound-healing assays. ***, 0.001. (C) Flow cytometry assays were performed to analyze the apoptosis of MDA-MB-231 and HCC1187 cells transfected with siControl and siLAPTM4B as indicated.
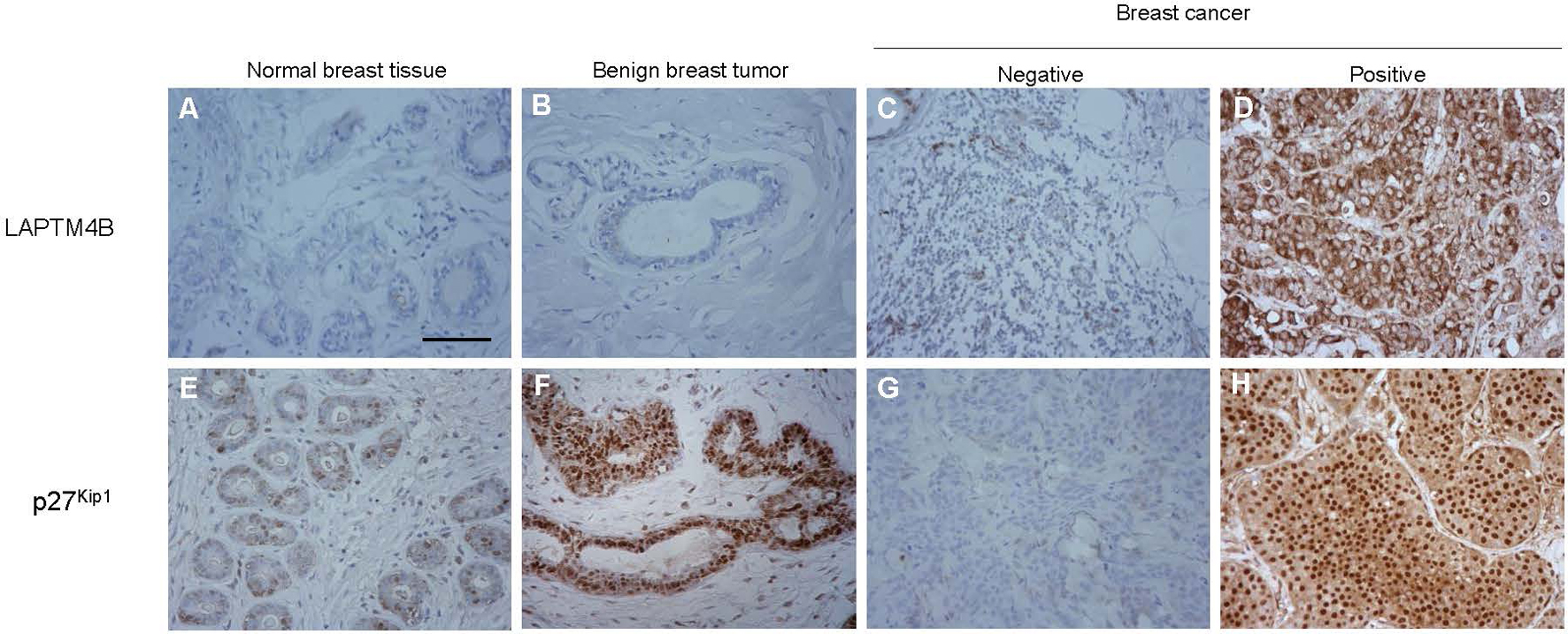
Evaluation of Immunohistochemistry staining for LAPTM4B and p27. Notes: (A, E) and (B, F) represented immunohistochemistry staining for normal breast tissue and benign breast tumor. (C, D, G, H) represented immunohistochemistry staining of breast cancer as indicated. Scar bars, 200 m.
Univariate and Multivariate analysis of overall survival
End point
No. of patients
Univariate analysis
Multivariate analysis
Hazard ratio (95% CI)
-value
Hazard ratio (95% CI)
-value
Age ( 50 years vs 50 years)
94
94
0.799 (0.468–1.040)
0.0833
n.a.
n.a.
LAPTM4B (positive vs negative)
101
87
1.662 (1.131–2.508)
0.0120
–
0.545
p27 (positive vs negative)
87
101
0.264 (0.163–0.365)
0.0001
4.107 (2.589–6.517)
0.001
Metastatic axillary nodes ( 3 VS 3)
82
106
1.104 (0.743–1.654)
0.6197
n.a.
n.a.
Diameter of primary tumor
84
104
1.082 (0.728–1.619)
0.6917
n.a.
n.a.
( 30 mm VS 30 mm)
Histologic grade (3 vs 1/2)
43
145
34.38 (13.87–80.28)
0.0001
–
0.985
TNM staging (III/IV vs I/II)
74
114
1.628 (1.126–2.597)
0.0137
–
0.157
Note: n.a. Not application.
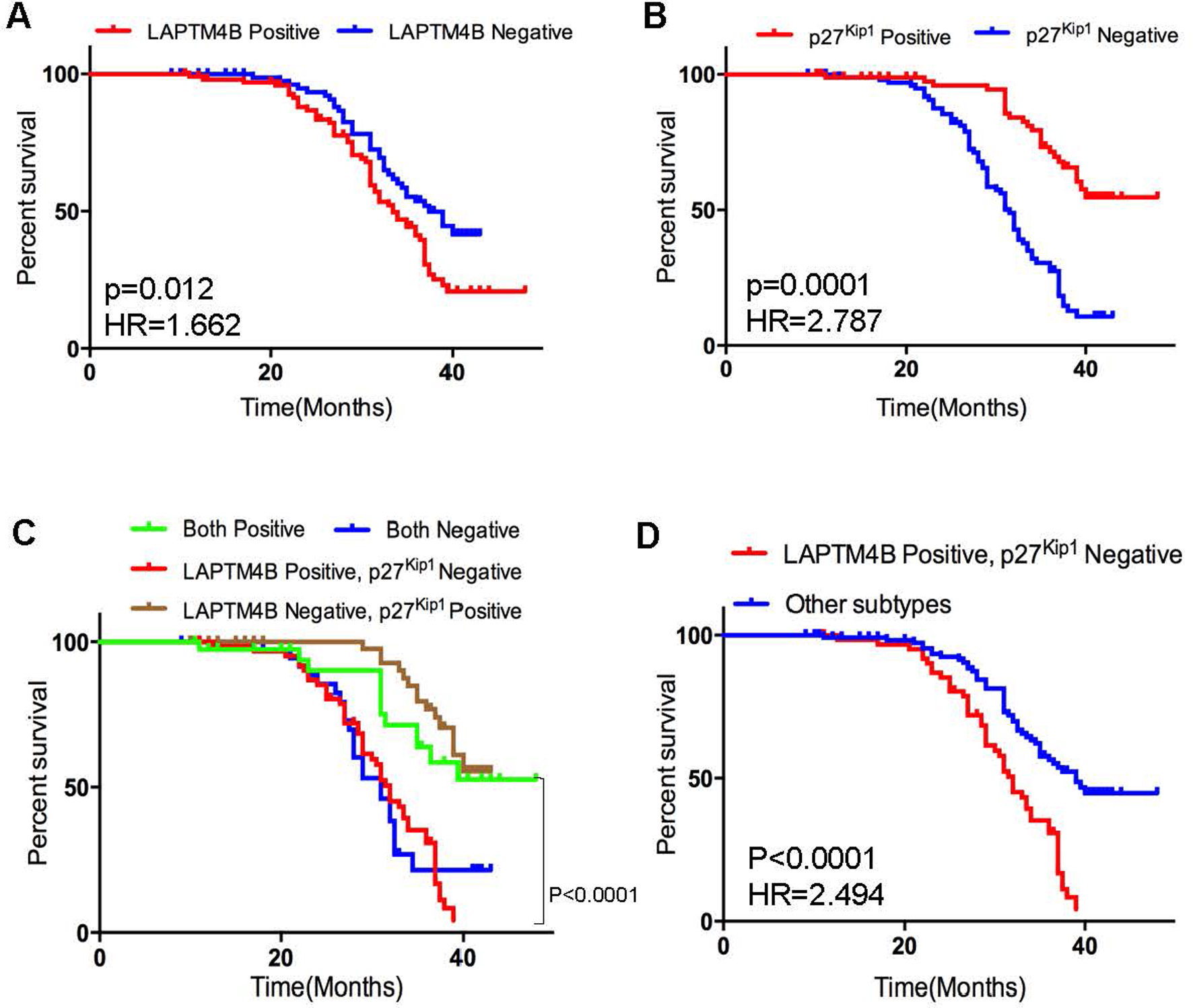
Kaplan-Meier analysis of overall survival in triple negative breast cancer. (A, B) Samples were divided into two groups with positive and negative expression of LAPTM4B and p27. (C, D) Kaplan-Meier curves of overall survival in different subgroups. -values are obtained from the log-rank test. Hazard ratio (HR) is calculated using GraphPad Prism.
Results
The recurrent LAPTM4B overexpression predicts poor prognosis
We analyzed the amplification of LAPTM4Bgene in the breast cancer data retrieved from cBioPortal and observed that the LAPTM4Bgene amplification is recurrent (Fig. 1A). We next assessed the prognostic significance of LAPTM4B gene amplification in the METABRIC breast cancer cohort, with results suggesting that cases with amplifications showed a worse prognosis of overall survival than those without amplification ( 0.0017) (Fig. 1B, left). To gain more insights whether LAPTM4B mRNA expression levels can also predict prognosis, patients were divided into top 50% ‘LAPTM4B high’ and bottom 50% ‘LAPTM4B low’ groups based on the LAPTM4B mRNA levels. By using the web-based analysis tool (http://kmplot.com), we drew a similar conclusion that ‘LAPTM4B high’ group showed a worse prognosis of overall survival ( 0.00001, HR 1.65) (Fig. 1B, right). Our findings indicated that the high expression levels of LAPTM4B at both gene and mRNA levels were correlated with poor prognosis in breast cancer.
LAPTM4B is overexpressed in triple-negative breast cancer
Furthermore, we analyzed the LAPTM4B mRNA expression in three subtypes of breast cancer across two independent cohorts, METABRIC and TCGA_Nature. We observed strikingly similar patterns that the TNBC displayed the highest expression levels of LAPTM4B (Fig. 1C and D). Together, these data indicated that the TNBC showed the highest expression levels of LAPTM4B when comparing with other subtypes.
Correlation between LAPTM4B and p27 expression
Previous studies [9] reported that the overexpression of LAPTM4B-35 in hepatocellular carcinoma cells played a critical role in the growth and metastasis of HCC, and inactivated FOXO4, leading to the reduction of cell cycle inhibitory protein p27. However, little is known about the involvements of LAPTM4B and p27 in the occurrence and development of breast cancer. To investigate whether the expression of LAPTM4B and p27 was correlated at the mRNA level, we analyzed gene expression data across the TCGA database and the online analytical system R2 (Genomics Analysis and Visualization Platform, https://hgserver1.amc.nl/cgi-bin/r2/main.cgi). We found that LAPTM4B expression was negatively correlated with CDKN1B expression in breast cancer (Fig. 2A, C and Table S1). The correlation of LAPTM4Band CDKN1B was also confirmed in triple-negative breast cancer (Fig. 2B and Table S1). Taken together, the results from our analyses showed that the overexpression of LAPTM4B was correlated with loss of p27 in breast cancers.
Knockdown of LAPTM4B restores p27 expression and inhibits breast cancer cell aggressiveness
To test the relevance of LAPTM4B to p27, we assessed the effect of LAPTM4B knockdown on p27 expression in vitro (Fig. 3A). The knockdown of LAPTM4B resulted in the elevated levels of p27 in the human triple-negative breast cancer cell line MDA-MB-231 and HCC1187 (Fig. 3A). We next confirmed the effect of LAPTM4B on cell migration using the wound-healing assay. LAPTM4B knockdown caused an apparent suppression of cell migration in MDA-MB-231 and HCC1187 cells (Fig. 3B). The additional knockdown of p27 rescued the suppression of MDA-MB-231 and HCC1187 migration (Fig. 3B), suggesting that p27 was responsible for the LAPTM4B effects on cell migration. Furthermore, to investigate the effect of LAPTM4B on apoptosis, the apoptotic rates of siRNA-treated MDA-MB-231 and HCC1187 cells were analyzed by flow cytometry, and similar apoptotic responses were observed in two cell lines. The apoptotic rate of siLAPTM4B cells was significantly increased compared with the siControl groups (Fig. 3C). These results demonstrated that the knockdown of LAPTM4B restored p27 protein levels, which inhibited breast cancer cell aggressiveness.
Clinicopathological relevance of LAPTM4B and p27 expression
To investigate the clinical relevance of LAPTM4B and p27 expression, we measured the LAPTM4B and p27 protein expression in 188 TNBC tissues with immunohistochemistry (Fig. 4). The median survival time of all patients was 32 months. Ninety-seven patients died during follow-up, and all causes of death were cancer-related. No significant statistical association of LAPTM4B or p27 expression was observed with age, number of metastatic axillary nodes, diameter of primary tumor, or histologic grade (Table 1). However, the expression of LAPTM4B was significantly associated with TNM staging ( 0.0194). We next investigated the correlation between LAPTM4B and p27 expression at the protein level. As shown in Table 2, consistent with the aforementioned database analyses (Figs 1 and 2), our results also showed a correlation between the levels of LAPTM4B and p27 expression ( 0.019). In addition, we performed the univariate Cox regression analyses to evaluate the clinical factors (Table 3). LAPTM4B expression ( 0.012, HR 1.662) (Fig. 5A), p27 expression ( 0.0001, HR 0.264) (Fig. 5B), histologic grade ( 0.0001, HR 34.38), and TNM staging ( 0.0137, HR 1.628) were significantly correlated with overall survival in the univariate Cox regression analysis (Table 3). Subsequently, patients were divided into the following four subgroups based on the expression levels of these two proteins based on immunohistochemical analysis, namely LAPTM4B/p27 co-expression, both negative, LAPTM4B positive/p27 negative, LAPTM4B negative/p27 positive. Interestingly, the survival time of patients with both negative and LAPTM4B positive/p27 negative was significantly shorter than the others ( 0.0001, Fig. 5C). The LAPTM4B positive/p27 negative subtype also showed poor prognosis as compared with the other patients ( 0.0001, Fig. 5D). However, according to the multivariate Cox regression analysis, p27 expression, not LAPTM4B expression, was considered as an independent predictor of survival ( 0.001, Table 3).
Discussion
In this study, through a comprehensive analysis of databases, we found LAPTM4B to be highly expressed in TNBC. The association of the expression of LAPTM4B and p27 in breast cancer was also found in multiple breast cancer cohorts. Remarkably, the knockdown of LAPTM4B restored p27 expression and inhibited breast cancer cell aggressiveness in vitro. Even more striking is the clinical findings that the higher LAPTM4B expression, the lower p27 expression ( 0.0001), and the LAPTM4B/p27- subgroup ( 0.0001) were poor prognostic indicators in 188 triple-negative breast cancer patients. However, the p27 expression, not the LAPTM4B expression, was considered as an independent indicator of survival in the multivariate Cox regression (Table 3, 0.001).
Previous investigations [14, 15, 16, 17] also reported that the reduction of p27 expression was associated with reduced patient survival in head and neck squamous cell carcinoma, lung cancer, prostate cancer and breast cancer. In this study, we concluded that p27 could serve as an independent prognostic factor to predict patient overall survival in triple-negative breast cancer patients (Table 3). By combining the analyses of LAPTM4B and p27 expression at the mRNA and protein levels, we showed an association between LAPTM4B and p27 expression (Fig. 2). The knockdown of LAPTM4B increased p27 expression, which provided us further in vitro proofs of their association. The immunohistochemical analyses of 188 TNBC patients also showed similar results (Table 2, 0.019). However, through the analyses of the METABRIC cohort, we did not observe the significant correlation between the expression of LAPTM4B and loss of p27 in METABRIC cohort (Fig. S1). We acknowledged that there were limitations that not all databases supported our conclusions in this study.
The LAPTM4B positive/p27 negative and both negative groups showed significant statistical differences as compared with the other two groups (Fig. 5C and D). Subsequently, we performed the receiver operating characteristic (ROC) curve analysis (Fig. S2) and the multivariate Cox regression (Table S2). The AUC (Area Under Curve) of LAPTM4B positive and p27 negative group (AUC 0.670, 95% CI 0.593–0.748) was larger than the other groups (Fig. S2). Indeed, the overexpression of LAPTM4B with loss of p27 was a potential prognostic indicator of triple-negative breast cancer. Further studies are required to unravel the mechanisms how the LAPTM4B and p27 expression promotes disease progression. To the best of our knowledge, our current study provides one of the most comprehensive reports on the correlation of LAPTM4B and p27 expression in TNBC.
Li et al. [18] reported that LAPTM4B was a therapeutic target for sensitizing tumors to chemotherapy. On the other hand, Berton et al. [19] provided insights into the mechanisms that the loss of p27 increased genomic instability and induced radio-resistance in breast cancer. In addition, Liu et al. [20] showed that TRIM62 suppressed tumor metastasis via regulating cell cycle related protein P27 by targeting c-Jun N-terminal kinase. However, these findings did not fully elucidate the implications of LAPTM4B and p27 expression in tumor metastasis and therapeutic responses. To be honest, we have to acknowledge the limitations of our study was due to the small sample size and a retrospective analysis. Additionally, the inadequate number of positive samples in different subtypes could also reduce the statistical power in data analysis. To further examine the oncogenic role of LAPTM4B and p27 in TNBC, much deeper research is required to gain more mechanistic insights into why triple-negative breast cancers expressing LAPTM4B with low levels of p27 expression resulting in a worse overall prognosis. In this way, the TNBC cells stably overexpressing/knocking-down LAPTM4B or p27 need to be generated to investigate the other effects of LAPTM4B and p27 on TNBC cells. The test in vivo should also be performed to exam the relationship between the expression of LAPTM4B and/or p27 and the proliferation and metastasis ability, as well as chemotherapy resistance of TNBC.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (NO. 81471751, and NO. 81673762 to Zuowei Zhao; No. 81650018 to Man Li), and Provincial Natural Science Foundation of Liaoning (NO. 2014921059 to Zuowei Zhao).
Conflict of interest
The authors declare no conflict of interest.
Supplementary data
The supplementary files are available to download from http://dx.doi.org/10.3233/CBM-182094.
References
1.
FriedmanAA.LetaiA.FisherD.E. et al., Precision medicine for cancer with next-generation functional diagnostics, Nat Rev Cancer15(12) (2015), 747–756.
2.
Brasó-MaristanyF.FilostoS.CatchpoleS. et al., PIM1 kinase regulates cell death, tumor growth and chemotherapy response in triple-negative breast cancer, Nat Med22(11) (2016), 1303–1313.
3.
FoulkesW.D.SmithI.E. and Reis-FilhoJ.S., Triple-negative breast cancer, N Engl J Med363(20) (2010), 1938–1948.
4.
RakhaE.A.Reis-FilhoJ.S. and EllisI.O., Basal-like breast cancer: A critical review, J Clin Oncol26(15) (2008), 2568–2581.
5.
LiX.SunS.LiN. et al., High expression of ccr7 predicts lymph node metastasis and good prognosis in triple negative breast cancer, Cell Physiol Biochem43 (2017), 531–539.
6.
DietzeE.C.SistrunkC.Miranda-CarboniG. et al., Triple-negative breast cancer in African-American women: disparities versus biology, Nat Rev Cancer15(4) (2015), 248–54.
7.
LiY.ZouL.LiQ. et al., Amplification of LAPTM4B and YWHAZ contributes to chemotherapy resistance and recurrence of breast cancer, Nat Med16(2) (2010), 214–218.
8.
MengY.WangL.ChenD. et al., LAPTM4B: an oncogene in various solid tumors and its functions, Oncogene35(50) (2016), 6359–6365.
9.
YangH.XiongF.WeiX. et al., Overexpression of LAPTM4B-35 promotes growth and metastasis of hepatocellular carcinoma in vitro and in vivo, Cancer Lett294(2) (2010), 236–244.
10.
XiaoM.JiaS.WangH. et al., Overexpression of LAPTM4B: an Independent prognostic marker in breast cancer, J Cancer Res Clin Oncol139(4) (2013), 661–667.
11.
XiaoM.YangS.MengF. et al., LAPTM4B Predicts Axillary Lymph Node Metastasis in Breast Cancer and Promotes Breast Cancer Cell Aggressiveness in Vitro, Cell Physiol Biochem41(3) (2017), 1072–1082.
12.
YinM.LouC.ZhangW. et al., LAPTM4B overexpression is a novel independent prognostic marker for metastatic ovarian tumors, Int J Gynecol Cancer22(1) (2012), 54–62.
13.
LiuY.ZhangQ.Y.QianN. et al., Relationship between LAPTM4B gene polymorphism and susceptibility of gastric cancer, Ann Oncol18(2) (2007), 311–316.
14.
ChuI.M.HengstL. and SlingerlandJ.M., The CDK inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapy, Nat Rev Cancer8(4) (2008), 253–267.
15.
CiesielskaU.ZatonskiT.NowinskaK. et al., Expression of cell cycle-related proteins p16, p27 and ki-67 proliferating marker in laryngealsquamous cell carcinomas and in laryngeal papillomas, Anticancer Res37(5) (2017), 2407–2415.
16.
RaghuD.PaulP.J.GulatiT. et al., E6AP promotes prostate cancer by reducing p27 expression, Oncotarget8(26) (2017), 42939–42948.
17.
BarnesA.PinderS.E.BellJ.A. et al., Expression of p27kip1 in breast cancer and its prognostic significance, The Journal of Pathology201(3) (2003), 451–459.
18.
LiY.ZhangQ.TianR. et al., Lysosomal transmembrane protein LAPTM4B promotes autophagy and tolerance to metabolic stress in cancer cells, Cancer Res71(24) (2011), 7481–7489.
19.
BertonS.CusanM.SegattoI. et al., Loss of p27kip1 increases genomic instability and induces radio-resistance in luminal breast cancer cells, Sci Rep7(1) (2017), 595.
20.
LiuT.Y.ChenJ.ShangC.L. et al., Tripartite motif containing 62 is a novel prognostic marker and suppresses tumor metastasis via c-Jun/Slug signaling-mediated epithelial-mesenchymal transition in cervical cancer, J Exp Clin Cancer Res35(1) (2016), 170.