Colorectal cancer (CRC) is the most common malignant disease worldwide and thus new therapeutic approaches are needed. 5-Fluorouracil (5-FU) remains the most widely used agent to treat colorectal cancer (CRC). However, its clinical efficacy is currently limited by the development of drug resistance. Esculetin (EST), a coumarin, was found to have anti-proliferative and anti-migration activity in cancer.
OBJECTIVE:
This research aims to evaluated the influence and possible mechanism of EST on the proliferation, migration and epithelial-mesenchymal transition of CRC cell lines.
MATERIALS AND METHODS:
Human CRC cell lines HT-29, SW480, HCT-116, and Caco-2 were treated with various concentrations of EST (0.2, 2, 20, 200, 2000 g/ml) or 5-FU (0.1, 1, 10, 100, 1000 g/ml) for 48 h, and cell viability was determined by the MTT and CCK-8 assay. The motility of HCT-116 cells was detected by scratch assay. Western blot was applied to detect the protein expression. Besides, levels of Wnt3a and VEGF in HCT-116 cell culture medium supernatant were analyzed by ELISA. The anti-tumor effect was detected with HCT-116 subcutaneous tumor bearing tumor model by monitoring the tumor vomume in vivo. Finally, the tumoral expression of VEGF was measured by immunohistochemistry, and the expression of Ki67, PCNA, -catenin, c-Myc, Cyclin D1, MMP2 and MMP7 was measured by Western blot analysis.
RESULTS:
EST inhibited HCT-116 cell proliferation in a dose-dependent manner. Western blot analysis revealed that EST decreased the expression of Ki67, PCNA, N-cadherin, E-cadherin, vimentin, fibronectin, -catenin, c-Myc, Cyclin D1, MMP2 and MMP7. Furthermore, EST reduced the release of Wnt3a and VEGF into HCT-116 cells culture medium. After EST treatment, the tumor volume was significant smaller than that of the control group, and the tumoral levels of VEGF were decreased. Moreover, western blot analysis indicated that the expression of Ki67, PCNA, -catenin, c-Myc, Cyclin D1, MMP2 and MMP7 were also significantly decreased after treated with EST. In addition, in vitro and in vivo anti-tumor results demonstrated that EST combined with 5-FU could increase the inhibitory effect of 5-FU on HCT-116 cells proliferation, migration and epithelial-mesenchymal transition.
CONCLUSIONS:
EST enhances the inhibitory effect of 5-FU on the proliferation, migration and epithelial-mesenchymal transition of CRC.
Colorectal cancer (CRC) is the third most commonly diagnosed cancer in males and the second most frequent cancer reported in females [1]. Colon cancer is a major cause of cancer incidence and mortality worldwide,which is one of the most commonly diagnosed malignant cancers clinically [2, 3, 4], limited by current screening and diagnostic techniques, most of the CRC was diagnosed in the late stage, so its pathogenesis and chemo-prevention are areas of intense investigation [5, 6]. Possible mechanisms of the anti-proliferative properties of current chemotherapeutic colon cancer agents include inhibitory effects on DNA synthesis, the induction of apoptosis, modulation of cell cycle control, and the regulation of signal transduction pathways [7, 8, 9, 10]. As a fact of that, it is of great scientific significance and clinical value to study novel therapies of CRC.
Epithelial-mesenchymal transition (EMT) plays a leading role in tumor progression, invasion, and metastasis, leading to poor prognosis and resistance [11, 12]. Triggered EMT promotes the transfer of polarized epithelial cells from the primary site to the secondary site, while inducing biological changes such as mesenchymal phenotype, lamellar foot and invasive foot [13, 14]. E-cadherin is an adhesion molecule at the junction of cells and is becoming a therapeutic target for inhibiting EMT in cancer cells [15]. Furthermore, in the majority of colon cancers, the canonical Wnt/-catenin pathway is constitutively active [16, 17, 18, 19], which leads to a disruption of the -catenin destruction complex and accumulation of -catenin. Multiple mutations lead to the nuclear accumulation of -catenin [20, 21]. This leads to the inappropriate activation of its target genes, including c-myc [22] and cyclin D1 [23, 24], and also plays an essential role in proliferation of colon cancer cells.
However, a number of studies have demonstrated that widely used, naturally-occurring compounds are more effective and safer chemopreventive agents, in comparison to the severe toxicity and side-effects associated with current chemoth-erapeutic agents for colon cancer [25]. EST is a commonly used traditional Chinese medicine that was first recorded in Shennong’s Herbal Classic. Modern pharmacological research has shown that EST has pharmacological effects of antimicrobial, anti-inflammatory and anti-tumor [26]. Moreover, several current studies reveal that EST could function as an immunomodulator, with strong inhibition of malignant cancer including hepatic cancer [27], lung cancer [28, 29] and leukemia [30]. Nevertheless, the field of this research on the functional mechanism of EST on colorectal tumor cells has rarely been reported.
In this work, we sought to investigate the effect of EST on proliferation and migration ability of colon tumor cells, and further explore its mechanism of function.
Materials and methods
Materials
Human colon cancer cell line Caco-2, HCT-116, SW480 and HT-29 was obtained from Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. 5-FU was purchased from Yangtze River Pharmaceutical Group Co., Ltd. (Jiangsu, China). EST was purchased from DELTA Information Centre for Natural Organic Compouds (China). DMSO was purchased from Sangon Biotech Co., Ltd. (Shanghai, China). DMEM medium, fetal bovine serum (FBS) and TRIzol Kit were purchased from Gibco Thermo Fisher Scientific Inc. 3-(4,5-Dimethyl-2-Thiazolyl)-2,5-Diphenyl Tetrazolium Bromide (MTT) was purchased from Sigma-Aldrich, Inc. (USA). CCK-8 reagent was purchased from Dojindo Molecular Technologies, Inc. RIPA lysis buffer and ECL Kit were purchased from Beyotime Biotechnology, Inc. (China). ELISA Kit for Human Wnt3a, VEGF protein was purchased from Cusabio Technology Llc. (Wuhan, China). All the antibodies used in this work were purchased from Santa Cruz Biotechnology, Inc. (USA).
Cell culture and administration protocol
Human colon cancer cells was cultured in DMEM medium (containing 10% FBS, 100 U/mL penicillin and 100 g/mL streptomycin) at 37C in an incubator with 5% CO and 95% humidified atmosphere. Cells were firstly incubated for 48 h with EST and 5-FU at several concentrations (EST: 0.2, 2, 20, 200, 2000 g/mL; 5-FU: 0.1, 1, 10, 100, 1000 g/mL), then, CCK-8 was applied to evaluate the in vitro cytotoxicity. In subsequent experiments, grouping and administration protocol was set as follows: control group (DMSO at equal volume), EST group (EST at 20 g/mL), 5-FU group (5-FU at 10 g/mL) and EST+5-FU group (EST 20 g/mL + 5-FU 10 g/mL). After dosing, cells were incubated with drugs at 37C in 5% CO atmosphere for another 48 h before the cell viability assay.
Cell viability assay by CCK-8
Cells in exponential phase were digested with 0.25% trypsin, and the density of cell suspension was adjusted to 5 10/mL. Cells were then seeded into 96-well plates with 100 L/well. Subsequently, cells were incubated as the administration protocol described above. After the administration, the culture medium in each well was removed and substituted with 100 L of fresh medium, and 10 L of CCK-8 solution was added to each well of the plate. After 3-hour incubation in the incubator, the absorbance at 450 nm (OD value) was measured using a microplate reader. The cell inhibitory rate was calculated using the following formula:
Inhibitory rate (%) 1 [(OD OD)/(OD OD)] 100%, where OD is the OD value of the untreated group, OD is the OD value of the drug treated group, and OD is the OD value of the control group.
Cell viability assay by MTT
HCT-116 cells were digested and seeded into 96-well plate as the method mentioned above. After the administration, 20 L of MTT (5 mg/mL) was added to each well. Cells were then incubated for another 4 h at 37C. Subsequently, the supernatant was removed, and 150 L of DMSO was added. The plates were shaken in the dark to dissolve the precipitated formazan crystals and the OD values were measured with a microplate reader at a wavelength of 492 nm.
Colony formation assay
Cells were grouped and treated according to the above methods. After the administration, cells were digested with trypsin and replated into 6-well plate at a density of 2 10/well for 2 weeks. The culture medium was then aspirated, and colonies were washed with PBS, fixed with methanol, stained with 0.4% methylene blue solution, and counted.
Western blot analysis
Cells were grouped and treated according to the above methods. After the administration, cells were harvested, washed with PBS and lysed in RIPA lysis buffer. The protein concentration was determined using BCA protein assay kit. Aliquots of the lysates (50 g of protein) were electrophoresed in a 10% Invitrogen Bis-Tris-Gel at 200 V for 40–50 min. The proteins were then transferred to PVDF membranes, which were subsequently blocked in 5% non-fat milk buffer for 2 h at room temperature. After that, the membrane were incubated with specific primary antibodies overnight at 4C, followed by wash with 1 TBST and incubation with secondary antibodies for 2 h at room temperature. Subsequently, the membrane was washed with 1 TBST for 3 times, and protein bands were detected using an ECL kit according to the manual of the kit, followed by exposure of the membrane.
ELISA
Cells were grouped and treated according to the above methods. After the administration, cells were counted and the culture medium supernatant was collected. Wnt3a and VEGF levels in the culture medium supernatant were measured using ELISA according to the manual of the ELISA kit.
Scratch assay
Cell suspension was prepared with HCT-116 cells of exponential phase, then cells were seeded into 6-well plate at at a density of 8 10/well and cultured to the formation of a confluent monolayer. Thereafter, a straight scratch was created on the monolayer by a sterile pipette tip. Cells were then treated as the methods mentioned above, along with 1 g/mL of mitomycin to inhibit cell division. After 24 h incubation, the closure of the scratch was observed every 2 h until the scratch area was filled with cells. The time required for cells to cross the scratch area was recorded.
In vivo test
Animal grouping
Forty mice were randomly divided into 4 groups with 10 mice in each group. The difference of the body weight between each group was not significant. Animal grouping and administration protocol was set as follows: control group (DMSO at equal volume), EST group (EST at 50 mg/kg/day via intragastric administration), 5-FU group (5-FU at 10 mg/kg/day via intraperitoneal injection) and EST+5-FU group (EST at 50 mg/kg/day + 5-FU at 10 mg/kg/day via intraperitoneal injection). All drug pretreatment in each group lasted for 2 weeks.
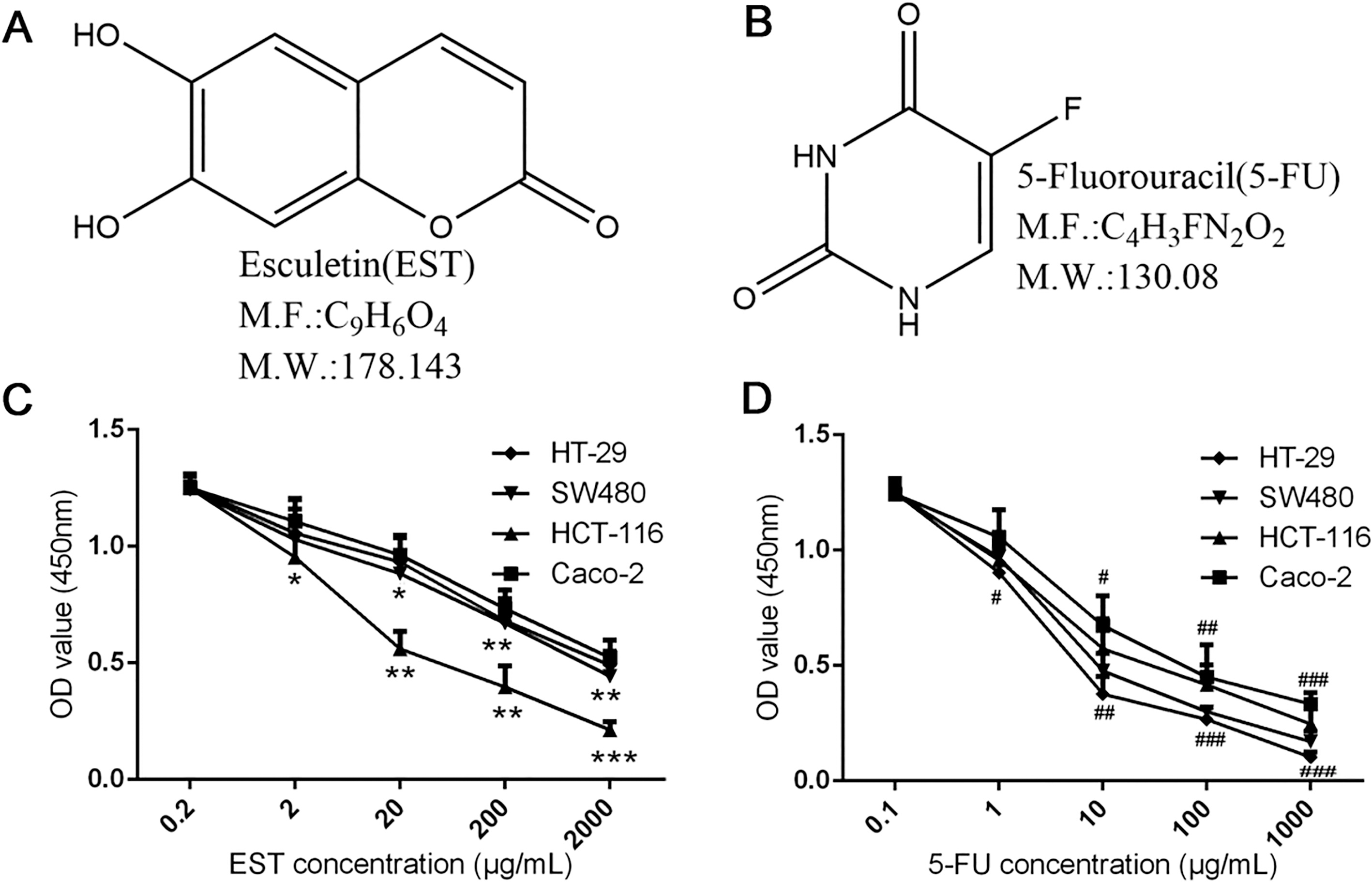
Inhibitory effects of EST and 5-FU with different concentrations on proliferation of 4 human colon cancer cell lines. A and B. Chemical structure of EST and 5-FU; C and D. Cell viability assay by CCK-8 (EST: 0.2, 2, 20, 200, 2000 g/mL; 5-FU: 0.1, 1, 10, 100, 1000 g/mL). 0.05, 0.01, 0.001 vs 0.2 g/ml EST group; 0.05, 0.01, 0.001vs 0.1 g/ml 5-FU group.
HCT-116 cells cryopreserved in liquid nitrogen were resuscitated, expanded, and injected to the right flank of 3 nude mice and 2 weeks later, mice were sacrificed and the tumor tissue was isolated and digested to sterile single cell suspension at a density of 1 10/ml. Trypan blue staining was applied to detect if the cell viability is more than 95%. Cells suspensions were then subcutaneously injected to the armpit of mice (2 ml per mouse). After tumor inoculation, animals were treated with drugs as described above for 2 weeks.
Indexes detection
Survival curve: After the tumor inoculation, mice were normally fed until it spontaneously died. Data were recorded and the survival curves were graphed.
Tumor volume: After the period of treatment, the freshly isolated tumor tissues were kept and the tumor volume was measured on the day of specimen collection (The tumor volume of the mice died during the treatment was not included in the statistics.).
Expression of VEGF in tumor: The streptavidin-peroxidase immunohistochemis try (SP IHC) method was conducted to detect the tumoral expression of VEGF according to the manual of the SP IHC kit. The positive substance showed fine grainy, with the brown yellow region was considered as positive cells.
Expression of Ki67, PCNA and -catenin in tumor, as well as the downstream markers c-Myc, Cyclin D1, MMP2 and MMP7: Western blot analysis was used to detect the expression of the biomarkers antibodies used in this work were purchased from Santa Cruz Biotechnology, Inc. (USA).
Statistical analysis
Statistical analysis was performed by SPSS 18.0. All the data were presented as the mean value standard deviation (SD). After the homogeneity test of variance, -test was performed when comparing the difference of mean value between groups with equal variance, while -test was used for groups with uneven variance. Log-rank test is carried out for comparing the survival curves. A value 0.05 was considered statistically significant.
Results
Inhibition effect of EST and 5-FU on proliferation of 4 human colon cancer celllines
As shown in Fig. 1C and D, after treatment with EST or 5-FU alone, both viability of 4 human colon cancer cell lines were inhibited by different degrees with dosage-dependent property. Notably, when the EST and 5-FU concentration reached 20 g/mL and 10 g/mL respectively, the inhibition of HCT-116 cell proliferation by these two drugs could be obviously observed as compared with control group ( 0.01).
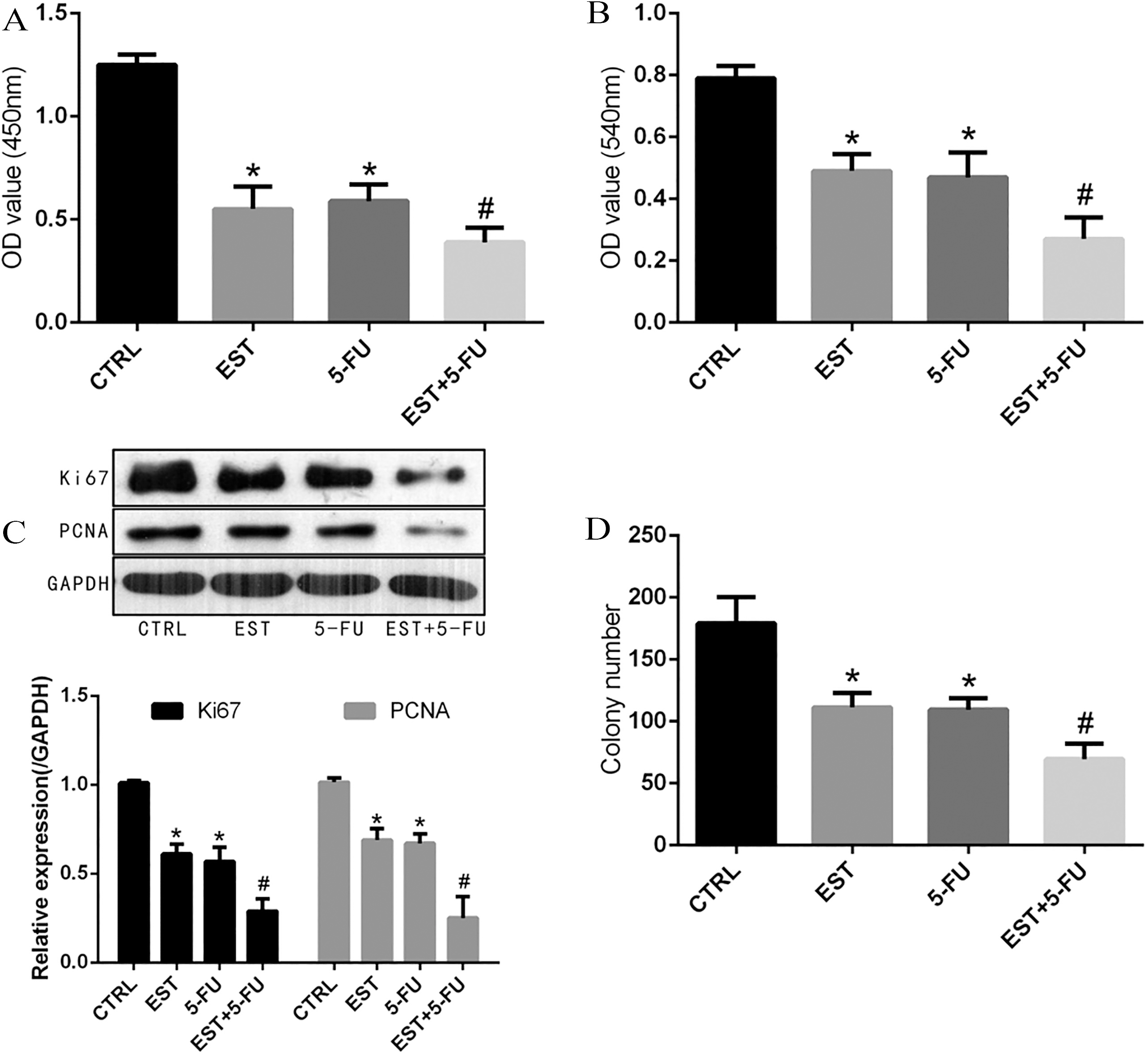
Inhibitory effect of EST on the proliferation of HCE-116 cells. A and B. Cell proliferation ability was measured by CCK-8 and MTT assay; C. Colony formation assay; D. Expression of proliferation-related protein Ki67 and PCNA. 0.05 vs CTRL group, 0.05 vs 5-FU group.
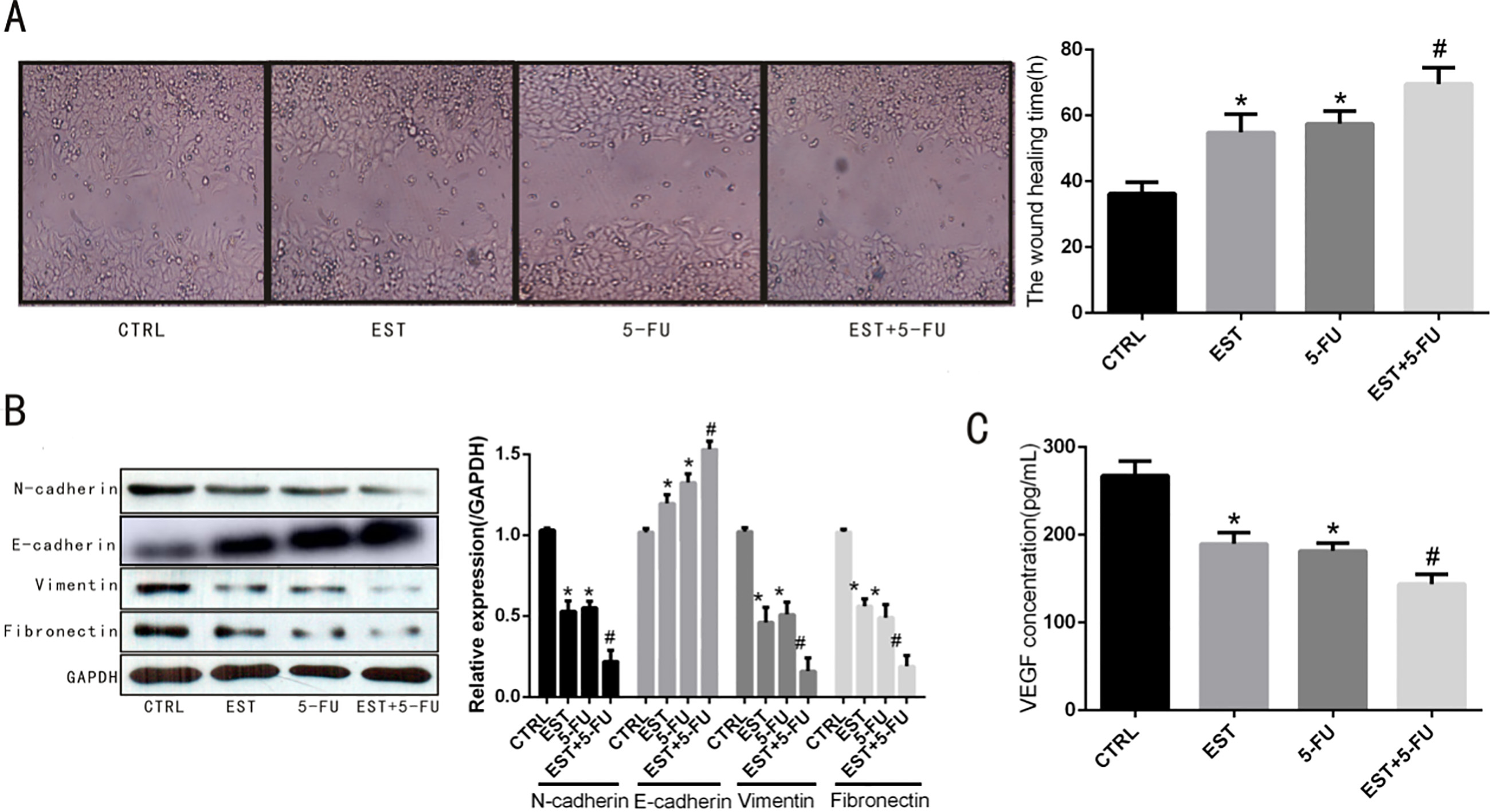
Effect of EST on HCT-116 cells migration. A. Time required for cells to cross the scratch area in the scratch assay. B. Expression of migration-related proteins N-cadherin, E-cadherin, vimentin and fibronectin. C. Content of VEGF in cell culture medium supernatant. 0.05 vs CTRL group, 0.05 vs 5-FU group.
EST enhanced the inhibition effect of 5-FU in HCT-116 cells proliferation andcolony formation
As displayed in Fig. 2A and B, results in CCK-8 and MTT assays confirmed that both EST and 5-FU could inhibit the proliferation activity of HCT-116 cells, and the combined treatment of EST and 5-FU showed significantly stronger inhibition efficiency when compared with mono-drug treatment ( 0.05). Meanwhile, colony formation assay showed that the average colony number of control group, EST group, 5-FU group and EST+5-FU group were 179 21.26, 111.33 10.57, 109.49 9.36 and 69.49 4.68 per well, respectively. The average colony number of EST group, 5-FU group and EST+5-FU group was significantly lower in contrast to control group ( 0.05). In addition, Fig. 2C indicated that the average colony number of EST+5-FU group was significantly lower compared with EST group and 5-FU group, respectively ( 0.05). Western blot analysis showed that the expression of proliferation-related protein Ki67 and PCNA was significantly decreased in EST group, 5-FU group and EST+5-FU group compared with control group ( 0.05). In addition, the expression of proliferation-related protein Ki67 and PCNA of EST+5-FU group was significantly lower compared with EST group and 5-FU group,respectively ( 0.05), as displayed in Fig. 2D.
EST enhanced the inhibition effect of 5-FU in HCT-116 cells migration and epithelial- mesenchymal transition
Scratch assay Fig. 3A showed that the time required for HCT-116 cells to cross the scratch area of control group, EST group, 5-FU group and EST+5-FU group were 36.25 3.46, 54.82 5.6, 57.49 3.85 and 69.59 4.89, respectively. The time required for cells to cross the scratch area of EST group, 5-FU group and EST+5-FU group was significantly longer than that of the control group ( 0.05). Besides, Fig. 3A indicated that the time required for cells to cross the scratch area of EST+5-FU group was significantly longer when compared with EST group and 5-FU group, respectively ( 0.05). Western blot analysis showed that the expression of EMT-related proteins N-cadherin, vimentin and fibronectin were significantly decreased across EST+5-FU treatment compared with EST treatment or 5-FU treatment alone, meanwhile, E-cadherin protein expression was significantly increased across EST+5-FU treatment compared with EST treatment or 5-FU treatment alone, as demonstrated in Fig. 3B. Moreover, the results of ELISA showed that content of VEGF in cell culture medium supernatant was significantly decreased after treating with EST+5-FU compared with EST treatment or 5-FU treatment alone, as demonstrated in Fig. 3C.
EST enhanced the inhibition effect of 5-FU in Wnt/-catenin signaling pathway in HCT-116 cell
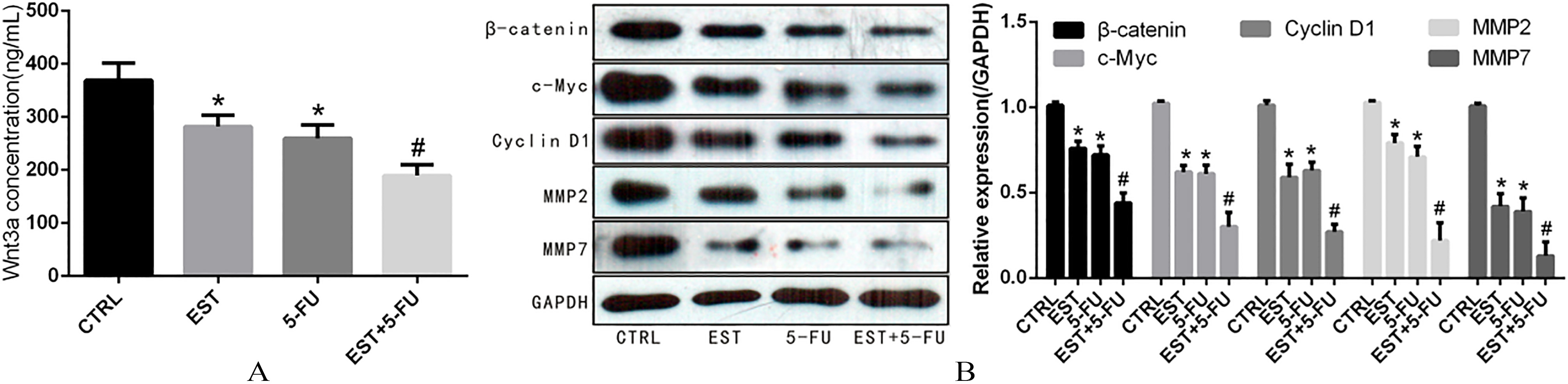
The results of ELISA showed that content of Wnt3a in cell culture medium supernatant was significantly decreased after treatment with EST and 5-FU. As shown in Fig. 4A, the concentration of Wnt3a of control group, EST group, 5-FU group and EST+5-FU group were 368.90 32.75, 281.93 21.51, 259.68 25.11 and 189.54 20.17 pg/ml, respectively. Western blot analysis showed that the expression of -catenin, c-Myc, Cyclin D1, MMP2 and MMP7 was significantly decreased after EST and 5-FU intervention, as demonstrated in Fig. 4B.
Effect of EST on Wnt signaling pathway in HCT-116 cell. A. Content of Wnt3a in cell culture medium supernatant; B. Expression of -catenin and its downstream target molecules c-Myc, Cyclin D1, MMP2 and MMP7. 0.05 vs CTRL group, 0.05 vs 5-FU group.
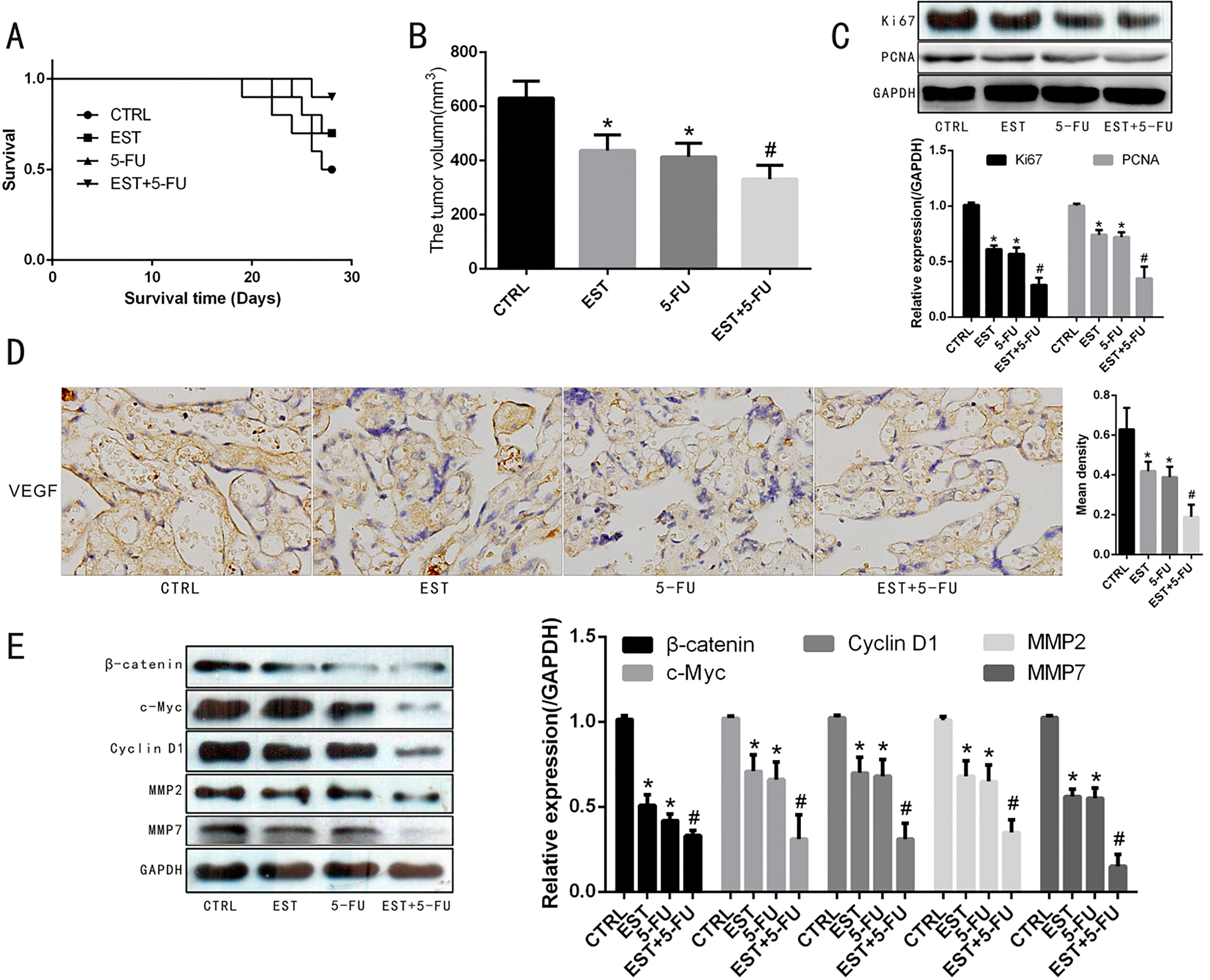
Effect of EST on HCT-116 tumor bearing mice. A. Survival analysis after tumor inoculation. B. Tumor volume of HCT-116 tumor bearing mice. C. Western blot assay of expression of Ki67 and PCNA in tumor. D. Immunocytochemical staining of level of VEGF in tumor. E. Western blot assay of expression of Wnt signaling pathway related proteins in tumor. Note: 0.05 vs CTRL group, 0.05 vs 5-FU group.
EST enhanced the inhibition effect of 5-FU in tumor growth of subcutaneous HCT-116 xenograft in mice
As shown in the survival curves analysis in Fig. 5A, although the difference between EST group and control group was not statistically significant, the survival of mice in EST+5-FU group was significantly improved than that of the control group ( 0.05). Tumor volume of the control group, EST group, 5-FU group and EST+5-FU group were 631.0 62.9, 437.1 57.9, 413.6 50.7 and 331.4 51.2 respectively. The tumor volume of the mice in the 3 drug-intervened groups were significantly smaller than that of the control group ( 0.05). In addition, Fig. 5B showed that the tumor volume of EST+5-FU group was significantly smaller when compared with EST group or 5-FU group, respectively ( 0.05). The immunocytochemical staining (Fig. 5D) showed that the proportion of VEGF-positive cells was obviously decreased across EST and 5-FU intervention. In addition, Western blot analysis showed that the expression of proliferation-related proteins of Ki67, PCNA and downstream molecules of Wnt signaling pathway (-catenin, c-Myc, Cyclin D1, MMP2 and MMP7) was significantly down-regulated after EST and 5-FU intervention, as demonstrated in Fig. 5C and E.
Discussion
Some studies reveal that the active component EST of Cortex fraxini in the classical prescription could effectively inhibit the proliferation of tumor cells [31]. Here, we probed into the further mechanism of EST on the inhibition of proliferation and migration of colon cancer cells, as well as the anti-tumor efficiency of EST combined with 5-FU.
After treated with EST of different concentration, the proliferation ability of colon cancer cells HCT-116 was obviously decreased with dosage-dependent property. The colony formation assay also showed that the EST treatment could significantly inhibit the formation of HCT-116 cell colony. Moreover, the scratch assay demonstrated that after EST intervention, the time required for cells to cross the scratch area was significantly extended compared with the control group, indicating that EST treatment could inhibit the migration ability of colon cancer cells. Notably, the results in EST+5-FU group showed that EST could enhance the inhibitory effect of 5-FU on the proliferation and migration of HCT-116 cell. Further studies on RT-PCR and western blot displayed that the expression of proliferation-related proteins Ki67 and PCNA, as well as migration-related molecules N-cadherin, E-cadherin, Vimentin and Fibronectin was significantly decreased after EST treatment. The results of ELISA confirmed the decrease of expression of VEGF in cell culture medium supernatant when cells were incubated with EST.
It is widely known that the Wnt/-catenin signaling pathway, which is involved in a variety of biological processes including proliferation, differentiation, apoptosis and metastasis, plays an important role in the development of tumor [32]. The intracellular aggregation of -catenin is translocated into the nucleus to achieve regulation of downstream target genes (such as C-Myc, Cyclin D1, MMP2, MMP7 [33, 34, 35]. A variety of studies have found that the aberrant activation of Wnt signaling is one of the important causes of colon cancer, and -catenin and its downstream target genes play a key role in it [36]. As a fact of that, Wnt/-catenin has become an important target for the treatment of CRC.
Studies have shown that EST has an inhibition effect on Wnt/-catenin signaling pathways. In our research, we validated the role of EST in inhibiting the migration of HCT-116 cell proliferation and whether its action mechanism is related to the Wnt/-catenin signaling pathway. First, the results of ELISA showed that the Wnt3a concentration in the HCT-116 cell culture medium supernatant in the EST-treated group was significantly lower than that in the control group. Then, WB results verified that after EST intervention, the expression of -catenin and its downstream cell cycle-related target molecules (c-Myc and Cyclin D1) and tumor migration-associated target molecules (MMP2 and MMP7) were significantly down-regulated in HCT-116 cells. Both results demonstrated the inhibition effect of EST on proliferation and migration of HCT-116 cells was related to the regulation of EST to Wnt/-catenin signaling pathway as well as its downstream cell cycle-related proteins (c-Myc and Cyclin D1) and proteolytic enzymes (MMP2 and MMP7). Finally, the results of subcutaneous HCT-116 xenografts in mice revealed that EST could effectively inhibit the expression of proliferation and migration-related molecules in HCT-116 tumor tissue, inhibit tumor growth, and prolong the survival of tumor-bearing mice. In the survival curves analysis, the difference between EST group and control group was not statistically significant. However, it is worth noting that the statistic difference between the combined treatment group and the control group was significant. Hence, the no significant difference between the EST or 5-FU group alone and the control group might be related to the relatively small sample size and short drug treatment time. Through cell proliferation, migration, and expression of interstitial transformation markers, we hypothesized that the mechanism of action of EST may be to reduce tumor growth by reducing VEGF expression and affecting Wnt/-catenin signaling pathways.
In conclusion, our research verified that EST can inhibit the expression of tumor cell cycle-related proteins and proteolytic enzymes, inhibit the proliferation and migration of colon tumors by inhibiting the activity of Wnt/-catenin signaling pathway, and can further significantly enhance the anti-tumor effect of 5-FU to HCT-116 both in vitro and in vivo. To sum up, this study provide further experimental evidence for EST as a potential clinical colon cancer drug, along with new therapeutic directions for clinical treatment of CRC, especially for patients with middle and late stage colon cancer and chemotherapeutic resistance.
Footnotes
Acknowledgments
This work was financially supported by the Natural Science Foundation of Shandong Province (ZR2010 HL018).
Conflict of interest
The authors have declared that no conflict of interest.
Supplement Pictures
Note: Representative xenograft tumors excised from the mice of each group.
References
1.
TorreL.A. et al., Global cancer statistics, CA: A Cancer Journal for Clinicians65(2) (2015), 87–108.
2.
WangY.N. et al., Clinical characteristics of ulcerative colitis related colorectal cancer in Chinese patients, J Dig Dis18(12) (2017), 684–690.
3.
LiK. et al., Spatio-temporal analysis of the incidence of colorectal cancer in Guangzhou, 2010–2014, Chin J Cancer36 (2017), 60.
4.
LiM. et al., Cancer mortality trends in an industrial district of Shanghai, China, from 1974 to 2014, and projections to 2029, Oncotarget8(54) (2017), 92470–92482.
5.
JemalA. et al., Cancer statistics, CA Cancer J Clin55 (2005), 259.
6.
ArberN. and LevinB., Chemoprevention of colorectal cancer: ready for routine use? Recent Results Cancer Res166 (2005), 213–230.
7.
ArmandJ.P. et al., CPT-11 (irinotecan) in the treatment of colorectal cancer, Eur J Cancer31 (1995), 1283–1287.
8.
CvitkovicE. and BekraddaM., Oxaliplatin: a new therapeutic option in colorectal cancer, Semin Oncol26 (1999), 647–662.
9.
GuichardS. et al., Sequence-dependent activity of the irinotecan-5FU combination in human colon-cancer model HT29 in vitro and in vivo, Int J Cancer73 (1997), 729–734.
10.
YuH.G. et al., The effects of acetylsalicylic acid on proliferation, apoptosis, and invasion of cyclooxygenase-2 negative colon cancer cells, Eur J Clin Invest32 (2002), 838–846.
11.
GavertN. and Ben-ZeevA., Epithelial-mesenchymal transition and the invasive potential of tumors, Trends Mol Med14 (2008), 199–209.
12.
BruzzeseF. et al., HDAC inhibitor vorinostat enhances the antitumor effect of gefitinib in squamous cell carcinoma of head and neck by modulating ErbB receptor expression and reverting EMT, J Cell Physiol226 (2011), 2378–2390.
13.
YilmazM. and ChristoforiG., EMT, the cytoskeleton, and cancer cell invasion, Cancer Metastasis Rev, 28 (2009), 15–33.
14.
ZhouB.B. et al., Tumour initiating cells: challenges and opportunities for anticancer drug discovery, Nat. Rev Drug Discov8 (2009), 806–823.
15.
KalluriR. and WeinbergR.A., The basics of epithelial-mesenchymal transition, J. Clin. Invest119 (2009), 1420–1428.
16.
SegditsasS. and TomlinsonI., Colorectal cancer and genetic alterations in the Wnt pathway, Oncogene25 (2006), 7531–7537.
17.
MorinP.J. et al., Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC, Science275 (1997), 1787–1790.
18.
FearnheadN.S.WildingJ.L. and BodmerW.F., Genetics of colorectal cancer: hereditary aspects and overview of colorectal tumorigenesis, British Medical Bulletin64 (2002), 27–43.
19.
MoravecM., Colorectal cancer and canonical Wnt signalling pathway, Casopis Lekaru Ceskych151 (2012), 335–342.
20.
BuchertM. et al., Genetic dissection of differential signaling threshold requirements for the Wnt/beta-catenin pathway in vivo, PLoS Genetics6 (2010), e1000816.
21.
KlausA. and BirchmeierW., Wnt signalling and its impact on development and cancer, Nature reviews Cancer8 (2008), 387–398.
22.
HeT.C. et al., Identification of c-MYC as a target of the APC pathway, Science281 (1998), 1509–1512.
23.
ShtutmanM. et al., The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway, Proceedings of the National Academy of Sciences of the United States of America96 (1999), 5522–5527.
24.
TetsuO. and McCormickF., Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells, Nature398 (1999), 422–426.
25.
KaurM. et al., Grape seed extract induces cell cycle arrest and apoptosis in human colon carcinoma cells, Nutr Cancer60(Suppl 1) (2008), 2–11.
26.
ZhuL. et al., Esculetin attenuates lipopolysaccharide (LPS)-induced neuroin-flammatory processes and depressive-like behavior in mice, Physiol Behav163 (2016), 184–192.
27.
WangJ. et al., Esculetin, a coumarin derivative, exerts in vitro and in vivo antiproliferative activity against hepatocellular carcinoma by initiating a mitochondrial-dependent apoptosis pathway, Braz J Med Biol Res48(3) (2015), 245–253.
28.
ZhuX.GuJ. and QianH., Esculetin Attenuates the Growth of Lung Cancer by DownregulatingWnt Targeted Genes and Suppressing NF-B, , 2017, S0300–2896.
29.
LeeR.H. et al., Esculetin exerts anti-proliferative effects against non-small-cell lung carcinoma by suppressing specificity protein 1 in vitro, Gen Physiol Biophys36(1) (2017), 31–39.
30.
RubioV. et al., Esculetin modulates cytotoxicity induced by oxidants in NB4 human leukemia cells, Exp Toxicol Pathol69(8) (2017), 700–712.
31.
ParkS.S. et al., Esculetin inhibits cell proliferation through the Ras/ERK1/2 pathway in human colon cancer cells, Oncol Rep25(1) (2011), 223–230.
32.
KretzschmarK. and CleversH., Wnt/β-catenin signaling in adult mammalian epithelial stem cells, DevBiol428(2) (2017), 273–282.
33.
ZhangZ. et al., HMGB3 promotes growth and migration in colorectal cancer by regulating WNT/β-catenin pathway, PLoS One12(7) (2017), e0179741.
34.
ParkS.B. et al., Anticancer activity of calyx of Diospyros kaki Thunb. through downregulation of cyclin D1 via inducing proteasomal degradation and transcriptional inhibition in human colorectal cancer cells, BMC Complement Altern Med17(1) (2017), 445.
35.
YuanX. et al., USP39 promotes colorectal cancer growth and metastasis through the Wnt/β-catenin pathway, Oncol Rep37(4) (2017), 2398–2404.
36.
ShangS.HuaF. and HuZ.W., The regulation of β-catenin activity and function in cancer: therapeutic opportunities, Oncotarget8(20) (2017), 33972–33989.