
Abstract
Looking back over 50 years of studying social insects, this article identifies eight outstanding problems in social insect biology: 1) Can life cycles of individual organisms serve as a reliable model for the colony cycle?; 2) Does the production of workers and sexuals in social hymenopterans fit the bang-bang model?; 3) Do females in social hymenopterans have a smaller individual behavioural repertory than those of solitary hymenopterans?; 4) Is memory in termite workers and soldiers limited to the sensory level?; 5) Do some social wasps and/or bees demonstrate coordinated colony defense?; 6) Is there an induced defense in termite colonies at the level of caste proportions?; 7) Are social wasps passing through a period of global population decrease?; and 8) Is there a relationship between the level of primary production in the habitat and the proportion of social species among the insects?
Introducción
Los insectos sociales se distinguen de la masa de especies solitarias por su hábito de vivir en grupos estructurados, o colonias. Aunque abarcan no más del 2 % de las especies de insectos, han llamado la atención científica y filosófica desde la antigüedad. Casi todos pertenecen a cuatro grupos: a) comejenes, o termitas (Blattaria, suborden Isoptera) con unas 3000 especies (Korb, 2021), b) avispas sociales (Hymenoptera: Vespidae) con unas 1100 especies (Jeanne, 2021), c) abejas sociales con aproximadamente 1000 especies en varios linajes de Hymenoptera (Engel et al., 2021), y d) las hormigas (Hymenoptera: Formicidae) con unas 12,000 especies descritas (AntWeb, 2024).
Se puede fechar el inicio de la sociobiología de insectos desde hace casi un siglo. Antes de entonces había obras sobre la biología de los comejenes (Escherich, 1909; Hegh, 1922), las avispas sociales (Réaumur, 1742), la abeja de miel (Bevan, 1838; Butler, 1609; Huber, 1814; Purchas, 1657; Réaumur, 1740; Thorley, 1744), otras abejas sociales (Buttel-Reepen, 1903, 1915; Sladen, 1912) y las hormigas (Emery, 1915; Escherich, 1906; Huber, 1810; Latreille, 1802; Wasmann, 1915; Wheeler, 1910). Sin embargo, Wheeler (1923, 1928) fue el primero en tratar la vida de estos insectos generalmente. Desde entonces han aparecido varios volúmenes de similar amplitud (Maidl, 1934; Wilson, 1971; Hermann, 1982; Starr, 2021a).
Durante el último año de mi bachillerato, conseguí mi propia copia de la ahora clásica obra de Wilson (1971), The Insect Societies. Entonces acababa de llegar a considerar los insectos sociales como el aspecto más interesante del universo. Durante los últimos 50 años he hecho un gran número de preguntas e hipótesis acerca de estos bichos. En algunos casos, he logrado resolver una u otra cuestión (no necesariamente por mis propios esfuerzos). En otros llegué a considerarla como no tan importante. Y hay otras que siguen no resueltas y contribuyen a mis episodios de insomnio. Mi propósito aquí es describir ocho de estas preguntas que me interesan con algunas sugerencias hacia sus resoluciones.
Desarrollo temático
1. ¿Puede ser el ciclo de vida del individuo un estimador confiable del ciclo de una colonia?
Las colonias de insectos sociales tienen muchas características en común con los animales individuales. Hace más de un siglo se ha notado que las colonias, igual que los organismos, pasan por un ciclo de fundación, crecimiento, madurez, senescencia y muerte (Starr, 2021b). Por este motivo, se pueden considerar como organismos al segundo nivel, o sea super-organismos (Hölldobler & Wilson, 2008; Starr, 2021c; Wheeler, 1911).
Ya hace mucho tenemos gran cantidad de literatura alrededor de los ciclos de vida de plantas y animales (Roff, 2002; Stearns, 1992, 2000). A diferencia de esto, la teoría del ciclo colonial aún está mucho menos madura y en gran parte consiste en poco más de una serie de analogías con la del ciclo de vida. Al leer obras sobre el ciclo de vida se podría sustituir toda mención de ciclo de vida con ciclo colonial sin que se haga absurdo. Claro que lo no absurdo y lo cierto no son necesariamente iguales. ¿Siguen los ciclos coloniales patrones semejantes a los ciclos de vida? No sabemos. Apenas existe literatura sobre los ciclos coloniales que se refiere a ellos como complejos adaptativos. En vez de brindar conclusiones, los conceptos actuales sobre ciclos de vida brindan un punto de partida en cuanto a los ciclos coloniales.
No cabe duda de que estas analogías son una rica fuente de conceptos e hipótesis (Bourke & Franks, 1995), pero existe el riesgo de aceptar estos como algo ya demostrado. En el próximo período hemos de construir una teoría general del ciclo colonial que pasará de ser una simple analogía con la más madura teoría del ciclo de vida.
Pueden existir diferencias significativas entre los dos tipos de ciclos, y esto surgiría del hecho de que los miembros de una colonia, a diferencia de las celdas de un mismo cuerpo, no son un clono. Es decir, los miembros de una misma colonia no tienen exactamente los mismos intereses en cuanto a la producción de nuevos miembros.
Se reconoce una distinción entre ciclos determinados e indeterminados, analógica a la distinción entre especies cuyos individuos reproducen toda su cría a una vez antes de morir (semelparidad) y los que reproducen repentinamente (iteroparidad) (Starr, 2021b). Entre las avispas sociales del neotrópico, por ejemplo, es probable que todas las especies de fundación independiente (e.g., Mischocyttarus y Polistes) tengan ciclos coloniales determinados (Fig. 1a), mientras que la mayoría de enjambradoras (e.g., Polybia y géneros emparentados) tengan ciclos indeterminados (Fig. 1c) (Starr, 2006). Pero no sabemos por falta de datos básicos si esta hipótesis sea correcta.
Tschinkel (1991) introdujo la sociometría para la descripción analítica de las colonias de insectos y enfatizó que era un campo de investigaciones descuidado, donde algunas cuestiones importantes apenas pueden progresar por carencia de datos básicos. Desafortunadamente, la colección de datos básicos hoy en día no alcanza suficiente desarrollo.
Como primera propuesta hacia una tal concepción en la sociobiología, Tschinkel (1991) presentó una lista de 38 aspectos mensurables de la colonia que consideraba como significativos. Reconociendo que la preparación de tal formulario, así como la colección de datos a cumplirlo, tiene que ser una obra colectiva, pidió una discusión de este asunto en la comunidad de sociobiólogos. Hasta ahora, la obra de Tschinkel (1991) queda poco comentada. Actualmente, alguien que necesite un conjunto de datos comparativos sobre algún aspecto de los insectos sociales normalmente los busca en la literatura antes de hacer una llamada para datos inéditos. Por ejemplo, tengo en frente de mí un mensaje mandado a una red electrónica. El colega está interesado en comparar la longevidad de reinas de diversos grupos; ya ha sacado de la literatura datos sobre 62 especies de hormigas y comejenes, y pide otros.
Este método de acumular datos comparativos se ha facilitado mucho por el internet, sin embargo, el acceso a dichos datos es difícil y lento debido a la falta de una base de datos pública, centralizada y recuperable como la que sigue el modelo de la Human Relations Area Files, base de datos antropológicos mantenida en la Universidad Yale (https://hraf.yale.edu/). Dada la existencia de la Unión Internacional para el Estudio de Insectos Sociales (UIEIS), es preciso pensar en formular, como proyecto colectivo una serie de parámetros sociométricos consolidados en un base de datos. Por tanto, se propone que la UIEIS considere la posibilidad de establecer y mantener una base de datos permanente sobre los insectos sociales.
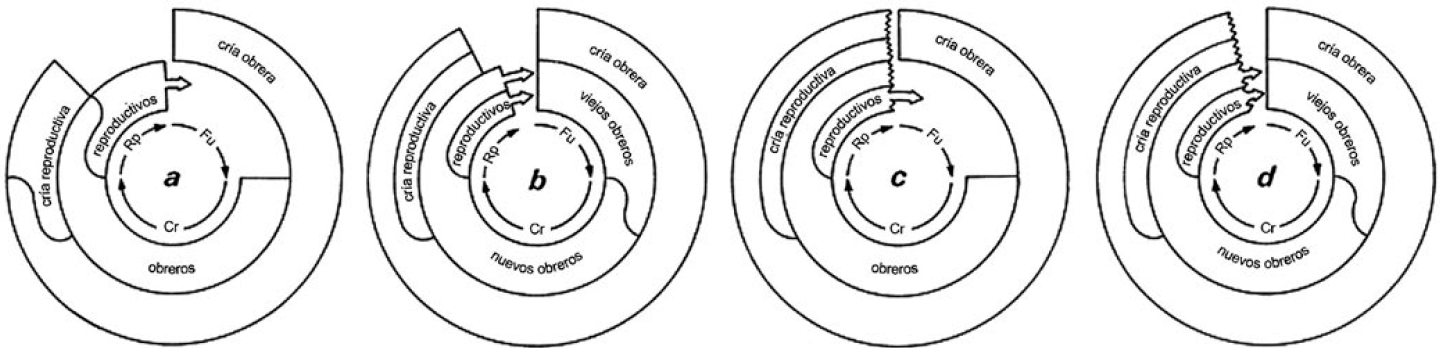
Los cuatro tipos básicos del ciclo colonial en insectos sociales. a. Fundación independente con una sola fase de producción de reproductivos. b. Fundación por enjambre con una sola fase de producción de reproductivos. c. Fundación independente con repetidas fases de producción de reproductivos. d. Fundación por enjambre con repetidas fases de producción de reproductivos. Fu. Fase de fundación, Cr. Fase de crecimiento. Rp. Fase de producción de nuevas reinas y machos. Modificada de Starr (2006).
2. ¿El patrón de producción de obreras y reproductores en insectos sociales se ajusta al modelo bang-bang?
Las colonias de himenópteros sociales producen individuos reproductores (reinas y machos) y no reproductores (obreras). ¿Cuál es el patrón temporal de producción de estos dos tipos de cría? A un extremo, la colonia podría producir ambos durante toda la temporada (Fig. 2a), pero esto no parece resultar en ninguna especie. Normalmente, una colonia empieza con la producción de obreras (fases de fundación y de crecimiento) y sólo después se ponen los primeros huevos destinados a desarrollarse como reinas y/o machos (fase reproductiva).
Al otro extremo, podrían separarse rigurosamente las dos fases (Fig. 2d). Por razones teóricas, se ha propuesto que la colonia optimizará su producción al seguir este patrón llamado bang-bang (Oster & Wilson, 1978). Sin embargo, los escasos datos ya disponibles sugieren que el ciclo no conforma estrictamente al modelo bang-bang, eso es, que hay una cierta superposición de las dos etapas de producción (Greene, 1984) (Fig. 2b-c).
Al momento de la escritura de este ensayo, no se ha sugerido ninguna base para este estado intermedio en contra al modelo bang-bang, ni menos una base para pronosticar diferencias entre especies. Un primer paso hacia este análisis sería la formulación de un índice que cuantifique la variación entre producción regular (Fig. 2a) y bang-bang (Fig. 2d). Este índice proporcionaría un medio para comparar las diferentes especies y poblaciones, lo que facilitaría hipótesis causales.
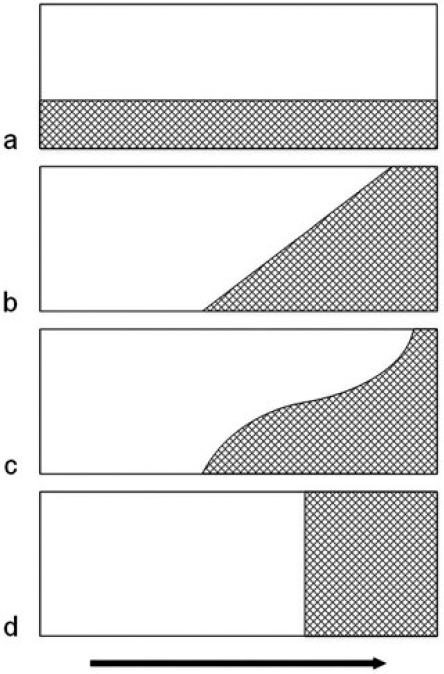
Patrones idealizados de la producción de huevos de cría de obreras (blanco) y de reproductivos (rayado) durante toda la temporada (flecha). a. Regular durante todo el ciclo. b-c. Patrones intermedios, con superposición parcial de las dos fases. d. Los dos tipos de cría están rigurosamente separados (bang-bang).
3. ¿Tienen las hembras de los himenópteros sociales un repertorio comportamental inferior a los himenópteros solitarios?
Según la hipótesis del cerebro social, las exigencias de la vida social en algunos linajes de vertebrados han conducido a cerebros aumentados. Se ha sugerido que lo mismo ocurre en los himenópteros, eso es que las especies sociales tengan cerebros más grandes en relación con el tamaño de aquellos que viven en solitario (Farris & Schulmeister, 2010). Con respecto a lo anterior, este ensayo sugiere exactamente lo contrario. Una hembra de una avispa o abeja solitaria tiene que hacer todo. Busca un sitio a nidificar, construye y/o excava el nido, pone huevos, forrajea, alimenta las larvas, y defiende su cría de los enemigos. Al contrario, y aunque las hembras (reinas y obreras) de himenópteros sociales también emprenden todas estas tareas, lo hacen de una manera colectiva.
En el caso de las especies altamente sociales, en particular, ningún individuo hace todo, ni una vez en su vida si no está especializado. Estudios morfológicos del cerebro en varios insectos tienden a esta hipótesis. En los himenópteros sociales investigados hasta ahora, la parte del cerebro involucrada con memoria y aprendizaje resulta menos desarrollada que en los individuos de conducta más especializada (O'Donnell, 2021). Esto parece implicar que las hembras de especies sociales serán de una manera no tan inteligentes que las solitarias; en el sentido de tener un repertorio comportamental más limitado. Esta hipótesis tiene su lógica, pero apenas hemos empezado a saber si es justa, o si tal vez lo contrario refleja mayor la realidad. Esta cuestión puede resolverse directamente al formular etogramas de especies sociales y solitarias emparentadas. No obstante, una complicación aparece cuando se considera que una gran parte de la actividad ocurre fuera del nido. También se puede preguntar si las cucarachas solitarias tienen repertorios comportamentales más ricos que los de las cucarachas eusociales, o sea comejenes. Muchas cucarachas solitarias, así como comejenes inferiores se dejan criar en el laboratorio bajo condiciones casi naturales.
4. ¿Tienen los obreros y soldados de comejenes una facultad de memoria que no sobrepase al nivel sensorial?
Los animales pequeños necesariamente tienen cerebros más pequeños y sencillos (Eberhard & Wcislo, 2012). Sin embargo, algunos insectos manifiestan capacidades de memoria impresionantes. Por ejemplo, una avispa solitaria del género Ammophila mantiene a la vez varios nidos activos, acordándose no solo de la ubicación sino también del estado de cada uno (O'Neill, 2001). Algunos insectos sociales se destacan igualmente por sus poderes de memoria. Las abejas forrajeras de la especie sin aguijón Melipona subnitida Ducke son capaces de recordar, durante un periodo de hasta 24 horas, el momento del día en que una parcela de flores ofrece la mejor recolección de recursos (Guimarães-Silva et al., 2021). Una abeja de miel, al encontrar una buena fuente de néctar y/o polen, no solo puede acordarse durante días del lugar de esta sino también los colores de las flores y hasta la hora aproximada de mayor productividad (Lindauer, 1971). Igualmente puede comunicar a sus compañeras su ubicación por medio de una danza, tomando en cuenta el movimiento del sol (Couvillon, 2021). Igualmente, las forrajeras de algunas hormigas muestran fuertes capacidades de orientación espacial, incluso en al menos una especie con el uso de patrones del dosel del bosque (Hölldobler, 1980). Además, una obrera al alejarse de su colonia durante algunas horas no olvida el olor de esta (Hölldobler & Wilson, 2008).
Estos son ejemplos de memoria a largo plazo, la que actúa durante al menos algunos minutos. Se distingue de la memoria a corto plazo que actúa por menos de un minuto, y de la memoria a ultra-corto plazo, o memoria sensorial, que se mantiene por menos de un segundo. Esta consiste efectivamente en no olvidar lo que se percibe en el momento inmediato.
La teoría de estigmergía en su forma original (Grassé, 1959) se basa en la construcción del nido por los comejenes superiores. Según esta teoría, los nidos a veces gigantes, complejos y de una forma característica de la especie se forman por un gran número de insectos minúsculos y generalizados siguiendo un algoritmo. Un individuo al encontrar por casualidad cualquier rasgo del nido en desarrollo responde de una manera estereotípica apropiada a ese rasgo. Las diferencias de una especie a otra en las reglas del algoritmo pueden resultar en nidos muy diferentes.
Este modelo no exige ningún papel para la memoria de corto plazo ni menos de largo plazo, nada más de la memoria sensorial. Además, se desconoce alguna actividad de los obreros o soldados, con la posible excepción de las pocas especies que forrajean al aire libre, y que parecen necesitar una facultad de memoria de aunque sea un segundo. Según esto, se podría pensar que los comejenes son los insectos adultos, ya sean solitarios o sociales, que más carecen de memoria.
La anterior hipótesis ya fue sugerida en la prensa popular (Srinivasan, 2018): “Los entomólogos consideran a los comejenes como poco inteligentes, ya que carecen de memoria y la capacidad de aprender.” La memoria de largo plazo, según este punto de vista, es de cierta manera una propiedad de la colonia y no del individuo, incluso su nido. La hipótesis plantea que, en los cerebros de los comejenes no reproductores, el cuerpo pedunculado (o cuerpo champiñón) se encuentra poco desarrollado (O'Donnell, 2021).
5. ¿Existe en algunas avispas y/o abejas sociales una respuesta coordinada en defensa de la colonia?
Al acercarse a una colonia de cualquier especie de Polistes se observa que las avispas se ponen en actitud de alerta (Fig. 3a). Si se acerca aún más, ellas muestran una serie de amenazas distintas y llamativas. Y, si uno continúa provocando, atacan.
Muchas veces sale una multitud de avispas al mismo momento, lo que da la impresión de un ataque coordinado. Esto sugiere tres posibilidades: a) el ataque simultáneo es una ilusión de nuestra parte, b) se trata simplemente de un mismo umbral de respuesta en muchos individuos, o c) existe comunicación visual entre individuos para que vuelen simultáneamente. Esta tercera posibilidad es la más interesante, ya que implica una organización de la defensa más sofisticada, pero no tenemos evidencia segura de que verdaderamente sea así.
La misma cuestión surge en otras avispas y abejas sociales con nido cubierto, si muchos individuos miran por la entrada o están en la superficie (Fig. 3b). Aunque no cuentan con un sistema de amenazas visuales como el de las avispas del género Polistes, estas abejas tienen la capacidad de coordinarse y volar juntas en ataque. Igualmente, la defensa colectiva de abejas de miel contra un ataque por el avispón gigante Vespa mandarinia es bien conocida (McClenaghan et al., 2019), pero no se ha confirmado como una defensa coordinada. La resolución de esta cuestión requerirá llevar a cabo una serie de observaciones estrictamente controladas, con el fin de provocar una respuesta de la colonia de manera regular al aumentar el nivel del estímulo.

Avispas sociales en estado de alerta. Izquierda: Polistes crinitus (Felton), con panal expuesto; todas las adultas pueden ver a otro animal que se acerque. Derecha: Angiopolybia pallens (Lepeletier), con panales dentro de una cubierta; sólo algunos adultos ven afuera del nido.
6. ¿Existe una defensa inducida en los comejenes que afecta la proporción de castas?
Cuando un rasgo de autodefensa se desarrolla a causa de ataques o amenazas previos, se caracteriza como defensa inducida (Harvell, 1990). Algunas hormigas y la mayoría de las especies de comejenes tienen una casta especializada para la defensa de la colonia (soldados). Dado que las colonias de una especie dada y su etapa de desarrollo tienen una proporción característica de castas, surge la pregunta de si la colonia produce una cantidad elevada de soldados en respuesta a perturbaciones o amenazas repetidas.
Hasta el momento de escritura de este ensayo, el único estudio experimental que se refiere a lo anterior, mostró que las colonias de la hormiga Pheidole pallidula (Nylander) aumentaron la proporción de obreras mayores (soldadas) en presencia constante de una colonia extranjera de la misma especie. En las colonias de control, las soldadas contaron con un promedio de 9,8 % de las obreras, siendo casi el doble que en colonias experimentales (Passera et al., 1996).
Una resolución de esta cuestión en cuanto a los comejenes se efectuará mejor con una especie de Nasutitermes u otra de la misma subfamilia, ya que tienen una proporción elevada de soldados (Fig. 4) (Haverty, 1977; Merritt & Starr, 2010).
A diferencia de las hormigas, los comejenes superiores (Termitidae), como los nasutitermitinos, apenas se pueden mantener en el laboratorio con su nido intacto, lo que no permite observaciones de lo que pasa adentro. Tal investigación tendría que realizarse a través de experimentos de campo. Por tanto, en este ensayo se sugiere lo siguiente:
Identificar un gran número de colonias activas de una misma especie en el mismo lugar. Extraer una muestra de cada una y calcular la ración de soldados/obreros. Dividir las colonias al azar en grupos control y experimental. A intervalos regulares durante algunos meses perturbar las colonias experimentales por destrucción mínima de sus nidos (Fig. 5), sin afectar las colonias de control. Finalmente extraer nuevas muestras para comparar la proporción de soldados en los dos grupos.

Obreros y soldados de Nasutitermes sp. Dos obreros a la derecha tienen un cuerpo generalizado, mientras que cuatro soldados a su izquierda defienden la colonia con compuestos glandulares expuestos de la proboscis. Foto por R. H. Scheffrahn.

Nido de Nasutitermes ephratae (Holmgren) con una parte triangular de la cubierta quitada.
7. ¿Experimentan las poblaciones de avispas sociales una disminución global en la actualidad?
Existe una crisis global en poblaciones de muchos anfibios. Los herpetólogos, al igual que los entomólogos, tienen sus lugares de campo donde regresan año por año. En conversaciones durante el Primer Congreso Mundial de Herpetología en 1989, muchos de los especialistas en anfibios aprendieron que la percibida disminución de sus poblaciones no representaba casos aislados, sino un patrón mucho más amplio. En los años siguientes se estableció que existe en realidad una crisis profunda y mundial en la salud de los anfibios por múltiples causas (Alford & Richards, 1999; Collins & Crump, 2009; Houlahan et al., 2000). Más recientemente se ha levantado el espectro de un patrón semejante en cuanto a los insectos. La gran mayoría de los estudios de largo plazo se han llevado a cabo en Europa y Norteamérica con énfasis en mariposas (Dirzo et al., 2014; Wepprich et al., 2019). Sin embargo, ya emerge evidencia de que se trata de un fenómeno general y mundial (e.g., Hallmann et al., 2017; Sánchez-Bayo & Wyckhuys, 2019).
Parece que los diferentes grupos de insectos no se afectan igualmente (Sánchez-Bayo & Wyckhuys, 2019). Esto levanta la cuestión, hasta ahora apenas investigada, si los insectos sociales en general se han afectados de otra manera que los solitarios a causa de vivir en grupos cooperativos y relativamente densos. Vale notar que la lista roja de la IUCN (Unión Internacional para la Conservación de la Naturaleza) de especies amenazadas incluye informes sobre casi 100 especies de abejorros (Hymenoptera: Bombus) y hormigas (Hymenoptera: Formicidae), y algunas abejas halíctidas (Hymenoptera: Halictidae), pero ninguna avispa social (Baillie et al., 2004).
Sin embargo, en los últimos años, varios especialistas en avispas sociales -incluyéndome- sospechamos que muchas poblaciones están atravesando una etapa de disminución. Por ejemplo, hace 15-20 años, en mi propia región ubicada, en el extremo nor-oriental de Suramérica, era común encontrar al menos de cualquiera de las ocho especies frecuentes por cada hora de búsqueda. Actualmente, sólo una de estas especies mantiene ese nivel de abundancia, las demás se han vuelto difíciles de encontrar en el mismo periodo de tiempo y cantidad. Además, situaciones similares han sido reportadas en comunicaciones directas con varios colegas entomólogos en otras partes del mundo, aunque las causas aún son desconocidas.
Las poblaciones de avispas se monitorean quizás más convenientemente que cualquier otro grupo mayor de insectos sociales, ya que nidifican de una manera relativamente llamativa y las especies se identifican en el campo con relativa facilidad. Por este motivo podemos tener confianza en los datos preliminares representados aquí.
Ejemplos de avispas sociales que eran abundantes en dadas localidades hace 15-25 años, con su estado actual. + más común hoy. 0 ningún cambio sustantivo. - distintamente menos común hoy.
Si resulta una disminución general en las avispas sociales, ¿cuáles serán las causas más importantes? Parece dudoso que el calentamiento global tenga un papel clave. Tampoco que resulte del uso de insecticidas, ya que a) estas se aplican sobre todo en hábitats agrícolas donde hay una abundancia mayor de individuos, pero una diversidad baja, y b) en algunos lugares se observan algunas especies abundantes a pesar de una aplicación fuerte de insecticidas. Además, tal aplicación no es un fenómeno reciente.
8. ¿Existe una relación entre el nivel de producción primaria en el hábitat y la proporción de insectos sociales?
La primera tentativa hacia un concepto general del impacto ecológico de los insectos sociales en general se vio en los preparativos del Programa Internacional Biológico (IBP) de 1964-1974 bajo el liderazgo de Brian (1978). Ya en esa época se reconocieron, por ejemplo, el papel de los comejenes como agentes en la descomposición de toda materia vegetal, las hormigas como los depredadores importantes de invertebrados terrestres, y las abejas como polinizadores eficaces. Desde entonces, el progreso ha sido lento hacia una respuesta concreta a las preguntas ¿Tienen los insectos sociales en su conjunto papeles distintamente suyos en el medioambiente? y ¿Cuál diferencia ecológica surge de que algunos linajes viven en sociedad?. El gran éxito de los insectos sociales se ve fácilmente en su gran abundancia/biomasa y algunos de sus impactos sobre el medioambiente (e.g., Souza & Delabie, 2021). Sin embargo, no es obvia cualquier relación entre su socialidad y su éxito ecológico. En particular, ¿cuáles son las diferencias en cuanto a sus efectos ecológicos entre una colonia de insectos sociales y un número igual de insectos solitarios emparentados? Wilson (1987, 1990, 2020) y Hölldobler y Wilson (2008) han propuesto una respuesta a esta pregunta:
Los insectos sociales predominan numéricamente en los hábitats terrestres. También muestran una dominancia en cuanto a la cosecha de los recursos más valiosos. Esta dominancia resulta precisamente de su socialidad.
En esta teoría los grupos altamente sociales (comejenes, hormigas y algunas abejas) se hallan al pico de la pirámide de dominancia. Como corolario, esta dominancia debe aparecer más en los ecosistemas terrestres de mayor estabilidad y producción, en especial del trópico.
Lo anterior, es una tesis lógica y muy atractiva, y por esta misma razón ha tenido fácil aceptación. Esta afirmación, en su forma actual, presenta una exageración en cuanto a su respaldo cuantitativo. Consideremos la primera parte: una afirmación ampliamente conocida por Wilson (1990) es que “En el bosque tierra firme cerca de Manaos, las hormigas y las termitas en conjunto componen casi 30 % de la biomasa animal según los resultados de Fittkau y Klinge (1973).” Esta cifra ha sido repetida y difundida tantas veces que muchos biólogos la consideran como un dato altamente comprobado para todas las selvas tropicales en general (e.g., Bradley, 2009), a pesar de que el estudio se refiere a un contexto específico.
Sin embargo, la realidad es otra. Si se examina esta afirmación en las obras de varios colegas, resulta que se basa en este solo artículo de hace casi 50 años (Fittkau & Klinge, 1973) casi sin replicación ni ampliación (e.g., Stork, 1987). Aunque bien plausible, no se sabe a ciencia cierta que los insectos sociales formen una mayor proporción de individuos o biomasa en los ecosistemas tropicales en comparación con los subtropicales y templados. Además, el enfoque principal de dicha investigación estaba centrado en la vegetación, y los datos sobre fauna —incluidos los referentes a la biomasa animal— fueron tratados de forma incidental, sin un diseño específico para su estudio detallado. Fittkau y Klinge (1973) dijeron abiertamente que “La falta de datos exactos sobre la densidad poblacional y biomasa de comejenes y hormigas —tan abundantes en el medioambiente de la Amazonía central— es lamentable”. Solamente podemos hacer referencia a la conjetura de Beck (1971) de que los tres cuartos de la fauna del suelo en la selva de la Amazonía central, en cuanto a su biomasa, son hormigas y comejenes. Por lo tanto, nos queda imposible evaluar el papel de estos animales del suelo en el flujo de energía en la selva de la Amazonía central.
Hoy tenemos un estimativo de la biomasa mundial de las hormigas, con el resultado poco sorprendente que es mayor en regiones tropicales y subtropicales (Schultheiss et al., 2022). Lo que no sabemos ni aproximadamente es como la biomasa de insectos sociales y solitarios varía geográficamente. Esta falta de conocimiento se convierte en una oportunidad para la ciencia organizada por lo que se sugiere que las sociedades entomológicas de esta región, incluida la Sección Andina y del Caribe de la UIEIS, piensen en organizar una investigación colaborativa sobre esta cuestión, de tal manera que se observe un crecimiento del conocimiento relacionado para el próximo período.
Es una cuestión temáticamente sencilla que podría resolverse, por ejemplo, por una comparación entre bosques del mismo tipo (seco o mojado) a diferentes latitudes de especies e individuos de a) comejenes y cucarachas, y/o b) avispas o abejas sociales y solitarias. Lo anterior partiendo de que el nivel de producción primaria en cada hábitat es conocido. Tal estudio serviría como un primer paso hacia resolver la pregunta ¿Qué tiene la socialidad que ver con el nivel de control del ecosistema y sus recursos?
Origen y financiamiento
Este artículo no tuvo financiación externa.
Conflicto de intereses
Los autores declaramos no tener conflicto de intereses.
Footnotes
Agradecimientos
Este ensayo se dedica a Edward O. Wilson (1929-2021; Harvard University, EE.UU.), pionero en el estudio de insectos sociales. La primera versión de ello se presentó poco antes del 90vo cumpleaños de Wilson como una conferencia de la Unión Internacional para el Estudio de Insectos Sociales (Sección Andina y del Caribe), cuyos socios agradezco por sus comentarios. El Editor y los revisores de la revista también contribuyeron sugerencias valiosas. La ![]() fue preparada por Nova Y. Starr (Toronto, Canadá).
fue preparada por Nova Y. Starr (Toronto, Canadá).