
Abstract
In this study, was evaluated through traditional morphometry, the effect of tem- perature on the phenotypic expression of size, elongation (length/width) and costal wing spots among families, progenies and sexes in populations of Nyssorhynchus triannulatus from southern Colombia. In the progenies of 11 families, obtained at 20, 24 and 28 °C the wings showed four costal spots patterns. Pattern I was present in all three breeding tempera- tures with the highest frequency. Within families, the average length/width of wing ratio was significantly higher in males than females, while the variation by sex of this character was similar. Among the families, the temperature had variable influence on the sizes of wings and costal spots. Within the components of phenotypic variation, the breeding temperature had no significant detectable effect on the length of the wing and several of the analyzed characters; however, the proportion of the subcostal pale spot length over the length of the distal sector dark spot (SCP/DSD) was the most sensitive to temperature changes. The genetic contribution of the wing size and the measured characters length, as well as the effect of the families (origin) on the phenotypic variation, was significant for almost all the characters analyzed. The effect of the sexes on the phenotypic variation of the studied characters was significant only for some characters, while there were few significant inter- actions between the three factors analyzed (temperature, family and sex) with a low effect on the phenotypic variation.
Introducción
En taxonomía, es indudable que los caracteres discretos fa- cilitan el proceso de las diagnosis específicas de los organis- mos biológicos, sin embargo, frecuentemente es necesario hacer uso de caracteres continuos, especialmente cuando se trata de especies estrechamente relacionadas. Esto último, es el caso de especies de mosquitos Nyssorhynchus, considera- dos aquí según la propuesta de Foster et al. (2017), quienes elevaron a género a este y otros subgéneros de Anopheles. En estos, cuando se trata de situaciones taxonómicas confu- sas, se emplean marcadores moleculares, de los cuales el có- digo de barras de ADN (COI) es uno de los más usados debido a la disponibilidad de cebadores universales que amplifican a través de un amplio rango de especies, su baja divergencia intraespecífica pero alta divergencia interespecífica (Bourke et al. 2018) (para Anophelinae, ver González et al. 2010; Gó- mez et al. 2015; Scarpassa et al. 2016; Saraiva et al. 2018). Sin embargo, no siempre se dispone de la facilidad logística de laboratorios para realizar estas pruebas moleculares y, por lo tanto, la información morfológica sigue teniendo un gran peso en la diferenciación de las especies de Anophelinae. En las caracterizaciones morfológicas, es común el uso de tasas que relacionan caracteres próximos, especialmente de las alas y patas posteriores (Faran 1980; Faran y Linthi- cum 1981; Linthicum 1988; González Obando y Carrejo Gironza 2009). Sin embargo, por la naturaleza de los datos discontinuos, especialmente para especies de un mismo gé- nero, subgénero, grupos o complejo de especies próximas, no se presenta una discontinuidad clara entre las tasas em- pleadas, dado que estas son variables entre los individuos de una misma población e incluso entre las progenies de una misma familia.
Debido a lo anterior, para la determinación específica del género Nyssorhynchus, generalmente se hace uso de informa- ción morfológica combinada y asociada de larvas, hembras y genitalia de machos, obtenidos a partir de la producción de isofamilias. Pero, no siempre se puede tener disponibilidad de estos estados y lo más práctico es la obtención de hem- bras. En estas, la mayor parte de la información diagnóstica de especies se logra principalmente a partir, de las manchas claras y oscuras del área costal de las alas. En el proceso de búsqueda de métodos apropiados para la diferenciación de especies a partir de la información de las alas de las hem- bras, el análisis de la forma de estas ha sido una opción. El empleo de morfometría geométrica ha tenido relativa uti- lidad en la diferenciación de especies crípticas de Anopheles (Petrarca et al. 1998; Calle et al. 2002; Gómez et al. 2013; Pacheco et al. 2017; Pacheco-Gómez et al. 2018). Sin em- bargo, la morfometría tradicional parece ser una herramienta de fácil aplicación por personal entrenado, bajo costo y gran potencial para la exploración de la diversidad de especies (Cazorla 2009). Además, ha sido de gran utilidad para es- clarecer problemas taxonómicos en especies de difícil diag- nóstico, describir variaciones morfológicas en poblaciones a diferentes altitudes y diferenciar poblaciones provenientes de diferentes ecorregiones (Belen et al. 2004; Bustamante et al. 2004; Dantur Juri et al. 2011; Dos Santos Silva et al. 2012; Rodríguez-Zabala et al. 2016). Por otro lado, para en- tender cómo se regula el desarrollo de los caracteres de un organismo hasta alcanzar un fenotipo ideal, la morfometría tradicional es una de las herramientas de estudio, que utiliza métodos estadísticos multivariados a colecciones de varia-bles de tamaño o conformación, en la que se definen medicio- nes con el fin de registrar aspectos biológicos significativos del organismo.
En el estudio de la variación morfológica, los cambios fenotípicos dependen de factores como pH, humedad, tem- peratura, fotoperiodo, estacionalidad, entre otros (Müller 2007), lo cual para especies de Culicidae se traduce en plas- ticidad fenotípica. De acuerdo con Motoki et al. (2012) las diferencias ambientales entre ecorregiones podrían conducir a diferenciación fenotípica de poblaciones de Anophelinae, expresada en cambios químicos, fisiológicos, del desarro- llo, morfológicos, de conducta, entre otros (Fusco y Minelli 2010).
En especies de Anophelinae, las alas presentan una serie de caracteres fenotípicos variables analizados por morfometría tradicional, ya sea dentro y entre poblaciones y progenies de isofamilias (Hribar 1995; Fajardo Ramos et al. 2008; Pacheco et al. 2017; Pacheco-Gómez et al. 2018) y entre especies (Ro- sa-Freitas et al. 1990; Rubio-Palis 2000). Mediante este mé- todo, la información de las alas puede ser usada para estudiar el aporte del componente genético a la variación fenotípica. La variabilidad en el tamaño del ala y en la longitud de las manchas costales, puede ser utilizada para analizar el aporte del componente genético a la variación fenotípica, el grado de canalización, de integración morfológica y de asimetría fluc- tuante presentes en respuesta a cambios ambientales como la temperatura. Para obtener información de estos caracteres, es fundamental la realización de experimentos de ecofisiología, donde se pueda establecer control de las variables a evaluar (por ejemplo, temperatura), manteniendo constantes el resto de las variables (humedad relativa, fotoperiodo, alimentación, densidad larval, entre otras).
Considerando lo anterior, más allá de lo meramente ta- xonómico, el uso de métodos morfométricos puede ser tam- bién aprovechado para medir el efecto ambiental sobre la expresión fenotípica de caracteres morfológicos, pero has- ta el momento, no se conocen trabajos publicados con los mosquitos como modelos biológicos para estudiar cómo se descompone la variación fenotípica de la forma, el tamaño y los patrones de manchas por efecto de cambios ambien- tales como la temperatura. En el presente trabajo, mediante morfometría tradicional, se evaluó el efecto de la tempera- tura en la expresión fenotípica del tamaño, alargamiento de las alas y longitud de las manchas costales entre familias, progenies y sexos, utilizando como organismo modelo a Nyssorhynchus triannulatus (Neiva y Pinto, 1922). Median- te este método, en esta especie se estimó los aportes de la variación ambiental (efecto de la temperatura), del compo- nente genético (variación entre familias) y la interacción genotipo-ambiente sobre la variabilidad fenotípica de los ca- racteres descritos arriba.
Materiales y métodos
Las hembras de anofelinos fueron recolectadas en el mu- nicipio de Puerto Asís, Putumayo, Colombia (0°30'02″N 76°29'56″O) sobre cebo humano protegido y con búsqueda activa del mosquito en reposo en el intradomicilio y vegeta- ción del peridomicilio. Los mosquitos se recolectaron entre las 18:00 y 24:00 horas, de tres a cinco días consecutivos en dos salidas (en noviembre 2018 y abril 2019) con un aspi- rador bucal y se depositaron en recipientes plásticos de 300 ml, hasta un máximo de 25 a 30 individuos/recipiente. Se les transportó en cajas térmicas al Laboratorio de Entomología de la Universidad del Valle, en Santiago de Cali, con suministro de azúcar diluida en agua en un algodón, para obtener isofa- milias y realizar la identificación respectiva.
Para la obtención de las progenies, cada una de las hem- bras recolectadas se colocó en una cámara de ovipostura. Cada cámara estuvo compuesta de un vaso plástico desechable de 200 cm3 de capacidad, con una capa de algodón húmedo en el fondo y sobre éste, papel filtro blanco acondicionado para la oviposición. Cada hembra se alimentó con sangre de cobayo y agua azucarada. Las cámaras se almacenaron en cajas térmicas acondicionadas para el mantenimiento de alta humedad relativa y una temperatura promedio de 24 °C. A partir del segundo día se realizaron observaciones diarias para identificar posturas de huevos. Cada postura/hembra se incubó separadamente en bandejas plásticas de 25 x 30 x 5 cm3, mantenidas en una cámara ambientada SANYO, modelo MLR 351H, regulada en el tiempo a 20, 24 y 28 °C. En cada temperatura, las larvas se criaron en agua reposada hasta al- canzar el estado adulto, manteniendo constantes la humedad relativa, la alimentación y un fotoperiodo de 12 horas luz/ noche. Las larvas de los estadios I y II se alimentaron con le- vadura de cerveza y las de estadios III y IV con una combina- ción de polvo de alimento para conejos y levadura de cerveza. Las pupas se individualizaron en vasos plásticos de 100 cm3 cubiertos con tul hasta obtener los respectivos adultos. Los adultos se alimentaron con solución azucarada por dos días y posteriormente se sacrificaron en frio y se preservaron en alfileres entomológicos. Cada ejemplar se rotuló con el códi- go de su respectiva postura. Nyssorhynchus triannulatus s.l. se identificó por los caracteres diagnósticos de las hembras y posteriormente se verificó a partir de la genitalias de los ma- chos obtenidos de cada isofamilia. Las genitalias de machos se procesaron y preservaron en placas, siguiendo el método de González Obando y Carrejo Gironza (2009). Esta espe- cie podría corresponder al linaje nombrado por Moreno et al. (2013) como Nys. triannulatus E, dado que el sitio de mues- treo se ubicó próximo a poblaciones amazónicas de Ecuador y del departamento colombiano de Amazonas. Sin embargo, es importante tener presente que Nyssorhynchus triannulatus es considerado un complejo en el cual se reconocen almenos tres especies hermanas, Nys. triannulatus s.s. (Neiva y Pinto), Nys. halophylus (Silva do Nascimento y Lourenço-de-Olivei- ra 2002) y Nys. triannulatus C (Silva-do-Nascimento et al. 2006; Moreno et al. 2013). Este último no ha sido caracteri- zado morfológicamente, pero en lo que respecta a las dos pri- meras, se pueden diferenciar por caracteres morfológicos de los genitales de machos, formas setales y aparato espiracular de larvas de cuarto estadio y aspecto general del huevo (Silva do Nascimento y Lourenço-de-Oliveira 2002).
Las alas derechas e izquierda y una de las patas posterio- res de cada ejemplar se preservaron en una placa portaobjeto utilizando un cubreobjeto de 22 mm, fijado con bálsamo de Canadá. La asignación y medición de las diferentes manchas costales se realizó siguiendo la nomenclatura propuesta por Wilkerson y Peyton (1990). Los análisis por morfometría tradi- cional se basaron en un registro fotográfico de alas (aproxima- damente 20 fotografias/ala) empleando una cámara fotográfica digital Canon Eos Rebel T5i adaptada a un microscopio Nikon modelo E-Ci, y el software Helicon Focus 5.3. (Helicon Soft Ltd.). Después, se procedió a la toma de medidas con ayuda del programa CorelDRAW Graphics Suite X7, midiendo la longi- tud, el ancho y los tamaños de las manchas costales del ala. Se usó la siguiente nomenclatura durante la toma de medidas las alas (Fig. 1): Prehumeral oscura (PHD), Humeral clara (HP), Humeral oscura (HD), Presector claro (PSP), Presector oscu- ro (PSD), Sector claro (SP), Sector oscuro proximal (PRSD), Sector claro accesorio (ASP), Sector oscuro distal (DSD), Sub- costal clara (SCP), Preapical oscura (PD), Preapical clara (PP), Apical oscura (AD), Apical clara (AP).
En las patas posteriores, se midió la longitud del tarsóme- ro dos (ta-III2) y área oscura basal del mismo. También, se calcularon las tasas alargamiento (longitud/ancho), SCP/DSD (longitud de la mancha subcostal clara)/(longitud de la man- cha sectorial oscura distal) y HP/PHD (longitud de la mancha clara humeral)/(longitud de la mancha oscura prehumeral). Además, se consideró la suma de las longitudes de las man- chas del sector claro (SP), sector oscuro proximal (PRSD) y sector claro accesorio (ASP), SP + PRSD + ASP. Para las pa- tas posteriores se usó la tasa de la longitud de la parte oscura basal del tarsómero dos (ta-III2) sobre la longitud del tarsóme- ro. Los datos se tabularon en una hoja de cálculo del programa Microsoft Excel para Office 365. Con el fin de aproximar la distribución de los datos a una distribución normal, previo a los análisis estadísticos, se procedió a su transformación usan- do la fórmula de Freeman y Tukey (1950). Para tasas se utili- zó la ecuación de Kihlberg et al. (1972). Los análisis descrip- tivos presentan el promedio, la desviación estándar y valores mínimos y máximos.
Para probar si la temperatura influyó en la variación de los rasgos estudiados se escogieron tres niveles del factor tempe- ratura (20, 24 y 28 °C) y se sometieron las progenies de cuatro isofamilias a 20 °C y 24 °C y tres a 28 °C. Para estimar el efecto del componente genético sobre la variación fenotípica de los rasgos estudiados se asumió que las hembras grávidas recolectadas fueron monoandricas y que el parentesco entre la descendencia de una sola hembra es de 0,5 (Tripet et al. 2003; Shutt et al. 2010; Richardson et al. 2015). Es decir, las progenies derivadas de cada hembra grávida fueron mayorita- riamente hermanos completos. En consecuencia, se estimó la heredabilidad en sentido amplio (H2) utilizando un modelo de hermanos completos, como lo describen Falconer y Mackay (1996) con algunas modificaciones. Por lo tanto, se consideró a las isofamilias como un segundo factor (genético) que afecta la variación fenotípica de los caracteres estudiados.
Por último, se incluyó como un tercer factor, el sexo, dado que dentro de las progenies por familia se obtienen hembras y machos desarrollados bajo las mismas condiciones dentro de la isofamilia. Con este factor se evaluó si las progenies presentaban dimorfismo sexual para los caracteres estudiados, usando análisis de variación (ANDEVA). Se probó la hipóte- sis en la que se consideró que los promedios para la longitud, la tasa largo/ancho y las longitudes de las manchas costales del ala derecha y la tasa PB/ta-III, dentro de isofamilias son iguales entre progenies hembras y machos, asumiendo que las variaciones de las progenies hembras y machos son desco- nocidas pero similares. Para el contraste de medias se usó la prueba t-Student; posteriormente se realizaron comparaciones múltiples con corrección de Bonferroni para probar cada hi- pótesis individual bilateral (H0 = µ1 - µ2 = 0) en un nivel signi- ficativo de a/m, donde a es el nivel de significancia deseado (0,05) y m es el número de hipótesis (Bland y Altman 1995).
Dado que, a partir del modelo general ninguno de los aná- lisis de variación mostró efectos de interacción significativa- mente diferentes de cero, el modelo se ajustó a uno de tres factores sin interacción: yijk= µ + αi+ βj + γk + µijk. Para probar este modelo, se usaron análisis lineales generalizados mix- tos, considerando como variable dependiente las medidas morfométricas y como fuentes de variación la temperatura, la procedencia de las progenies (familias) y el sexo. También se usó la prueba de Levene (1960) W, para evaluar la igualdad de las variaciones para cada una de las variables analizadas, cal- culada para dos o más grupos (3 temperaturas, 11 familias).
Resultados
En las 11 familias de Nys. triannulatus, criadas en las tres temperaturas, se registraron 148 ejemplares (98 hembras y 50 machos) (Tabla 1). Solo cuatro patrones fueron observados, dos a 20 °C, tres a 24 °C y cuatro a 28 °C. El patrón I corres- pondió a la presencia de las manchas costales nombradas en la nomenclatura propuesta por Wilkerson y Peyton (1990) (Fig. 1A). La no observación de la accesoria clara (ASP) definió los patrones II y III. El patrón II correspondió a la extensión del sector claro (SP) con desaparición del sector oscuro proximal (PRSD) y al menos parte de la ASP, es decir que ésta com- prende un área clara grande, la fusión de las manchas SP + PRSD + ASP (Fig. 1B). El patrón III (Fig. 1C) se identificó por la ausencia de la mancha ASP la cual fue ocupada por ex- tensiones del sector oscuro (SD); en este caso, SP se observó más pequeña que en el patrón II y algo similar a la del patrón I. El patrón IV ocurrió cuando el sector claro (SP) fue ocu- pado por extensiones del presector oscuro (PSD) y el sector oscuro proximal (PRSD) (Fig. 1D), es decir que la ASP sigue siendo observable, pero el sector claro no. Tanto en hembras como en machos, el patrón costal I estuvo presente en las tres temperaturas evaluadas y fue el más frecuente (Fig. 2), sobre todo a 24 °C, donde alrededor del 95% de las progenies pre- sentaron este tipo de alas, o sea que pocas veces se observó fusión de algunas de las manchas costales, o por lo menos ausencia de la accesoria clara (ASP). El tipo II también estuvo presente en las tres temperaturas, aunque no en los machos de especímenes criados a 28 °C. El patrón III solo se observó en ejemplares obtenidos a 24 y 28 °C, mientras que el patrón IV solo se observó a 28 °C.
Patrones de manchas costales y frecuencias observadas en progenies de Nyssorhynchus triannulatus, criados en cámaras ambientales a tres temperaturas diferentes.
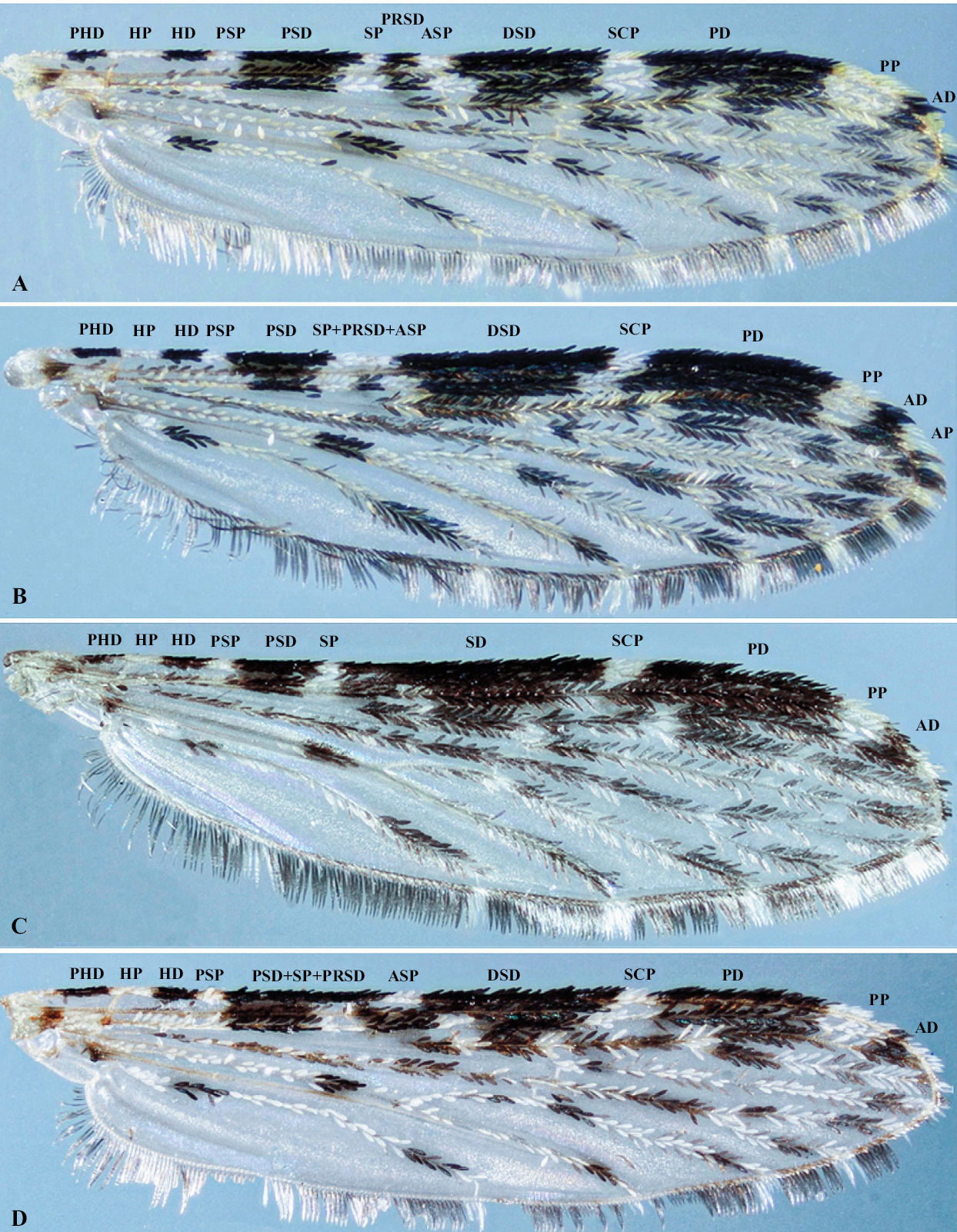
Patrones costales observados en progenies de Nyssorhynchus triannulatus criados en cámaras ambientales a tres tempera- turas diferentes. A. Tipo I (Sin fusión de manchas costales). B. Tipo II (SP + PRSD + ASP). C. Tipo III (ausencia de ASP). D. Tipo IV (PSD + SP + PRSD). Nomenclatura: Prehumeral oscura (PHD), Humeral clara (HP), Humeral oscura (HD), Presector claro (PSP), Presector oscuro (PSD), Sector claro (SP), Sector oscuro proximal (PRSD), Sector claro accesorio (ASP), Sector oscuro distal (DSD), Subcostal clara (SCP), Preapical oscura (PD), Preapical clara (PP), Apical oscura (AD), Apical clara (AP).
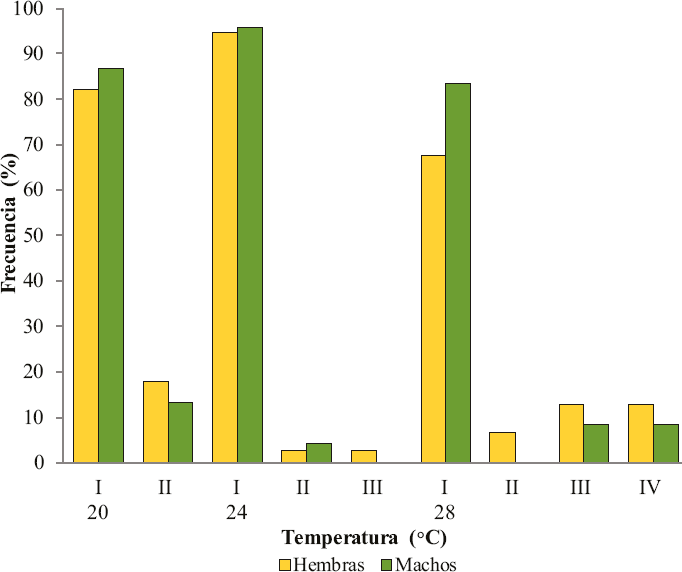
Frecuencias de los patrones de manchas costales observados en hembras (N = 98) y machos (N = 50) de progenies de Nyssorhynchus triannulatus, criados en cámaras ambientales a tres temperaturas.
El porcentaje esperado de patrones II + III + IV fue 14,9% en cualquiera de las tres temperaturas. En ambos sexos, el porcentaje observado a 20 °C fue similar al esperado (hem- bras = 17,9%; machos = 13,3%); a 24 °C, fue menor que el esperado (hembras = 5,1%; machos = 4,3 %) mientras que a 28 °C se incrementó significativamente en hembras y fue similar en machos (hembras = 32,3%; machos = 16,7%). La Figura 2 muestra claramente esta tendencia.
La longitud de las alas derechas de progenies hembras y machos, mostró tamaños muy variables entre familias de in- dividuos criados a la misma temperatura (Tabla 2). En hem- bras, las variaciones a 20 °C, varió entre 2,13 y 3,22 mm (
Longitud de las alas derechas (mm) de Nyssorhynchus triannulatus criados en cámaras ambientales a tres temperaturas: 20 °C (familias 1-4), 24 °C (familias 5-8) y 28 °C (familias 9-11) considerando la procedencia de las progenies (familias) y el sexo.
D. E.: Desviación estándar; Temp.: Temperatura.
La comparación de la longitud promedio de las alas dere- chas de las progenies hembras (H) y machos (M) con respecto a la temperatura, no mostró diferencias significativas en nin- guna de las tres comparaciones. En hembras, los valores de t-student para las parejas 20 - 24 °C, 20 - 28 °C y 24 - 28 °C, fueron respectivamente: 0,44 (p = 0,6619), 1,43 (p = 0,1589) y 1,42 (p = 0,1615) mientras que en machos fueron 0,39 (p = 0,6992), 0,81 (p = 0,4264) y 1,42 (p = 0,1653).
La longitud promedio de las alas en las progenies hem- bras fue significativamente mayor que la de machos en las familias 1-4 criadas a 20 °C, en la familia 5 criada a 24 °C y en la familia 11 criada a 28 °C (6/11 = 54,5 %) (Tabla 3). Lo contrario se observó para las familias 6 y 7 criadas a 24 °C (2/11 = 18,2%).
Pruebas de t-student con su significancia (p) para comparar longitud promedio de progenies hembras (H) y machos (M) en cada familia de Nyssorhynchus triannulatus criadas a 20 °C, 24 °C y 28 °C.
n1: Número de hembras por familia; n2. Número de machos por familia.
No hubo diferencias significativas en la longitud prome- dio de progenies hembras y machos en la familia 8 criada a 24 °C y en las familias 9 y 10 criadas a 28 °C (3/11 = 27,3 %).
Tampoco se presentaron diferencias significativas en la longitud promedio de progenies hembras y machos en la fa- milia 8 criada a 24 °C y en las familias 9 y 10 criadas a 28 °C (3/11 = 27,3%).
La prueba de Levene (1960) W, utilizada para evaluar la igualdad de las variaciones en la longitud del ala, mostró que las progenies hembras y machos criadas a 20 °C tuvieron una variación significativamente mayor que a 24 y 28 °C (H: W = 10,351; p < 0,0001 y M: W = 11,291; p < 0,0001).
En el ANDEVA realizado para evaluar los efectos produ- cidos por la temperatura, la procedencia de las progenies ana- lizadas (familias) y el sexo sobre la variación en la longitud del ala derecha (Tabla 4) se observó que la temperatura (F = 338,6603; p < 0,0001) y la familia (F = 16,1903; p < 0,0001) tienen efectos significativos mientras que el sexo no (F = 1,4213; p = 0,2356). Al descomponer la variación fenotípica de la longitud del ala derecha, el factor temperatura, hizo el mayor aporte a la variación (96,5%), seguido del factor fami- lia con 2,9% (Tabla 4).
Componentes de la variación fenotípica (%), prueba F para contrastar variación y su significancia (p), para los caracteres longitud, tamaños de manchas costales, de la fusión (SP + PRSD + ASP) y las proporciones del ala derecha de progenies hembras y machos de Nyssorhynchus triannulatus, considerando como factores las temperaturas (T) de crianza (20, 24 y 28 °C), las procedencias (familias = F) y sexo (S) con las interacciones significativas (Temperatura*Sexo = TS y Familia*Sexo = FS).
La tasa largo/ancho de las alas derechas fue variable entre las progenies de las diferentes familias y se mostró como una característica importante para describir el alargamiento del ala. Los análisis descriptivos se presentan en la Tabla 5. Con excepción de la familia tres, la tasa media del largo/ancho de las alas derechas fue mayor en machos que en hembras. Hay un claro dimorfismo sexual para el alargamiento promedio del ala. El alargamiento promedio en las progenies hembras fue significativamente menor que la de las progenies machos tanto en las familias 2, 6-9 (Tabla 6) como cuando se conside- raron todas las progenies (t-student = - 5,1364; p < 0,0001).
Tasa largo/ancho de las alas derechas de Nyssorhynchus triannulatus criadas en cámaras ambientales, a tres temperaturas: 20 °C (familias 1-4), 24 °C (familias 5-8) y 28 °C (familias 9-11) considerando la procedencia de las progenies (familias) y el sexo.
D. E.: Desviación estándar; Temp.: Temperatura.
Pruebas de t-student con su significancia (p) para comparar la tasa promedio largo/ancho del ala derecha de progenies hembras y machos en cada familia de Nyssorhynchus triannulatus criadas a 20, 24, 28 °C.
n1: Número de hembras por familia; n2: Número de machos por familia;
El alargamiento promedio en hembras fue 3,575 ± 0,232 mientras que en machos fue 3,798 ± 0,2010. Este dimorfismo, también se mantuvo considerando cada temperatura de cría de las larvas. Los valores de t-student fueron: - 3,0810 a 20 °C (p = 0,0041), - 5,0455 a 24 °C (p < 0,0001) y - 2,7638 a 28 °C (p = 0,0098). Estos resultados sugieren que las alas de los machos son significativamente menos anchas que las de las hembras (Tablas 6 y 7). También se puede observar que el 70% de las tasas largo/ancho del ala derecha en hembras se registra entre 3,50 y 3,70 mientras que, en machos el 68,0 % se observa entre 3,9 y 4,1 (Fig. 3).
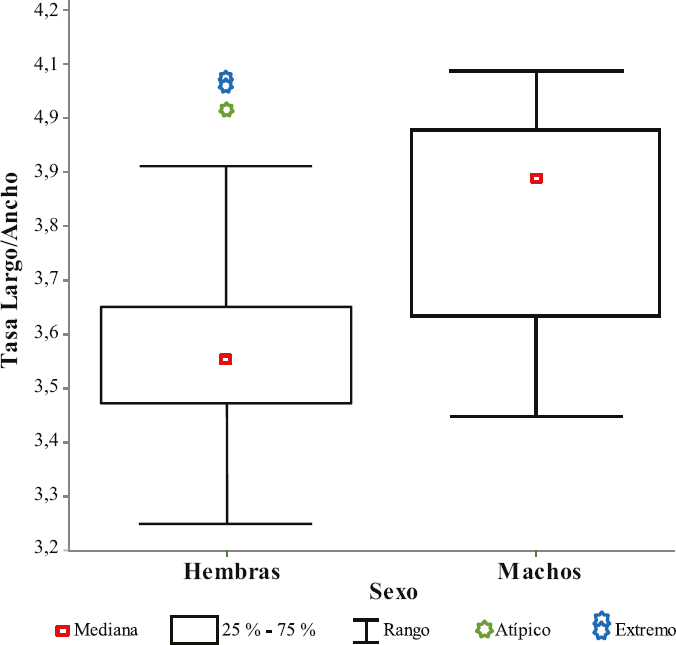
Diagrama de caja mostrando la distribución de las tasas lar- go/ancho de las alas derechas de hembras y machos de Nyssorhynchus triannulatus. En resumen, se indican cinco números: “mínimo”, primer cuartil (Q1), mediana, tercer cuartil (Q3) y “máximo”. También se indi- can los valores atípicos observados en hembras (4 entre 4,0 y 4,1).
En el ANDEVA para evaluar los efectos producidos por la temperatura, la procedencia de las progenies (familias) y el sexo sobre la variación en la tasa largo/ancho del ala dere- cha (Tabla 4) se observó que la temperatura (F = 616,7516; p < 0,0001), la familia (F = 41,02; p < 0,0001) y el sexo (F = 28,72; p = 0,0004) tienen efectos significativos. Al descomponer la variación fenotípica de la tasa largo/ancho del ala derecha, el factor temperatura hizo el mayor aporte a la variación (98,0%), seguido del factor familia con 1,6 % y por último el sexo con 0,1% (Tabla 4). Al igual que la variación en la longitud total del ala derecha, la tasa largo/ ancho fue susceptible a cambios ambientales, en este caso, a la temperatura de cría de las progenies. Igualmente, la H2 estimada en sentido amplio fue baja (2 * 1,6 % = 3,2 %). Esto significa que este carácter proporcionalmente tiene un componente genético bajo, lo cual posiblemente se deba a que los individuos dentro de las familias fueron muy variables y, por ende, el residual se hace proporcionalmente mayor res- pecto al componente entre familias de individuos criados a la misma temperatura (Tabla 4).
En el Anexo 1 se indican los promedios con sus respec- tivas desviaciones estándar (D. E.) de la longitud de las alas derechas, de los tamaños de las manchas costales, de las pro- porciones promedio y de la tasa de la mancha oscura basal/ longitud del tarsómero posterior dos (PB/ta-III2) en proge- nies hembras y machos de Nys. triannulatus, criadas a tres temperaturas: 20, 24 y 28 °C. Con base en el Anexo 1 se con- firmó la existencia de dimorfismo sexual en cada una de las temperaturas de crianza (Tabla 5). Entre el 80 y 85% de los caracteres mostraron diferencias significativas entre sexos. En cualquiera de las tres temperaturas de crianza, la longitud promedio del ala es significativamente mayor en hembras respecto a machos, mientras que los promedios de los tama- ños de SCP y PP y de la tasa SCP/DSD fueron menores en hembras respecto a machos. A 20 °C, los promedios de la longitud de las manchas BP + PHP, SP, PRSD, de la fusión (SP + PRSD + ASP) y de la tasa HP/PHD fueron significa- tivamente mayores en hembras respecto a machos, mientras que las manchas PHD, HP, HD, PSD, ASP, SCP, PD, PP, AD y AP, las tasas SCP/DSD y PB/ta-III, fueron significativa- mente mayores en machos respecto a hembras.
A 24 °C, los promedios de la longitud de las manchas BP + PHP, PSP, PRSD, DSD, PD, AD y AP, la tasa HP/PHD y de la tasa PB/ta-III, fueron significativamente mayores en hembras respecto a machos, mientras que las manchas PHD, HP, HD, PSD, SP, SCP y PP, la tasa SCP/DSD y la longitud de la fusión (SP + PRSD + ASP) mostró lo contrario. A 28 °C, los promedios de la longitud de la mancha PD, de la tasa HP/PHD fueron significativamente mayores en hembras res- pecto a machos, mientras que las manchas BP + PHP, PHD, HP, HD, PSP, SP, ASP, SCP, PP y AP, las tasas SCP/DSD y PB/ta-III, y la fusión (SP + PRSD + ASP) en los machos fueron significativamente mayores respecto a hembras. Al parecer, a esta temperatura de cría, en machos se favorece el aumento de la longitud de las manchas claras sobre las manchas oscuras.
Los tamaños promedio de las manchas costales PRSD y DSD fueron mayores en hembras respecto a machos a cual- quiera de las tres temperaturas de crianza, pero solo significa- tivas a 24 °C. La tasa HP/PHD fue mayor en hembras respec- to a machos, pero esta diferencia no fue significativa a 24 °C. Los tamaños promedio de PHD, HP y HD fueron menores en hembras respecto a machos a cualquiera de las tres tempera- turas de crianza, pero no fue significativo para HP a 24 °C. El tamaño promedio de la mancha PSD fue menor en hem- bras respecto a machos, pero esta diferencia no fue signifi- cativa a 28 °C. El tamaño promedio de la mancha BP + PHP inicialmente significativamente mayor en hembras respecto a machos a 20 °C y 24 °C, se hace significativamente menor a 28 °C. También muestra esta tendencia PSP. El tamaño promedio de la mancha SP que a 20 °C es significativamente mayor en hembras respecto a machos, se hace significativa- mente menor a 24 °C y 28 °C, triplicándose esta diferencia a 28 °C.
La fusión (SP + PRSD + ASP) también muestra esta ten- dencia. El tamaño promedio de la mancha AD que a 20 °C es significativamente menor en hembras respecto a machos, se hace significativamente mayor a 24 °C y 28 °C. El tamaño promedio de la mancha PD es significativamente menor en hembras respecto a machos a 20 °C, mayor a 24 °C y vuelve a ser significativamente menor a 28 °C. También muestra esta tendencia la tasa PB/ta-III2.
Las pruebas estadísticas por cada una de las temperaturas de crianza por familia se muestran en el Anexo 2. Entre el 68 y el 95% de la longitud promedio de los caracteres analiza- dos mostraron diferencias significativas entre las progenies hembras y machos. De estas diferencias, entre el 16 y el 60 % mostraron longitudes promedio de las hembras mayores que las de los machos y del 40 al 84% de las comparaciones mostraron lo contrario. De las 11 familias analizadas, siete mostraron la longitud promedio de las alas de hembras sig- nificativamente mayor que las de los machos (7/11 = 0,636), mientras que las manchas HP (10/11 = 0,909), PSD (8/11 = 0,727), SCP (9/11 = 0,818), PP (7/11 = 0,636) mostraron longitudes promedio de las manchas costales de los machos significativamente mayores que las de las hembras. La tasa promedio de SCP/DSD en las progenies machos fue signi- ficativamente mayor que las de las hembras en 7 de las 11 familias (7/11 = 0,636). Esto se debe principalmente a que la longitud promedio de la mancha costal SCP es significa- tivamente mayor que la de las hembras en 9 de 11 familias analizadas.
En las progenies criadas a 20 °C, la relación de las longi- tudes promedio de DSD y SCP varia, por ejemplo, en las fa- milias 1 y 2 la diferencia entre sexos (H-M) es positiva para DSD y negativa para SCP mientras que en las familias 3 y 4 ambas son negativas y en la familia 4 no se observa dimor- fismo sexual en estas manchas. En cambio, en las progenies criadas a 24 °C la diferencia entre sexos (H - M) es positiva para DSD y negativa para SCP en las 4 familias estudiadas y son significativas para el dimorfismo sexual. En las proge- nies criadas a 28 °C, en la familia 9, la diferencia entre sexos es negativa y significativa para DSD y positiva no significa- tiva para SCP; en la familia 10 es positiva para DSD y nega- tiva para SCP, ambas significativas; y en la familia 11 DSD y SCP son negativas y significativas para dimorfismo sexual. Esta variación en la dirección del dimorfismo sexual a tem- peraturas de 20 °C y 28 °C indica el efecto que la tempe- ratura sobre las longitudes de las manchas de este sector. Parece ser que la temperatura de 24 °C es óptima debido a que muestra mayor estabilidad fenotípica (Tabla 7, Anexos 1 y 2).
Diferencias en las longitudes de alas, manchas y proporciones de manchas costales y tarsómero posterior dos, en progenies hembras y machos de Nyssorhynchus triannulatus, criados a tres temperaturas. Comparación de medias de hembras y machos dentro de cada temperatura (°C) mediante Pruebas t-student y su significancia (p < 0,05/2 = 0,025 para las proporciones).
E.E.(p).: Error estándar del promedio (hembras + machos); (*) A partir de datos transformados según Freeman y Tukey (1950) y Kihlberg et al. (1972).
La tasa promedio de PB/ta-III, mostró dimorfismo se- xual significativo en ocho de nueve familias (8/9 = 0,889), en cinco fue significativamente mayor en hembras que en machos (5/8 = 0,625) y en tres lo contrario (3/8 = 0,375). Este dimorfismo sexual no se asocia significativamente con la temperatura de cría de las progenies (- 1,5156 ≤ t ≤ 0,0188; 0,1397 ≤ p ≤ 0,9885).
El mayor dimorfismo sexual se detectó en la familia 11 con 19 caracteres con diferencias significativas entre sexos (19/20 = 0,95). De estas diferencias el 16 % mostró longi- tudes promedio de hembras significativamente mayores que la de los machos y el 84% restante lo contrario. El menor dimorfismo sexual se detectó en la familia 7 con 12 caracte- res con diferencias significativas entre sexos (12/20 = 0,60). De estas diferencias el 42% mostró longitudes promedio de hembras significativamente mayores que la de los machos y el 58% restante reveló lo contrario. La familia 6 tam- bién mostró un dimorfismo significativo alto (18/20 = 0,90) pero al contrario de la familia 11, las longitudes promedio de hembras fueron significativamente mayores que la de los machos (11/18 = 0,61) mientras que lo contrario de observó en el 39% restante. En general, existe dimorfismo sexual en los promedios de la longitud, el alargamiento, los tamaños de las manchas costales y en la tasa promedio PB/ta-III. En todos los casos, se presentó variación en el dimorfismo se- xual asociada con la procedencia (familias) de las progenies. En el 29,2% de los casos con dimorfismo sexual se favorece la mayor longitud de las manchas oscuras en hembras mien- tras que en el 70,8 % se favorece la mayor longitud de las manchas claras en machos.
En la Tabla 4 se registraron los componentes de la varia- ción fenotípica (%) aportados por los factores temperatura, familias y sexos con sus interacciones. Esta tabla muestra un efecto altamente significativo de la temperatura de crianza sobre la variación de la longitud, el alargamiento (tasa longi- tud/ancho) y la fusión de las manchas costales SP + PRSD + ASP del ala derecha, con una variación explicada alta, entre 69,4 % y 98,0 %; a las proporciones PB/ta-III, HP/PHD y SCP/DSD y DSD, con una variación explicada entre 8,4 % y 25,5%, a las machas HP, HD, PHD, PRSD, PP y AD con aportes a la variación fenotípica bajos, entre 2,4% y 6,3%. No mostró un efecto significativo sobre la variación fenotí- pica de las manchas costales BP + PHP, PSP, PSD, SP, ASP, SCP, PD y AP.
En esta tabla también se observó un efecto altamente sig- nificativo de la familia sobre la variación fenotípica de las proporciones HP/PHD, SCP/DSD y PB/ta-III, de los tama- ños de las manchas PHD, HD, PSP, PSD, DSD, SCP, AD y AP con una variación fenotípica explicada alta, entre 21,7% y 45,3 %; de las manchas HP, SP, PRSD, y la fusión SP + PRSD + ASP, con aportes a la variación fenotípica entre 18,8 % y 16,0%. Además, mostró efectos significativos (p < 0,05) pero aportes a la variación fenotípica bajos para la longitud (2,9%) y la tasa largo/ancho (1,6%).
El efecto del sexo sobre la variación fenotípica de los ca- racteres estudiados fue significativo para las tasas largo/ancho (0,1%) y SCP/DSD (8,0%) y los tamaños de las manchas PSD (7,1%), DSD (3,1 %), SCP (10,5 %), PD (5,7 %) y PP (4,4%). No mostró efecto significativo sobre la variación fe- notípica de las manchas costales BP + PHP, PD y PP.
No se observaron interacciones significativas entre los tres factores analizados y los valores registrados en la Tabla 4 fue- ron menores al 8,2% (Familia*Sexo) para la mancha PP.
Discusión
Pocas veces se presentó fusión de algunas de las manchas cos- tales o por lo menos ausencia de la accesoria clara (ASP), lo cual corresponde a lo comúnmente observado en especímenes criados en su ambiente natural aunque, según Faran (1980), Nys. triannulatus s.l. presenta frecuentemente la vena C casi completamente oscura y las manchas claras ocasionalmente poco diferenciadas. Pero esto no parece estar de acuerdo con lo descrito formalmente para las especies de este complejo, dado que las manchas claras, a pesar de su tamaño pequeño, casi siempre están presentes. En estudios para esta misma es- pecie, se ha observado, aunque en baja frecuencia, de tres a cuatro tipos de patrones costales adicionales, pero restringi- dos a progenies de familias obtenidas de muestras de pobla- ción del departamento del Meta (Marín Londoño 2014).
Por otro lado, en la descripción original de Nys. halophylus, se indicó que ASP no está presente en el 30 % de los ejemplares examinados, es decir que el patrón costal I podría estar cercano al 70%, lo cual parece ser también fre- cuente en algunas poblaciones de Nys. triannulatus s.s. (Sil- va do Nascimento y Lourenço-de-Oliveira 2002). En algunas familias de muestras de poblaciones (datos no publicados) de los departamentos colombianos de Huila, Meta, Putumayo y Valle del Cauca, la frecuencia de este patrón varió entre 33,5 y 100%, es decir que en esta especie este carácter es altamente polimórfico y parece contribuir a la plasticidad de los carac- teres fenotípicos. El patrón costal II se puede confundir con el III (como se describe en el presente escrito), debido a que ambos involucran a la mancha costal ASP. Sin embargo, en el patrón II se expresa un tamaño-mayor de la mancha clara que involucra la SP, PRSD y ASP y por lo tanto, la SP es obser- vada con mayor tamaño que en el patrón I, mientras que en el patrón III, hay expresión de la ASP, es decir que queda incor- porada a la SD y por lo tanto, esta mancha oscura se observará más grande que en el patrón I.
Para mayor claridad, sobre los patrones costales II y III, conviene resaltar que para las especies del subgénero Nyssorhynchus, Wilkerson y Peyton (1990) definen al sector claro (SP) como un grupo de escamas claras presentes sobre la costa, subcostal y/o radio, que coinciden con la posición de la vena transversal sc-r, mientras que para la posición de la mancha costal ASP coincide la posición de la vena transversal r1 -rs.
La temperatura en la cual ocurrió el desarrollo de las for- mas inmaduras, como en todos los organismos ectotermos, no solo influye de manera marcada en su velocidad de desarrollo preimagal (datos no evaluados) sino que también parece ser uno de los factores que contribuye a la variación en la expre- sión de patrones de manchas costales (Fig. 1). La relación en- tre las frecuencias de los patrones costales y el incremento de la temperatura (20, 24 y 28 °C) mostró asociación significati- va en hembras (χ2 = 7,4; p = 0,0065) mientras que en machos no fue evidente (χ2 = 0,03; p = 0,8607). De acuerdo con esto, el incremento en la temperatura parece favorecer la aparición de fusiones de manchas costales en hembras, mientras que en machos el patrón de manchas parece ser más estable.
La longitud del ala es una característica altamente corre- lacionada con el tamaño del insecto, es decir que, a mayor ta- maño de alas mayor tamaño corporal, por eso es considerado para especies de Culicidae como un estimador indirecto del tamaño corporal (Dujardin 2011). En las descripciones de las especies del complejo se ha observado que esta especie pre- senta un tamaño relativamente pequeño en comparación con otras especies del subgénero Nyssorhynchus (Faran 1980). En el presente estudio, la longitud promedio de las alas dere- chas en Nys. triannulatus fue significativamente menor que las descritas por Silva do Nascimento y Lourenço-de-Oliveira (2002) para Nys. triannulatus y similares a los de Nys. halo- phylus. Estos autores observaron diferencias morfométricas significativas entre Nys. halophylus y Nys. triannulatus (p < 0,05). Sin embargo, también hay que considerar que las con- diciones ambientales de los criaderos (temperatura, densidad, físico química del agua y cantidad y calidad del alimento) in- fluyen en el tamaño larval y por ende en el tamaño de pupas y adulto (Lanciani y Le 1995). Esto es una posible explicación para entender por qué las progenies obtenidas en laboratorio suelen ser más pequeñas que las criadas en su ambiente na- tural.
Por otro lado, independiente de las características genéti- cas de cada especie y de la temperatura en la cual ocurre su de- sarrollo larval, se ha observado que la alimentación larval en especies de Anopheles afecta no solo el tamaño sino también la velocidad de su desarrollo, la supervivencia y caracteres biológicos de los adultos (Araújo et al. 2012), esto incluye, desde luego, su tamaño corporal y en general “fitness” de cada población o especie. Con base en los resultados del presente estudio (Tabla 2), la longitud del ala es una característica alta- mente variable entre las familias, aun para progenies criadas a la misma temperatura. Las diferencias en tamaño de alas, encontradas entre progenies obtenidas en temperaturas de 20 y 24 °C, pueden explicar en parte las diferencias observadas entre las poblaciones de algunas especies de Anopheles (Ro- dríguez-Zabala et al. 2016; Pacheco et al. 2017; Pacheco-Gó- mez et al. 2018).
Es decir que en condiciones naturales es también posible que la longitud pueda ser un marcador morfológico de pobla- ciones y debido a que el tamaño corporal puede influenciar la capacidad vectorial de especies de Anopheles, es importan- te incluir la longitud alar como un estimador indirecto de su importancia epidemiológica para la malaria (verificable con datos de incriminación), taxonomía y asociación con aspectos propios de su bionomía (Pacheco-Gómez et al. 2018).
En anofelinos se ha observado que la reserva de ener- gía que requiere una hembra para la búsqueda de su fuente de alimentación con sangre depende de su tamaño corporal (Dudley 2002). Un mayor tamaño puede estar relaciona- do con un mayor vigor y esto con una mayor longevidad. Esto es de gran importancia epidemiológica porque se re- laciona con una mayor competencia vectorial para la trans- misión del agente etiológico de la malaria, ya que reali- zan mayor número de ingestas de sangre, necesarias para el desarrollo del ciclo gonotrófico de cada hembra, y con esto se incrementa la probabilidad de adquirir y trans- mitir los parásitos de la malaria humana. Por otro lado, la forma y longitud de las alas constituyen característi- cas que contribuyen a definir la eficacia del vuelo (Dud- ley 2002), rasgo fundamental para la búsqueda de sitios aptos para la oviposición, fuentes de sangre (Charlwood 1996), la dispersión de genes de resistencia a insectici- das (Norberg 1995) y la propagación de esporozoitos de Plasmodium spp. (Dudley 2002). Estos rasgos del ala no dependen solo del acervo genético (Fusco y Minelli 2010) sino que son afectados por variables ambientales, como la temperatura, la humedad relativa y la altitud (Gómez et al. 2013; Gómez et al. 2014; Lorenz et al. 2014).
La variación en la longitud del ala derecha fue muy sus- ceptible a cambios de temperatura (Tabla 7). Si se asume que las familias de hermanos completos estiman el componente genético de la generación parental, la H2 estimada en sentido amplio fue baja (2 * 2,9 % = 5,8 %). Esto significa que este carácter tiene un componente genético proporcionalmente bajo, posiblemente atribuible a que los individuos dentro de las familias fueron muy variables y por ende el residual se hace proporcionalmente mayor respecto al componente entre familias de individuos criados a la misma temperatura. Estos resultados coinciden con los encontrados por Gómez y Co- rrea (2017) quienes a partir del análisis morfométrico de An. albimanus concluyen que el tamaño del ala es variable a nivel intra e interespecífico puesto que cada subgénero presenta una tendencia de tamaño de ala particular, posiblemente relacio- nada con la historia evolutiva de estos linajes.
Los resultados indicaron que las alas de esta especie tien- den a ser estrechas y por ende presentan menor resistencia inducida (Cerón-Muñoz et al. 2013). Por otro lado, sugieren que las de los machos son significativamente menos anchas que las de las hembras. Desde el punto de vista adaptativo, un ala más ancha tiene más resistencia inducida y menos resis- tencia parasita que un ala alargada y más estrecha como la de los machos. Esto significa que las alas en hembras son más re- sistentes. Posiblemente esto esté asociado con el mayor tama- ño o peso de las hembras mientras que la resistencia parasita es menor en machos, lo cual favorece una mayor dispersión al aumentar la velocidad de desplazamiento activo (Bomphrey et al. 2017).
Las dos variables, longitud y tasa largo/ancho mostraron un aporte muy alto del efecto temperatura. Sin embargo, este componente podría estar sobreestimado por el ambiente co- mún dentro de cada uno de los tres niveles de temperatura (20, 24 y 28 °C) generando más parecido entre las familias del mismo nivel por el efecto de condiciones adicionales como la cantidad y calidad de la alimentación, aspectos fisicoquímicos del agua y densidad de larvas, pero dado que estas variables no fueron medidas experimentalmente, fue imposible cuanti- ficar el efecto de esta fuente de variación (Falconer y Mackay 1996). Estas mismas razones, podrían hacer que el efecto de la familia se reduzca y, por lo tanto, se subestime la H2 en sentido amplio.
Los machos y las hembras de una misma especie a me- nudo difieren en los rasgos relacionados con la morfología, la fisiología y el comportamiento, y se cree que tales dispari- dades surgen de la sincronización de la asignación de recur- sos, especialmente con respecto a la reproducción (Badyaev 2002). Frecuentemente, los machos buscarán y competirán por las hembras, mientras que las hembras elegirán entre los machos competidores y generarán la descendencia (Petrie et al. 1991; Miller y Svensson 2014). Tales antecedentes gene- rales conducen a inversiones desproporcionadas en rasgos entre los sexos, con las hembras invirtiendo relativamente más en su aparato productivo y en la cría en la mayoría de los casos (Thornhill y Alcock 1983; Olof et al. 1994; Mi- ller y Svensson 2014). Esto podría explicar el dimorfismo sexual encontrado para los promedios de la longitud, el alar- gamiento, los tamaños de las manchas costales y en la tasa promedio PB/ta-III, (Anexo 2). En el 29,2 % de los casos con dimorfismo sexual se favorece la mayor longitud de las manchas oscuras en hembras mientras que en el restante por- centaje se favorece la mayor longitud de las manchas claras en machos.
El efecto de la temperatura para la tasa SCP/DSD podría explicarse bajo el supuesto de considerar que el fenotipo más estable se da a 24 °C, en donde el tamaño de estas dos man- chas es antagónico (mayor longitud de DSD menor longitud SCP o viceversa). Esta relación es muy sensible a la tempera- tura como lo propusieron Sueur et al. (1992). A 20 o 28 °C se estabiliza mostrando en algunas familias relaciones fenotípi- cas directas significativas y en otras no.
Los aportes más altos del factor familia para PHD, HD, y las proporciones HP/PHD, SCP/DSD (42,5 %) y PB/ta-III, podrían indicar que el aporte genético sobre la variación feno- típica es significativo, independientemente de la temperatura donde se criaron las progenies. Por otro lado, el hecho de que se presente significancia en este componente entre tasas tam- bién podría indicar covariación genética significativa en estas características, lo cual podría explicarse por acción pleiotró- pica o epítasis de genes involucrados en la pareja de caracte- res. Esto último, también podría explicar porque la temperatu- ra muestra un mayor efecto sobre la fusión SP + PRSD + ASP (69,4%) que, sobre cada una de las manchas individuales, sobre las cuales el efecto no fue significativo (SP y ASP) y sólo aportó el 2,8% a la variación fenotípica de PRSD.
Conclusiones
Independientemente de la temperatura de crianza de las lar- vas, las progenies de familias de Nys. triannulatus del sur de Colombia obtenidas en cámaras ambientales reguladas entre 20 y 28 °C, muestran alta frecuencia de solamente uno de los patrones costales (I) y dimorfismo sexual para la tasa largo/ ancho de las alas, la cual es significativamente mayor en ma- chos.
En cada una de las temperaturas de crianza se presenta dimorfismo sexual en muchos de los otros caracteres analiza- dos. Dependiendo del sexo, temperaturas más altas favorecen el aumento de la longitud de las manchas claras u oscuras. En hembras, en casi la tercera parte de los casos, se favorece la mayor longitud de las manchas oscuras, mientras que, en machos, aproximadamente las dos terceras partes de los ca- sos, presentan mayor longitud de las manchas claras.
En general, en las familias, la temperatura influye sobre los tamaños de las manchas costales y tasas que relacionan algunas manchas vecinas, pero el efecto de los sexos sobre la variación fenotípica de los caracteres es significativo so- lamente para algunos de estos, presentándose pocas inte- racciones significativas entre temperatura, familia y sexo. Sin embargo, es importante resaltar, que la longitud de las alas también varía significativamente entre progenies de una misma temperatura y entre familias de individuos. Por otro lado, el aporte genético al tamaño, así como de las familias, a la variación fenotípica, es significativo en casi todos los ca- racteres que fueron analizados en las alas de N. triannulatus.
Origen y financiación
Este manuscrito fue originado en el proyecto de investigación (No. 71106), titulado “Efecto de la temperatura en la expresión fenotípica del área costal de Anopheles (Nyssorhynchus) triannulatus (Diptera: Culicidae: Anophelinae), usando cámaras ambientadas diseñadas en la Universidad del Valle”. Este fue financiado por la Universidad del Valle, presupuesto asignado mediante convocatoria interna.
Contribución de los autores
Ranulfo González Obando: Verificación taxonómica, fotografías, realización de mediciones, colaboración en la interpretación de los análisis y escritura del manuscrito. Coordinación de las actividades de revisión del manuscrito.
Heiber Cárdenas Henao: Realización de mediciones, análisis, interpretación y escritura del manuscrito.
Omar Alejandro Marín-Londoño. Recolección, cría y manejo del material biológico, preparación de placas, fotografías, ayuda en el análisis de los datos y escritura del manuscrito. Todos los autores aprobaron la versión final del manuscrito.
Footnotes
Agradecimientos
Agradecemos a la Universidad del Valle y a su Vicerrectoría de Investigaciones por la financiación concedida a los autores a través del proyecto No. 71106, y a la Vicedecanatura de In- vestigaciones de la Facultad de Ciencias Naturales y Exactas por su apoyo en los trámites de ejecución. A Jefferson Pan- che por su invaluable ayuda en el trabajo de recolección de ejemplares, así como a las personas de Puerto Asís quienes colaboraron en el proceso de obtención de las muestras de Anopheles. A Nelson Toro y la sección de Genética del Depto. de Biología de la Universidad del Valle. HCH y RGO, agra- decen al Departamento de Biología, Facultad de Ciencias Na- turales y Exactas, y Vicerrectoría de Investigaciones, Univer- sidad del Valle, Santiago de Cali, Colombia, por su continuo soporte y apoyo.
Anexo 1
Números de hembras (n1) y machos (n2), promedios y desviaciones estándar de la longitud (mm), de los tamaños de manchas costales (*), de las proporciones HP/PHD, SCP/DSD y (PB/ta-III2) y de la fusión SP + PRSD + ASP, en las alas derechas de las progenies hembras y machos de Nys. triannulatus, criadas a tres temperaturas.
Anexo 2.
Pruebas t-student con su significancia (p) para comparar longitud promedio y tamaños promedio de las manchas costales-ala derecha (*) y tasa promedio de la mancha oscura basal/longitud del tarsómero posterior dos (PB/ta-III2) de progenies hembras (H) y machos (M) en cada familia de Nys. triannulatus. Temperaturas: (a) 20 °C; (b) 24 °C; (c) 28 °C.