
Abstract
Shade grown coffee crops generate environments more similar to mountain ecosystems than monocultures of coffee exposed to direct sunlight. Although butterflies are considered useful bioindicators of the state of ecosystem health, their diversity remains unknown in coffee agroecosystems in Santander, where coffee is grown under shade. The structure and composition of the assembly of the Papilionoidea in a shade coffee plantation (Hacienda La Capellanía, Curití, Santander) at an altitude of 1756 m (August-September, 2014) are described. In three parallel transects of 50 m inside the crop, nine Van Someren-Rydon traps were baited with decomposed fruit, and additional specimens were captured with an entomological net. The sampling coverage was estimated at 81.67% (Clench equation). Overall, 341 individuals of 49 species of the family Nymphalidae (spp. = 39), Pieridae (spp. = 7), Papilionidae (spp. = 2) and Lycaenidae (sp. = 1) were captured. The most abundant species were Cissia pompilia, Graphita griphe and Hermeuptychia harmonia. Species richness (α = 16), dominance (D = 0.07), evenness (J' = 0.81) and effective number of species (°D = 58.8, ¹D = 23.73 and βD = 14.26) were estimated. Vegetation structure was related to the composition of butterfly communities (canonical correspondence analysis). The abundance of butterflies was significantly associated with the percentage of tree cover, height, trunk diameter, and the abundance of trees in the coffee plantation. This study is one of the first to describe the diversity of butterflies associated with shade coffee in the department of Santander. Traditional shade coffee agroecosystems may benefit the diversity of butterflies because the heterogeneity of their plant structure offers a greater availability of resources and shelter for them.
Introducción
En Latinoamérica se tienen cerca de cinco millones de hectáreas de cultivos de café (FAO 2015) representando éste una de las actividades agropecuarias más importantes para países como México, Brasil y Colombia (Perfecto et al. 2005). Desde el siglo XIX, Colombia cuenta con una actividad cafetera como principal producto de exportación y es uno de los principales cultivos esenciales en el desarrollo económico del país (Lozano y Yoshida 2008) alcanzando entre 772.000 y 949.000 ha sembradas de café en 22 departamentos (Federación Nacional de Cafeteros de Colombia 2009; FAO 2015).
Moguel y Toledo (1999) clasificaron los sistemas de producción de café según su complejidad, la estructura de la vegetación y cómo se va reduciendo gradualmente la cobertura de sombra. En los cultivos rústico tradicional y policultivo tradicional se conservan los árboles nativos para generar la sombra; en los policultivos comerciales los árboles nativos se remueven y son sustituidos por árboles de interés comercial; en los monocultivos con sombrío los árboles son reemplazados por una o dos especies de árboles leguminosos para proveerle sombra al café; finalmente, en los monocultivos sin sombrío se remueve la totalidad del dosel exponiendo al sol las plantas de café. Los últimos tres sistemas involucran el uso de agroquímicos (Moguel y Toledo 1999).
Como resultado de las características climáticas y altitudinales de la región norte colombiana, en los departamentos de Casanare, Santander y Norte de Santander se produce únicamente café bajo sombra (Federación Nacional de Cafeteros de Colombia 2010). Perfecto et al. (2005) y Koh y Gardner (2010) propusieron que los sistemas productivos como los cafetales con sombrío pueden generar beneficios a la biodiversidad y a la conservación de especies a diferencia de otros cultivos tecnificados.
Se han realizado diversos trabajos que describen la diversidad de fauna asociada a los sistemas de cafetales con sombrío en Colombia (p.e. Rivera y Armbrecht 2005; Ortegón-Martínez y Pérez-Torres 2007; Pérez-Torres et al. 2009). En el caso de las mariposas, se ha estudiado la diversidad y composición asociadas a este tipo de sistemas productivos (Muriel & Kattan 2009; Millán et al. 2009; Muriel et al. 2011), principalmente en la región cafetera del departamento de Antioquia y poco se conoce sobre la diversidad de éstas en los cafetales con sombrío del departamento de Santander.
En este trabajo se emplearon mariposas adultas como modelo de estudio debido a que: son consideradas como organismos indicadores de diversidad (Brown y Freitas 2000; Van Swaay et al. 2015); por la relación directa que presentan con sus plantas hospedantes (Wahlberg 2001; Bonebrake et al. 2010); a su respuesta ante los cambios y perturbaciones ambientales (Brown 1997); a su papel como proveedores de servicios ecosistémicos como la polinización (Gilbert 1972; Tobar et al. 2001); y a la facilidad de colectar, manipular e identificar en campo (Brown y Freitas 2000).
Este estudio tuvo como principal objetivo, describir la riqueza y diversidad del ensamblaje de mariposas diurnas (Lepidoptera: Papilionoidea) y su relación con la estructura del paisaje en un agroecosistema de café con sombrío en el municipio de Curití (Santander, Colombia). Dada la escasez de información de la biodiversidad asociada a sistemas productivos en el departamento de Santander, y teniendo en cuenta que el café en este departamento principalmente se siembra bajo sombra, es importante conocer qué tanto los cafetales con sombrío contribuyen a la diversidad de mariposas en esta región.
Materiales y métodos
Área de estudio.
El estudio se desarrolló en un cafetal de sombra con un área cercana a las 10 ha, ubicado en la vereda Macaregua-San Francisco dentro del municipio de Curití (06°39'01,8"N y 73°06′07,7"O) en la región del Chicamocha del departamento de Santander (Colombia) (Fig. 1).
La región presenta una temporada seca (noviembre a marzo) y una húmeda (abril a octubre), con precipitaciones que varían entre los 1.100 mm anuales en la parte nororiental sobre el río Chicamocha y los 2.000 mm al sur de la región (Mejía 2008). Dada la variación y la heterogeneidad del relieve la temperatura oscila entre los 12 °C y los 30 °C presentándose una humedad relativa del 60 al 80% a una elevación promedio de 1.700 msnm (Castañeda 2012).
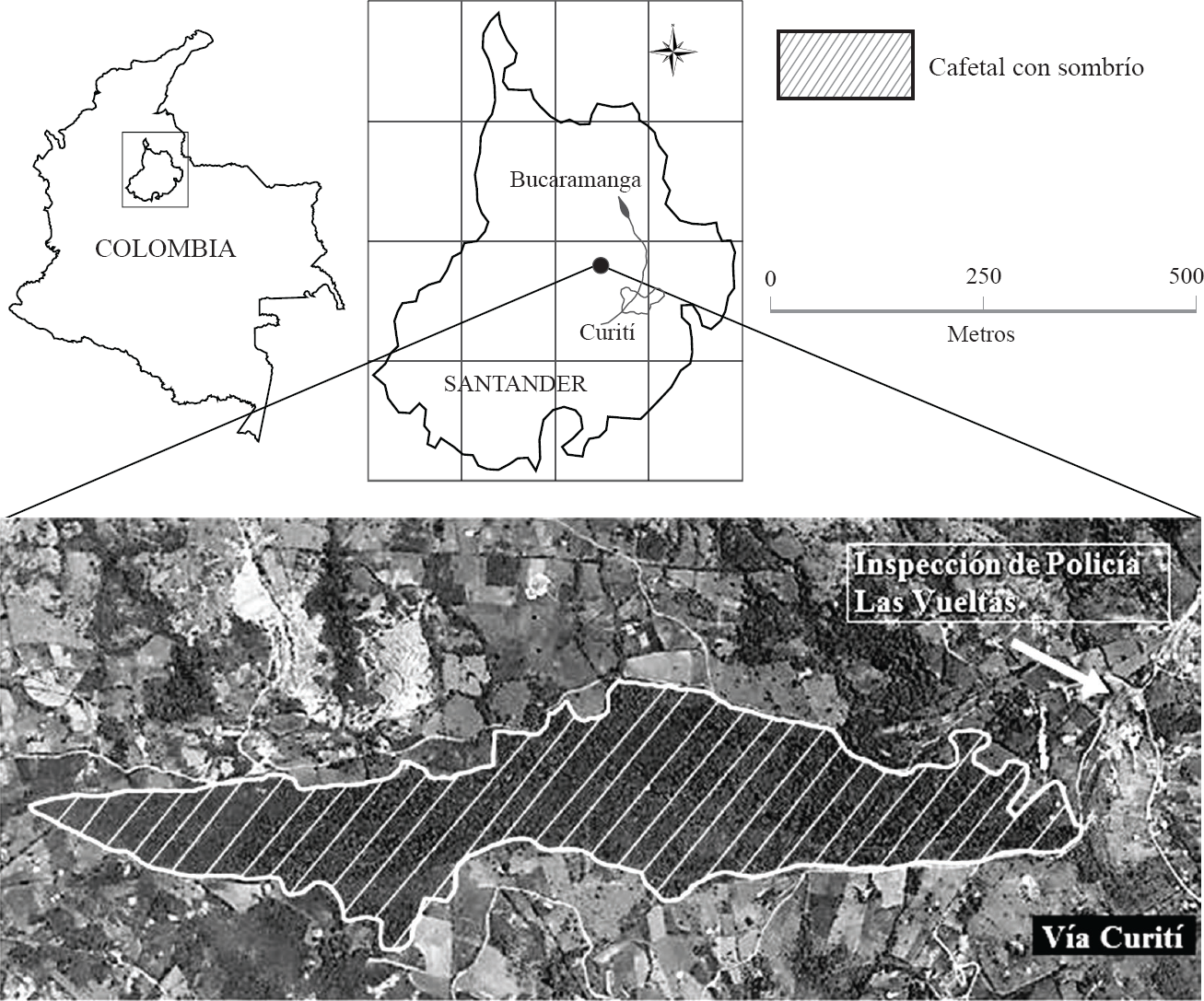
Ubicación del cafetal con sombrío y su posición respecto a la cabecera municipal de Curití (Santander, Colombia).
En el municipio de Curití se encuentran Bosques secos Tropicales, Bosques secos Premontanos, Bosques húmedos Premontanos y Bosques húmedos Montano Bajos (Mejía 2008). Muchas de estas coberturas se han transformado en zonas destinadas a la ganadería, los sistemas agroforestales, los cultivos perennes y semipermanentes de café, tabaco, fríjol y maíz; además existen relictos de bosques ribereños y rastrojos (Mejía 2008). De acuerdo a Rodríguez et al. 2006, la zona se encuentra en el zonobioma alterno hígrico y/o subxerofítico tropical (ZAST) cañón del río Chicamocha y al orobioma subandino cordillera Oriental. En la plantación se siembra café orgánico desde hace aproximadamente 40 años y no se emplean agroquímicos ni fertilizantes.
Muestreo.
El trabajo de campo se realizó entre los meses de agosto y septiembre de 2014, correspondientes a la temporada de lluvias de la zona. Se instalaron nueve trampas tipo Van Someren-Rydon (Freitas et al. 2014) activadas del 20 al 25 de agosto y del primero al cinco de septiembre.
Se hizo una modificación del transecto lineal propuesto por Pollard y Yates (1993) para la recolección de mariposas: se establecieron tres transectos paralelos a 50 m de distancia entre sí. En cada transecto se instaló una trampa tipo Van Sommeren-Rydon a los 0, 25 y 50 m. No fue posible colocar las trampas a mayor distancia debido al relieve escarpado y a la dificultad del terreno y la cobertura. Las trampas fueron cebadas con fruta en descomposición (banano fermentado, miel de caña de azúcar y cerveza) para atraer mariposas (Villareal et al. 2006; Freitas et al. 2014; Marín et al. 2014) y el cebo fue remplazado cada dos días o en caso de encontrase seco. Las trampas fueron colocadas entre uno a 1,50 m de distancia del suelo (DeVries 1988) y estuvieron activas las 24 horas.
Se obtuvo un total de 80 horas de muestreo efectivo empleando las redes entomológicas y revisando las trampas Van Someren-Rydon desde las 8:00h hasta las 16:00h durante los 10 días de muestreo. Este muestreo lo realizó una persona.
Los ejemplares recolectados fueron sacrificados mediante presión digital del tórax (Villareal et al. 2006) y guardados de manera independiente en sobres de papel milano con el registro de sus respectivos datos de campo. Las recolectas se realizaron bajo el "permiso marco de recolección de especímenes de especies silvestres de la Diversidad Biológica con fines de investigación científica no comercial" según la resolución 0546 de 29 de mayo de 2014 otorgado a la Pontificia Universidad Javeriana.
Los especímenes fueron examinados y procesados en el Laboratorio de Ecología Funcional de la Pontificia Universidad Javeriana de Bogotá, en donde fueron montados y, posteriormente determinados a nivel de especie y subespecie a través de las claves ilustradas y revisiones taxonómicas de Le Crom et al. (2002; 2004). Además fueron comparados con la base de datos Butterflies of America (Warren et al. 2016). Los ejemplares de difícil identificación fueron consultados con expertos en los diferentes grupos de mariposas de la región neotropical. Finalmente fueron depositados en la Colección de Entomología (MPUJ-ENTO) del Museo Javeriano de Historia Natural de la Pontificia Universidad Javeriana de Bogotá.
Caracterización vegetal.
Debido a que las mariposas responden a las características estructurales de la vegetación (Ribeiro et al. 2012), esta se caracterizó en el cafetal con sombrío. Para cada trampa Van Someren-Rydon se demarcó una parcela de 5 x 5 m y se registró la altura total, el diámetro a la altura del pecho (DAP), el porcentaje de cobertura y el número de árboles.
Análisis de datos.
La composición de la comunidad de especies de mariposas se analizó mediante el índice alfa (a) de Fisher (Moreno 2001) empleando el software PAST 2.17 (Hammer et al. 2001). Se realizó una curva de acumulación de especies considerando el número de especies en función del número de individuos totales. Para eliminar la influencia del orden de adición de las muestras, la muestra fue aleatorizada 1.000 veces mediante el uso del programa EstimateS 9.1.0. (Colwell 2013).
Con el objeto de calcular el número de especies esperadas, la curva fue ajustada al modelo de Clench mediante una regresión no lineal utilizando el programa Statistica 12 (StatSoft 2013). El modelo tiene en cuenta la ganancia de experiencia en el método de muestreo en campo, por lo que la probabilidad de obtener un mayor número de especies nuevas añadidas al inventario se incrementa a medida que se tiene una mayor experiencia en el método de muestreo (Moreno 2001; Jiménez-Valverde y Hortal 2003). La ecuación de Clench es: E(S) = (ax) / (1 + bx). Donde: a = la ordenada del origen, la intercepción en Y representa la tasa de incremento de la lista al inicio de la colección; b= pendiente de la curva; x= número de especies esperadas en xáµ¢ muestras. Este modelo se considera asintótico debido a que predice la riqueza específica como el número de especies al cual la curva de acumulación de especies alcanza la asíntota, la cual es calculada como la relación a/b (Soberón y Llorente 1993; Moreno 2001).
Se calcularon los valores de dominancia (índice de Simpson) y de equidad (a partir del índice de Shannon) (Moreno 2001). La diversidad se expresó en términos de los números efectivos de especies (Jost 2006). En éstos últimos, el exponente q determina la sensibilidad del índice de las abundancias relativas de las especies e indica cuántas especies son consideradas en la muestra analizada dependiendo de su nivel de rareza (Moreno et al. 2011). La diversidad de orden q⁰ (⁰D) es insensible a las abundancias de las especies, por lo que corresponde a la riqueza de especies; q¹ (¹D) incluye a todas las especies con un peso proporcional a su abundancia en la comunidad, es decir, equidad; el exponente con valores mayores a 1 (q>1) tiene sensibilidad a las especies comunes y corresponde a la dominancia (Moreno et al. 2011). Estos análisis fueron realizados mediante el programa SPADE (Chao y Shen 2010) para obtener los respectivos intervalos de confianza del 95%.
Con el fin de evaluar la relación de la estructura vegetal con la riqueza de especies de mariposas encontrada en el sitio de muestreo, se realizó un análisis de correspondencia canónica (ACC). Se seleccionaron las especies que fueron capturadas con red entomológica y las trampas Van Someren -Rydon dentro de la parcela establecida de 5 x 5 m alrededor de cada trampa. Los cálculos se realizaron con el programa PAST 3.02 (Hammer et al. 2001).
El ACC es un método que toma el promedio ponderado a un análisis simultáneo de varias especies y variables ambientales (Braak y Verdonschot 1995). Debido a que el ACC emplea la estadística de regresión lineal, ayuda a estimar y evaluar estadísticamente los efectos de las variables ambientales en las comunidades biológicas. De esta manera se puede explicar cómo las especies responden a condiciones ecológicas como las variables ambientales, e identifica cuál de las variables es más importante en determinar la composición de una comunidad (Braak y Verdonschot 1995).
Resultados
Estructura y composición del ensamblaje de mariposas.
Se colectó un total de 341 individuos pertenecientes a 49 especies de cuatro familias de Papilionoidea: Lycaenidae (2 %), Nymphalidae (79,6 %), Papilionidae (4,1 %) y Pieridae (14,3%) (Tabla 1, Fig. 2). La subfamilia más dominante fue Satyrinae con 15 especies (30,61 %) dentro de ésta, la especie dominante fue Cissia pompilia (n = 66), seguida de Graphita griphe (n = 39) y Hermeuptychia harmonia (n = 25). A esta subfamilia le siguieron las subfamilias Danainae y Heliconinae, cada una con 12,24 %. De la familia Pieridae, la subfamilia Coliadinae fue la que presentó la mayor cantidad de especies (8,16%) (Fig. 2).
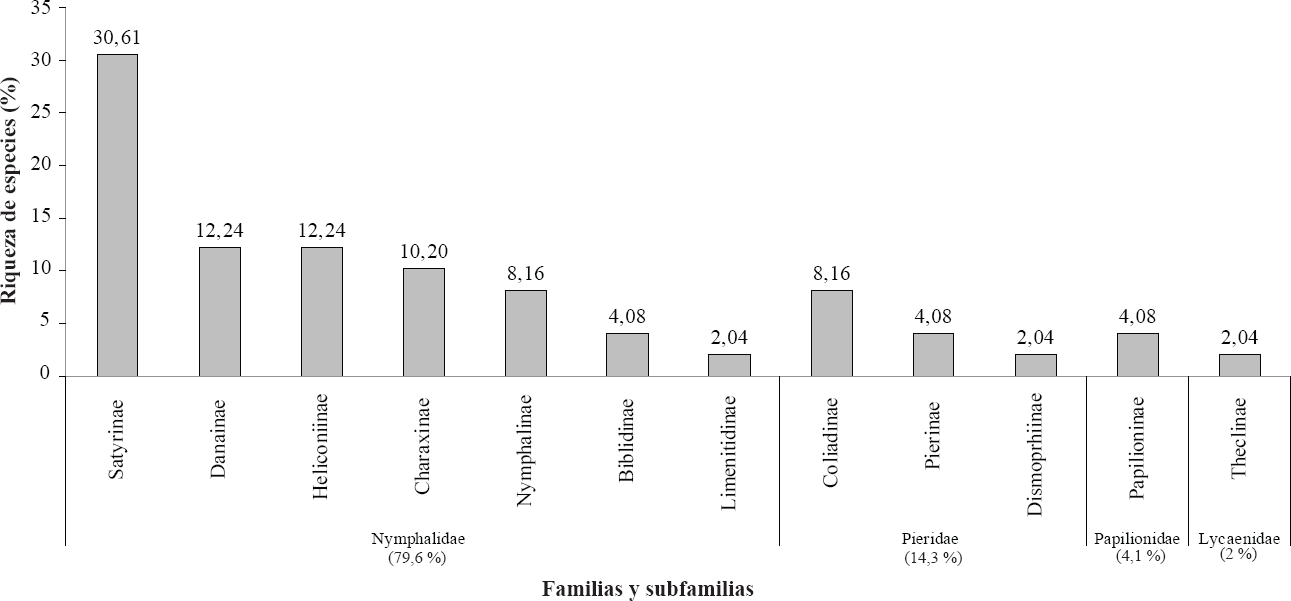
Riqueza y abundancia de las familias y subfamilias de mariposas encontradas en el agroecosistema de café bajo sombra.
De acuerdo con el ajuste al modelo de Clench de la curva de acumulación de especies (Fig. 3) representada como el número de especies en función del número de individuos totales, las 49 especies observadas representaron el 81,67 % de las especies esperadas (S = 60).
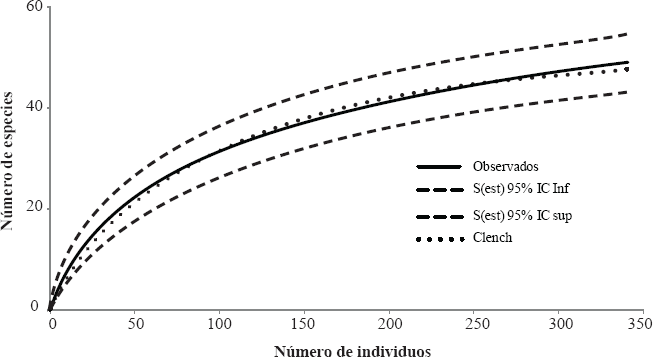
Curva de acumulación de especies representada como el número de especies en función del número de individuos totales. Se presenta el ajuste de la curva por la Ecuación de Clench y sus respectivos intervalos de confianza del 95%.
El valor de riqueza alfa de Fisher fue a = 13,75 < 15,68 < 17,62; la dominancia fue D = 0,058 < 0,073 < 0,087 y la equidad fue J' = 0,785 < 0,814 < 0,843. Se presentan los valores en términos de números efectivos de especies con sus respectivos intervalos de confianza del 95% (Fig. 4). La diversidad de orden ⁰D (que expresa la riqueza esperada de especies) estuvo entre 58,8 (Chao1) y 62,1 (ACE) indica que la representatividad del muestreo estuvo entre el 78,9 % y el 83,3%. Estos valores de representatividad, junto con el valor presentado anteriormente del ajuste al modelo de Clench indican que se cuenta con una representación plausible de las especies presentes en el cafetal con sombrío estudiado. Por otra parte, los valores ¹D = 23,73 y βD = 14,26 sugieren que en el ensamblaje de mariposas del cafetal se presentan algunas especies dominantes (aproximadamente 15) y un alto número (n = 34) de especies raras (menos de seis individuos). Lo anterior se refleja en las abundancias relativas de cada una de las especies y explica que la curva de distribución de abundancias (Fig. 5) se haya ajustado al modelo log-serial (P = 0,9674).
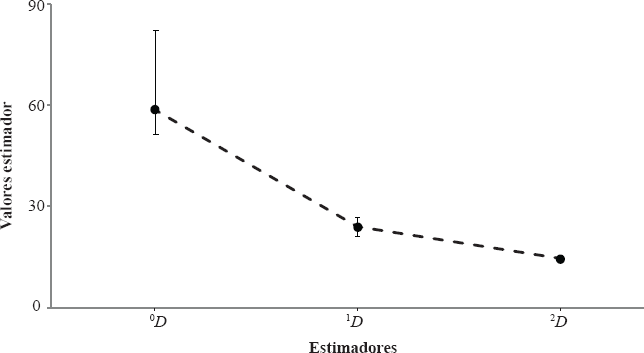
Perfil de diversiad de las mariposas asociadas al agroecosistema de café bajo sombra. ⁰D = Estimador de riqueza de espcies, ¹D = Exponente del índice de Shannon y βD = inverso del índice de Simpson. Las líneas verticales representan los intervalos de confianza del 95%.
Especies de mariposas asociadas a un cultivo de café bajo sombra en el municipio de Curití (Santander, Colombia). n = abundancias.
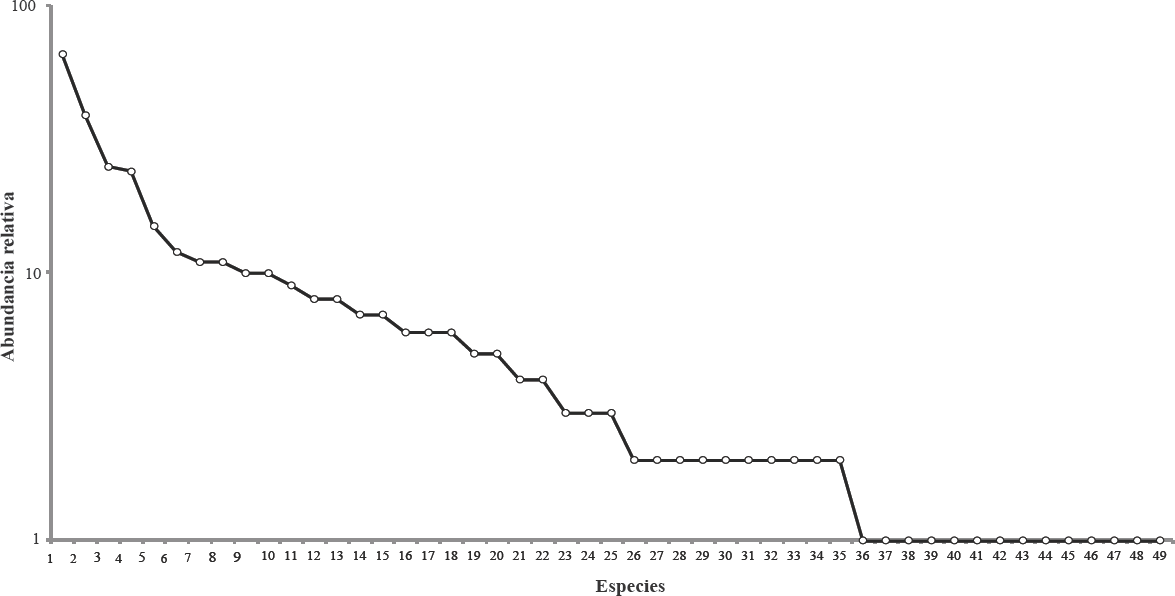
Distribución de abundancias relativas de las especies de mariposas del cafetal con sombrío del municipio de Curití (Santander, Colombia). 1. Cissia pompilia, 2. Graphita griphe, 3. Hermeuptychia harmonia, 4. Heliconius erato hydara, 5. Taygetis virgilia, 6. Yphthimoides renata, 7. Eurema mexicana bogotana, 8. Taygetis laches, 9. Heliconius clysonymus clysonymus, 10. Mechanitis polymnia, 11. Athesis clearista, 12. Dircenna jemina, 13. Hermeuptychia hermes, 14. Leptophobia aripa, 15. Pareuptychia metaleuca, 16. Episcada salvinia apia, 17. Opsiphanes cassina numatius, 18. Phoebis sennae marcellina, 19. Anthanassa drusilla drusilla, 20. Caligo brasiliensis galba, 21. Dryas iulia, 22. Eueides isabella, 23. Eurema arbela gratiosa, 24. Prepona pylene gnorima, 25. Zaretis ellops, 26. Archaeoprepona demophon muson, 27. Ascia monuste, 28. Eunicamonima, 29. Greta andromica, 30. Heliconius ethilla semiflavidus, 31. Magneuptychia tiessa, 32. Memphis lyceus, 33. Oressinoma typhla, 34. Pedaliodes manis, 35. Smyrna blomfildia, 36. Adelpha alala completa, 37. Colobura dirce, 38. Dismorphia crisia foedora, 39. Episcada hymenaea, 40. Eueides aliphera aliphera, 41. Eurema albula marginella, 42. Hamadryas fornax fornacalia, 43. Heraclides thoas nealces, 44. Opsiphanes bogotanus bogotanus, 45. Panthiadespaphlagon, 46. Pronophila unifasciata bogotensis, 47. Pterourus coroebus syndemis, 48. Siderone galanthis thebais, 49. Siproeta stelenes.
Estructura de la cobertura vegetal.
La estructura vegetal del cafetal fue compleja y de acuerdo a Perfecto et al. (2005) se clasifica como policultivo tradicional. El estrato arbustivo estuvo conformado por plantas de café de cuatro años con alturas de hasta 1,70m, presentaba ramas superpuestas que dificultaban el desplazamiento en el lugar de muestreo. El estrato arbóreo presentó árboles de 5,1 m y un DAP promedio de 17,2 cm; el porcentaje de cobertura fue de 68 %.
Relación del ensamblaje de mariposas con la estructura de la vegetación.
El ACC evidenció la separación de las variables medidas para la estructura vegetal: porcentaje de cobertura, altura de los árboles, DAP y densidad (número de árboles). Se encontró un grupo de especies (S = 23) que no se asociaron a dichas variables (Fig. 6). Un total de 21 especies se agruparon y relacionaron con las variables de la estructura vegetal, que en este caso fueron las especies más abundantes. La mayoría de las especies agrupadas alrededor de las variables descriptoras de la vegetación, pertenecieron a la subfamilia Satyrinae. La abundancia de las especies Cissia pompilia y Graphita griphe (Satyrinae) fue explicada por el DAP. Por su parte, la abundancia de Hermeuptychia harmonia (Satyrinae), se explicó por el porcentaje de cobertura y número de árboles de la vegetación.
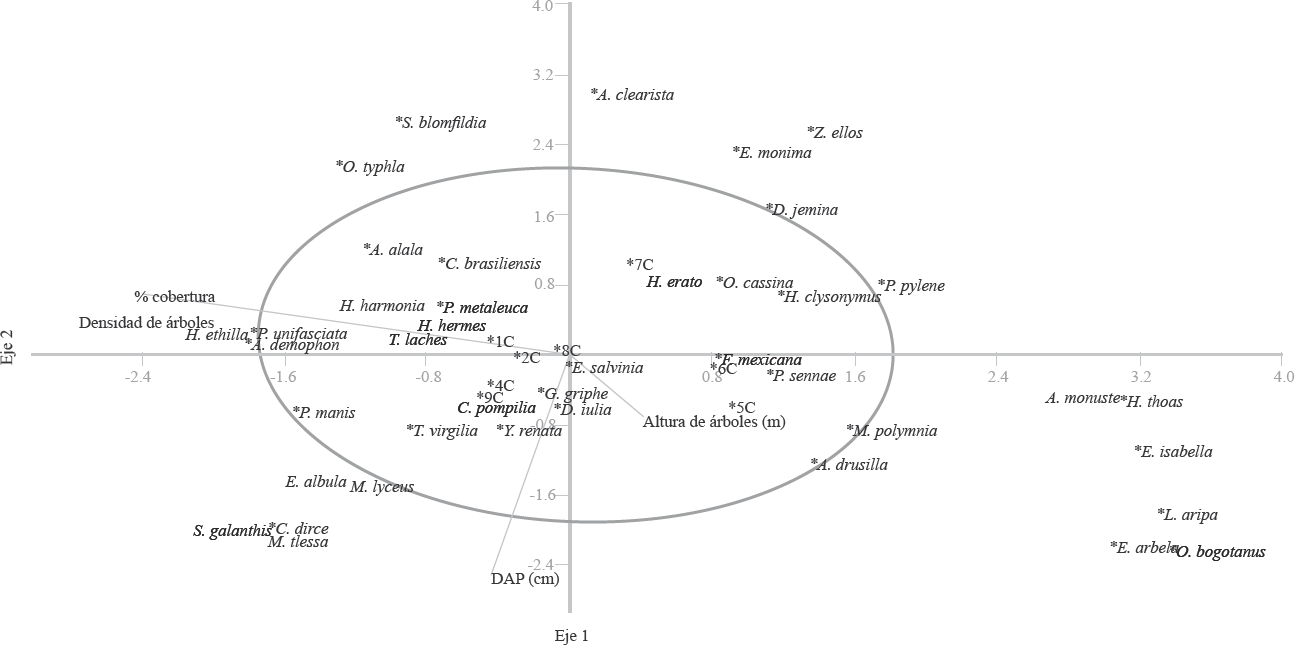
Análisis de correspondencia canónica. En círculo las especies asociadas a las variables de la estructura vegetal (S = 21).
Discusión
En el presente estudio Nymphalidae tuvo el mayor número de individuos y especies. Esto no es sorprendente ya que esta familia presenta una alta riqueza de especies con amplia distribución geográfica y una amplia diversidad de hábitats (DeVries 1987).
El ajuste de la curva de acumulación de especies al modelo de ecuación de Clench (81,67%), permite considerar las muestras obtenidas como una representación plausible del ensamblaje de mariposas presentes en el cafetal con sombrío. La riqueza de especies registradas en este estudio (S = 49) es más alta a la encontrada por Millán et al. (2009), quienes registraron una riqueza de 18 especies en un policultivo de café bajo sombra en el municipio de Caloto (Cauca). Además se encontró un mayor número de especies (S = 39) a la reportada por Muriel y Kattan (2009) para cafetales con sombrío en el departamento de Antioquia. Es posible que esta mayor riqueza se deba a la historia del manejo del cultivo de café en la región del Santander, donde tradicionalmente se ha sembrado café con sombrío. Mientras que en la región de Antioquia y Cauca se tiene principalmente café de sol (Federación Nacional de Cafeteros de Colombia 2010). Mantener históricamente el cultivo de café de sombra a nivel regional pudo favorecer una mayor persistencia de las diferentes especies de mariposas, por el mantenimiento de la cobertura arbórea (más hábitats disponibles) y por favorecer una mayor conectividad entre las diferentes áreas de bosque en la región. Sería importante evaluar este aspecto en un futuro próximo.
Debido a que el cafetal con sombrío de este estudio fue catalogado como rústico o tradicional (Perfecto et al. 2005) y es orgánico, el uso de árboles nativos probablemente provee potenciales plantas hospedantes específicas que requieren las mariposas de esta región para el desarrollo de sus ciclos de vida. Por ejemplo, las Solanaceae, Apocynaceae y Gesneriaceae son abundantes en cafetales de sombra en Colombia y sirven como plantas hospedantes para muchas especies de mariposas (Muriel et al. 2011), así como también Impatiens balsamina (Balsaminaceae), una especie invasora de cafetales que florece brindando recursos a mariposas nectarívoras (Carrero et al. 2013).
El tipo de manejo orgánico en los cultivos de café con sombrío asemejan bosques en estado de conservación que pueden mantener la biodiversidad de mariposas y además contribuyen a evitar la pérdida de especies como resultado de la fragmentación de los bosques (Perfecto et al. 2005; Bhagwat et al. 2008).
La presencia de diferentes especies de árboles y arbustos silvestres en el cafetal favorece la heterogeneidad de la estructura vegetal y, posiblemente, una mayor fuente de recursos como flores y frutos que permite el establecimiento de especies de subfamilias como Danainae, Heliconiinae, Satyrinae y Coliadinae que presentan hábitos nectarívoros y generalistas, además de ofrecer una mayor oferta de espacios como refugio (Carrero et al. 2013; DeVries 1987).
La mayor abundancia de Cissia pompilia, Graphita griphe y Hermeuptychia harmonia (Satyrinae) puede deberse a algunos de sus comportamientos típicos. Estas especies tienden a preferir el sotobosque en busca de frutos en descomposición para su alimentación y, debido a sus patrones de coloración crípticos, buscan protección bajo la sombra (DeVries 1987; Vélez y Salazar 1991).
Aunque en este estudio no se realizó una descripción de la diversidad florística del cafetal, si se observó una gran variedad de flores que potencialmente son recurso para las mariposas de hábito nectarívoro o polinizadoras como por ejemplo, Heliconius erato, Heliconius clysonymus y Heliconius ethilla (Heliconiinae) (DeVries 1987). Estas especies se caracterizan porque pueden cargar polen gracias a las estructuras especializadas que presentan sus probóscides (Gilbert 1972).
De Lycaenidae se registró la menor cantidad de especies, posiblemente debido a un sesgo en el método de captura. Prieto y Dahners (2006) indicaron que las especies de esta familia presentan colores crípticos, tamaño pequeño y baja atracción los cebos de las trampas Van Someren-Rydon. Además, son poco frecuentes de encontrar en estudios e inventarios, en ocasiones debido a falta de experiencia para encontrarlas.
Las especies raras consideradas así por su baja frecuencia en este estudio (n < 18) como por ejemplo Adelpha alala completa, Pronophila unifasciata bogotensis, Episcada hymenaea, Archeoprepona demophon muson, Dismoprhia crisia foedora, Hamadryas fornax fornacalia y Siproeta stelenes también han sido reportadas como raras en estudios en otras zonas de bosques premontanos o subandino conservados del país (Ramírez et al. 2007; Ríos-Málaver 2007; Gaviria-Ortíz y Henao-Bañol 2011). La baja representatividad de estas especies no necesariamente indica que el cafetal presente alto grado de intervención antrópica, sino que son poco comunes o escasas por la época del año en la que se realizaron los muestreos (época húmeda) o poco atraídas al cebo empleado. En este sentido, se recomienda hacer muestreos en diferentes épocas del año (y así poder analizar las variaciones temporales en la abundancia) y usar otros atrayentes como heces y pescado en descomposición.
En este estudio se encontró que a menor cobertura de vegetación se incrementaba el número de especies raras de mariposas. Muriel y Kattan (2009) comparando cultivos de café expuestos al sol y de sombra evidenciaron un cambio en el comportamiento de las mariposas, las cuales forrajeaban más en los cultivos de café de sombra y los usaban para su desplazamiento. Esto debido, probablemente, a la mayor presencia de plantas hospedantes de la familia Asteraceae, las cuales emplean para colocar sus huevos.
Muriel y Kattan (2009) y Muriel et al. (2011), indicaron que los agroecosistemas de café de sombra son favorables para las especies de mariposas, pero que no dependen únicamente del tipo de manejo que presente el cultivo, sino que también de otras condiciones como la diversidad de plantas hospedantes y el área de vegetación natural que las coberturas presenten. Esto coincide con lo descrito por Perfecto et al. (2005), quienes señalaron que la riqueza de mariposas aumenta a medida que aumenta el porcentaje de cobertura de los árboles que generan la sombra a los policultivos de café.
El análisis de correspondencia canónica reflejó que la mayoría de especies asociadas a las variables de la estructura de la vegetación pertenecían a la subfamilia Satyrinae. Las plantas hospedantes de estas mariposas no presentan metabolitos secundarios por lo que éstas últimas desarrollan coloraciones crípticas, y vuelan en el sotobosque donde hay altos niveles de humedad y pueden evitar la depredación (DeVries 1987). La alta complejidad estructural del cafetal bajo sombra facilita el mantenimiento de diversas especies de mariposas diurnas. Esto puede estar relacionado con la disponibilidad de refugio y recursos alimenticios para los adultos como flores, frutos y plantas hospedantes importantes para el desarrollo de los estados inmaduros de las mariposas asociadas al cultivo de café. Dado que en el departamento de Santander la mayoría de cafetales son de sombra, se debe evaluar con mayor profundidad el papel de estos sistemas productivos en el mantenimiento de la diversidad de mariposas a nivel regional.
Conclusiones
Los policultivos de café tradicionales bajo sombra pueden ser benéficos para la diversidad de lepidópteros debido a la heterogeneidad de su estructura vegetal que, a su vez, favorece una amplia disponibilidad de recursos y refugio para las mariposas.
En comparación con cultivos de café bajo sombra de otras regiones del país, el alto número de especies registradas en este estudio, así como el número de especies raras (50%), resalta la importancia de estos sistemas para el mantenimiento de la diversidad regional de mariposas en el departamento del Santander.
Las 49 especies encontradas en este trabajo constituyen el mayor número de especies publicadas hasta el momento para un cultivo de café de sombra tanto para el departamento de Santander como para Colombia.
Footnotes
Agradecimientos
A Cristóbal Ríos-Málaver por su colaboración en la identificación de las especies de mariposas. A Federico Álvarez Hincapie (Corporación Universitaria Lasallista de Medellín), José Fernando González-Maya (Fundación ProCAT-Colombia) y a Trevor Williams (Instituto de Ecología, A.C., México) por sus comentarios y aportes para el mejoramiento del documento. Al Laboratorio de Ecología Funcional de la Universidad Javeriana por el apoyo logístico (ID Proyecto 5696). A los evaluadores del manuscrito por sus sugerencias y comentarios. A Elia Isabel Acosta por su colaboración en el trabajo de campo. A los habitantes de las veredas San Francisco Macaregua de Curití (Santander).