
Abstract
The tribe Euglossini (orchid bees) is vital for tropical pollination, making their diversity and distribution across altitudinal gradients crucial for ecological studies and conservation. This study assesses Euglossini diversity in northern Colombia's Sierra Nevada de Santa Marta and Serranía del Perijá, focusing on altitudes from 600 to 2200 meters above sea level (m a.s.l.). Using Barcode Index Numbers (BIN) and the COI gene, 1258 male specimens were collected, with 415 individuals sequenced. We identified 21 Molecular Operational Taxonomic Units (MOTUs), from which BOLD: AEF4646 (Eufriesea) was identified as the most variable, BOLD: AEF4645 (Eufriesea) as the most divergent molecular species, and also species like BOLD: AAC0547 (Euglossa) are widely distributed in both SNSM and SP at 800 and 1000 m a.s.l., while BOLD: AAC3172 (Euglossa) is strongly associated with 800 m a.s.l. communities. Additionally, BOLD: ABY5852 (Euglossa) prefers sites between 1400 m and 1800 m a.s.l. Diversity analyses revealed no significant differences in species richness between the gradients, but richness declined with altitude. Eufriesea and Euglossa species were broadly distributed in the Sierra Nevada, while Euglossa and Eulaema dominated the Serranía del Perijá. Beta diversity analyses showed distinct distribution patterns, with intermediate elevations marking a transition zone. Above 2000 m a.s.l., evolutionary processes likely sustain Euglossini populations, emphasizing the role of these regions in bee diversification.
Introduction
Bees from the tribe Euglossini, or orchid bees, are key pollinators of various plant families and thrive in humid, lowland areas (Abrahamczyk et al., 2011; Ascher & Pickering, 2020; Ramírez et al., 2002). However, studies such as those by González et al. (2005) have facilitated the observation and collection of Euglossini along the Colombian Andean Mountain range, primarily below 2000 meters above sea level (m a.s.l.), with a few records extending up to 2500 m a.s.l. Similarly, Nates-Parra et al. (2006) assessed the diversity of bees of the family Apidae in the urban ecosystems of Bogotá, recording transient Euglossini species at elevations as high as 3000 m a.s.l. This latter finding aligns with the results of Parra-H. and Nates-Parra (2012). Colombia hosts around 134 Euglossini species across five genera: Aglae, Exaerete, Eulaema, Eufriesea, and Euglossa (Parra-H. & Nates-Parra, 2012; Parra-H. et al., 2016; Nates-Parra, 2016).
This study addresses a critical gap: the lack of data on Euglossini diversity and altitudinal distribution in the Caribbean region of Colombia. While research in the Andes and Pacific regions has provided valuable insights into their biology and distribution, the northern parts of Colombia remain largely underexplored (González et al., 2005; Martínez-Puc & Merlo-Maydana, 2014; Nates-Parra et al., 2006; Otero & Sandino, 2003; Parra-H. & Nates-Parra, 2012). The Sierra Nevada de Santa Marta (SNSM) and Serranía del Perijá, two isolated peripheral mountain systems in northern Colombia, are particularly interesting due to their sharp altitudinal gradients, unique climatic conditions, and geographical isolation. These factors create distinct biodiversity patterns and high levels of beta diversity, mainly in insects, making them critical for understanding species distributions and conservation strategies (Noriega & Realpe, 2018; Prieto et al. 2024; Prieto et al. 2008; Prieto & Vargas, 2016; Prieto & Rodriguez, 2007).
In Colombia, Parra-H. and Nates-Parra (2012) conducted a preliminary evaluation of the biogeographical classification of Euglossini species. Their study suggests that ecological factors such as flight efficiency, nectar intake, and social behavior shape the geographic distribution of these bees (Parra-H. & Nates-Parra, 2012). As habitat conditions shift, Euglossini species exhibit varying levels of specialization. They recommended including the Sierra Nevada de Santa Marta (SNSM) within the Inter-Andean provinces, highlighting the limited representation of the Caribbean province due to its low species diversity. A related study by Pérez-Buitrago et al. (2022) examined the seasonal distribution of Euglossini bees in the Orinoquía region, finding higher species abundance during the rainy season, corresponding to greater food and water availability for plants.
Studies in Brazil have revealed highly generalist and specialized Euglossini species, with their distribution closely tied to habitat characteristics (Oliveira & Campos, 1996; Sofia et al., 2005; Tonhasca et al., 2002a). In northern Brazil, generalist species dominate over specialized ones (Dec & Alves-Dos Santos, 2019). Ribeiro-Pinto et al. (2019) observed a marked decline in species richness and abundance along the altitudinal gradient of the Brazilian Atlantic Forest, primarily driven by temperature changes. Similar patterns have been documented in other insect groups, such as dung beetles, where richness decreases with altitude due to climatic and vegetation factors (Nunes et al., 2016). Rubio (2015) also observed slight variations in spider diversity along altitudinal gradients in the Argentine Yungas.
As isolated mountain systems, the SNSM and Serranía del Perija, are ecological outliers compared to the main Andean range. The SNSM, often described as a “mountain island,” is separated from the Andes by surrounding lowlands, which limit horizontal colonization and foster unique assemblages of locally adapted species (Noriega & Realpe, 2018). Over time, climatic changes and compressed vegetation zones have created barriers for vertical migrations, further influencing species distributions (Velásquez-Tibata et al., 2012). According to Noriega and Realpe (2018), the sharp altitudinal gradient of the SNSM leads to distinct transitions between lowland, mid-elevation, and high-mountain faunas, with a notable turnover zone between 1,200 and 1,600 m a.s.l. These patterns emphasize the importance of isolation and geography in driving biodiversity and underscore the need for targeted conservation efforts in these ecosystems.
Given these findings, further research along altitudinal gradients in Colombia is necessary to document species presence and habitat preferences. Such studies would clarify relationships between habitat characteristics, species occurrence, and evolutionary processes (Parra-H. & Nates-Parra, 2012). For instance, studies on Euglossini and dung beetles have shown that higher elevations are generally associated with lower species richness. However, these trends are not universal, highlighting the need for more specific, taxon-focused research.
Taxonomic challenges in Euglossini arise from morphological complexity, unclear descriptions, reliance on male traits, and inconsistent data collection (Bonilla-Gómez & Nates-Parra, 1992; Freitas et al., 2009; Parra-H. et al., 2016; Sheffield et al., 2009; Silveira et al., 2002). Specimen identification remains complex, and comprehensive taxonomic work is constrained by limited resources and a shortage of specialists (Paz et al., 2011; Silveira et al., 2002). This study uses Molecular Operational Taxonomic Units (MOTUs) and DNA barcoding to investigate Euglossini diversity along altitudinal gradients in the Sierra Nevada de Santa Marta and Serranía del Perijá. DNA barcoding, successful in other taxa, provides a detailed overview of Euglossini diversity in Caribbean regions and clarifies general distribution patterns (DeSalle et al., 2005; Floren et al., 2020; McFadden et al., 2019; Prieto et al., 2016; Prieto et al., 2008; Prieto et al., 2021; Janzen & Hallwachs, 2019; Paz et al., 2011; Ratnasingham & Hebert, 2013; Sheffield et al., 2009; Smith et al., 2005; Wong et al., 2009). DNA barcoding enhances the speed of ecological research by allowing species to be studied without traditional Linnaean binomial names. This method facilitates the rapid acquisition of biological knowledge and enables data comparison with other studies that use COI sequencing. Consequently, it improves the alignment between morphological and molecular identifications (López-Uribe & Del Lama, 2007). This research highlights the crucial role of Euglossini bees in tropical forest pollination and their contribution to ecosystem sustainability through essential cross-pollination (Parra-H. et al., 2016; Ramírez et al., 2002; Murren, 2002).
Materials and methods
Study area
The study was conducted in two areas of the Colombian Caribbean region: the Sierra Nevada de Santa Marta (SNSM) and Serranía del Perijá (SP). The SNSM (10°52′N, 73°43′W) is a pyramidal mountain system in Magdalena with diverse ecosystems ranging from semi-desert scrublands to montane forests and moorlands up to perpetual snow, spanning 0 to 5775 m a.s.l. (ProSierra, 2018; Tobias-Loaiza & Tamaris-Turizo, 2019). The SP (10°00′N, 72°57′W) extends across Cesar, La Guajira, and Norte Santander, bordering Venezuela, and features ecosystems from tropical dry forests to montane rainforests and paramos (Aguilera Diaz, 2016; Andrade Correa et al., 2019).
Sampling
Euglossine bees were collected under permit 2017094283-1-000 from ANLA, granted to Universidad del Atlántico. Sampling was conducted at 200-meter intervals along altitudinal gradients from 600 to 2500 m a.s.l. In the Sierra Nevada de Santa Marta (SNSM), stations ranged from Minca (600 m a.s.l.) to La Tagua village (2200 m a.s.l.). In the Serranía del Perijá (SP), stations ranged from Poso Azul (600 m a.s.l.) to El Cinco (2300 m a.s.l.). These elevations span multiple ecological zones and bioclimatic floors, as described by Josse et al. (2009), and transition from lowland tropical zones (infratropical, below 1,200 m a.s.l.) to montane cloud forests (mesotropical to supratropical, 1,200-2,800 m a.s.l.). While our analyses focus on individual elevations, we use these ecological classifications to contextualize species distribution patterns along altitudinal gradients.
Specimens were collected between 8:00 a.m. and 12:00 noon using baits of eucalyptol, cineole, and methyl salicylate (Dressler, 1982; Pearson & Dressler, 1985; Tonhasca et al., 2002b). 1258 male specimens were captured and preserved in 96 % ethanol. Each specimen was given an identification code and recorded with capture details, including date, location, altitude, and coordinates, using a Garmin eTrex10 GPS (Annex 1). A duplicate sampling was performed at each station, 10 meters away, to avoid interference between baits.
DNA extraction, purification, and sequencing
For DNA analysis, 415 individuals from each area were sampled. Each specimen was mounted on entomological pins, and the left foreleg was removed and placed individually in microplates of 95 samples with a drop of absolute ethanol. DNA extraction, amplification, purification, and sequencing of the mitochondrial COI gene barcode were performed at the Canadian Center for Barcoding Development (CCDB) in Ontario, Canada, following the protocol by Ivanova et al. (2006).
Data analysis
Delimitation of MOTUs and BIN codes (barcode index numbers)
Euglossini MOTU diversity index
To analyze Euglossini community structure along Colombia's altitudinal gradients in the Caribbean using COI gene data, R 4.4.1 GUI 1.80 (R Core Team, 2021) was employed, utilizing the following packages: Vegan (Oksanen et al., 2024), Biodiversity R (Kindt, 2024), BAT (Cardoso et al., 2015), and ggplot2 (Wickham, 2016). EstimateS 9.1.0 (Colwell, 2013) was used for descriptive community ecology.
Results
A total of 1258 specimens of the tribe Euglossini were collected: 957 from the altitudinal gradient of the Sierra Nevada de Santa Marta (SNSM) and 325 from the Serranía del Perijá (SP). The collected euglossine bees belonged to four genera, Euglossa, Eufriesea, Eulaema, and Exaerete, and were grouped into 21 MOTUs (Figure 1).
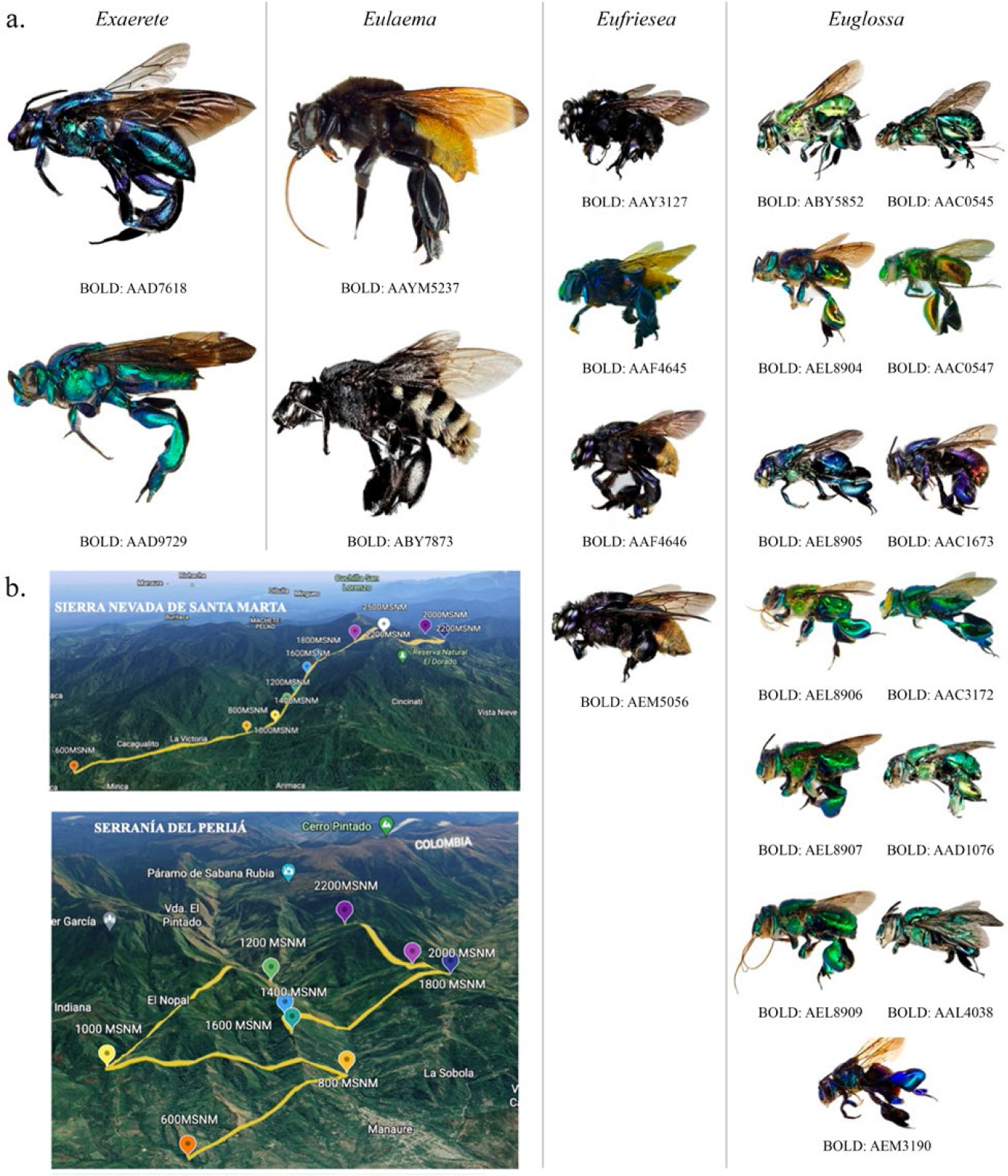
Molecular species from the Sierra Nevada de Santa Marta and Serranía del Perijá: a) The 21 MOTUs (greater than 500 bp) recovered, organized by genera, and b) the sampling stations in each area.
Delimitation of MOTUs using BIN codes (Barcode Index Number)
218 DNA sequences were compiled and grouped into 21 BIN numbers (MOTUs) from sequences of > 500 bp using BOLD Systems. Of these, 13 species belong to the genus Euglossa (BOLD:AEL8907, BOLD:AEM3190, BOLD:AAC0547, BOLD:AEL8906, BOLD:AAC0545, BOLD:AAC1673, BOLD:AAC3172, BOLD:AAD1076, BOLD:AAL4038, BOLD:ABY5852, BOLD:AEL8904, BOLD:AEL8905, BOLD:AEL8909), 4 to Eufriesea (BOLD:AEM5056, BOLD:AEF4646, BOLD:AAY3127, BOLD:AEF4645), 2 to Eulaema (BOLD:AAM5237, BOLD:ABY7873) and 2 to Exaerete (BOLD:AAD9729, BOLD:AAD7618). Recognizing 6 molecular species exclusive from the SNSM, 2 from SP and 13 present in both mountain configurations (Table 1). The 232 sequences obtained were easily aligned and analyzed in BOLDSystems and MEGA, comparing the 21 MOTUs at the inter- and intrageneric levels through paired genetic distances.
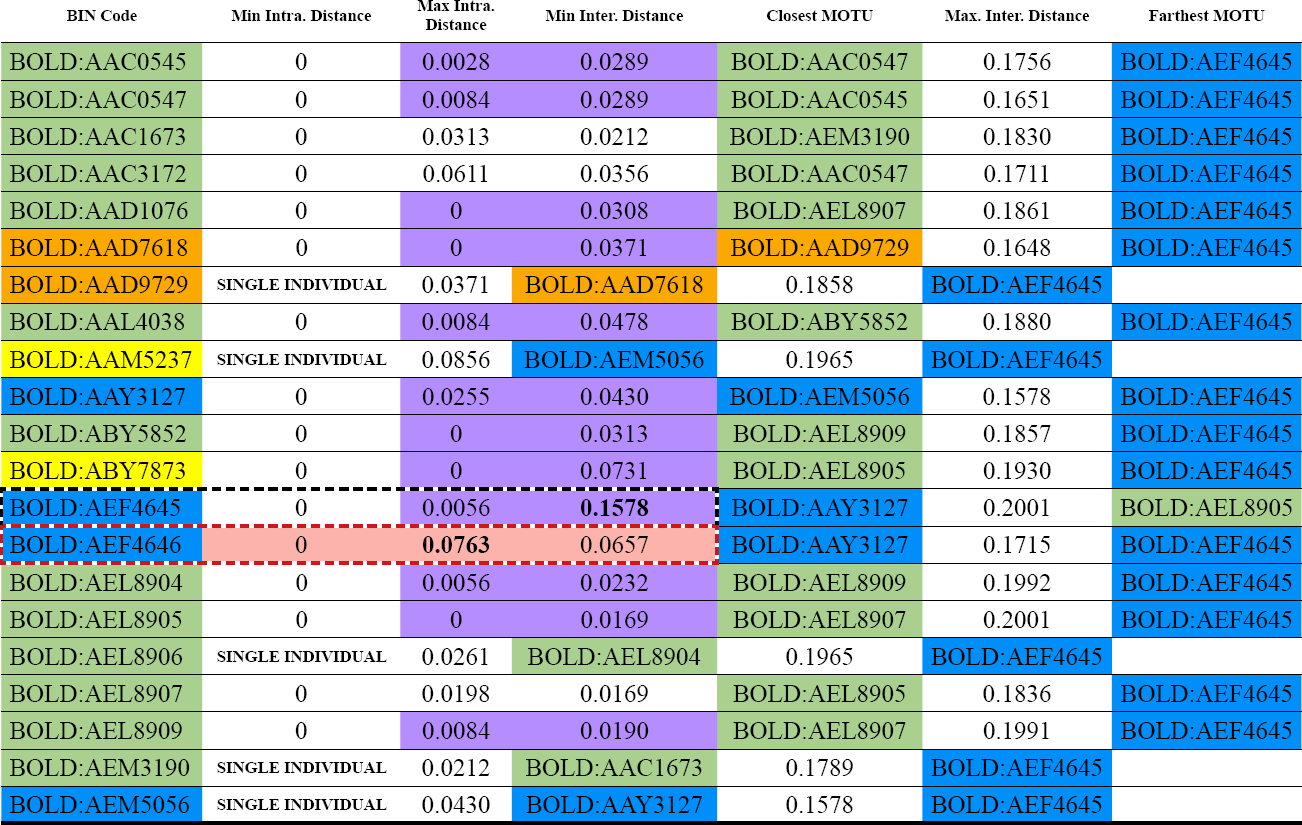
To establish barcode gaps, the maximum and minimum inter- and intraspecific distances were calculated (Table 2). The maximum intraspecific distance identified BOLD: AEF4646 (Eufriesea) as the most variable MOTU. The inter-specific distance analysis revealed BOLD: AEF4645 (Eufriesea) as the most divergent molecular species. A 6% genetic distance with a 1% average standard error was used to establish a general barcode gap for Euglossini, indicating the presence of distinct species when divergence exceeds 6 %. NJ-trees (Annex 2) indicate that species like BOLD: AAC0547 (Euglossa) are widely distributed in both SNSM and SP at 800 and 1000 m a.s.l., while BOLD: AAC3172 (Euglossa) is strongly associated with 800 m communities. Additionally, BOLD: ABY5852 (Euglossa) appears to prefer sites between 1400 m and 1800 m a.s.l.
Taxonomic genera and BIN numbers. BIN numbers associated with each of the four Euglossini genera and the study area where the taxa were collected are presented.
Inter- and intraspecific genetic distances for MOTUs from Sierra Nevada de Santa Marta (SNSM) and Serranía del Perijá (SP) were characterized, showing maximum and minimum values. The most variable species are highlighted with a dotted red border and colored pink, while the most divergent species have a double border. DNA barcode gaps are marked in purple. Species colors are assigned as follows: Euglossa in green, Exaerete in orange, Eulaema in yellow, and Eufriesea in blue.
The amplified sequences, recovered molecular diagnostic characteristics for 15 species with three or more sequences (Annex 3). 81 molecular characters were identified, with 54 being diagnostic or partial diagnostic for 12 of these species. However, no diagnostic characteristics were found for species like BOLD: AAC1673, BOLD: AEL8907, and BOLD: AEL8909, potentially due to their high genetic variability.
An altitudinal distribution graph (Figure 2) and a species abundance table for the 1258 individuals and 21 molecular species from SNSM and SP illustrate the broad distribution of the most common Molecular Operational Taxonomic Units (MOTUs) and the elevation preferences of the species.
Estimates of sampling richness and completeness indicate that expected species richness (SEst) increases with elevation in both regions. As elevation reaches 1400 m a.s.l. in our study, these values rise; however, at higher elevations an supratropical zones, species numbers are declining. This pattern may be linked to environmental conditions and the availability of resources such as food and shelter at different altitudes. In SNSM, sampling completeness was about 98.3 % (Chao1) and 77.8 % (Chao2), while in SP, it was 86.6 % (Chao1) and 75 % (Chao2). Suggesting, additional sampling is needed to enhance representativeness in both, SNSM and SP.
Rarefaction curves indicated no significant differences in species richness between SNSM and SP. Completeness was higher at mesotropical zones (600-1600 m a.s.l.) and decreased as elevation was gained in both regions.
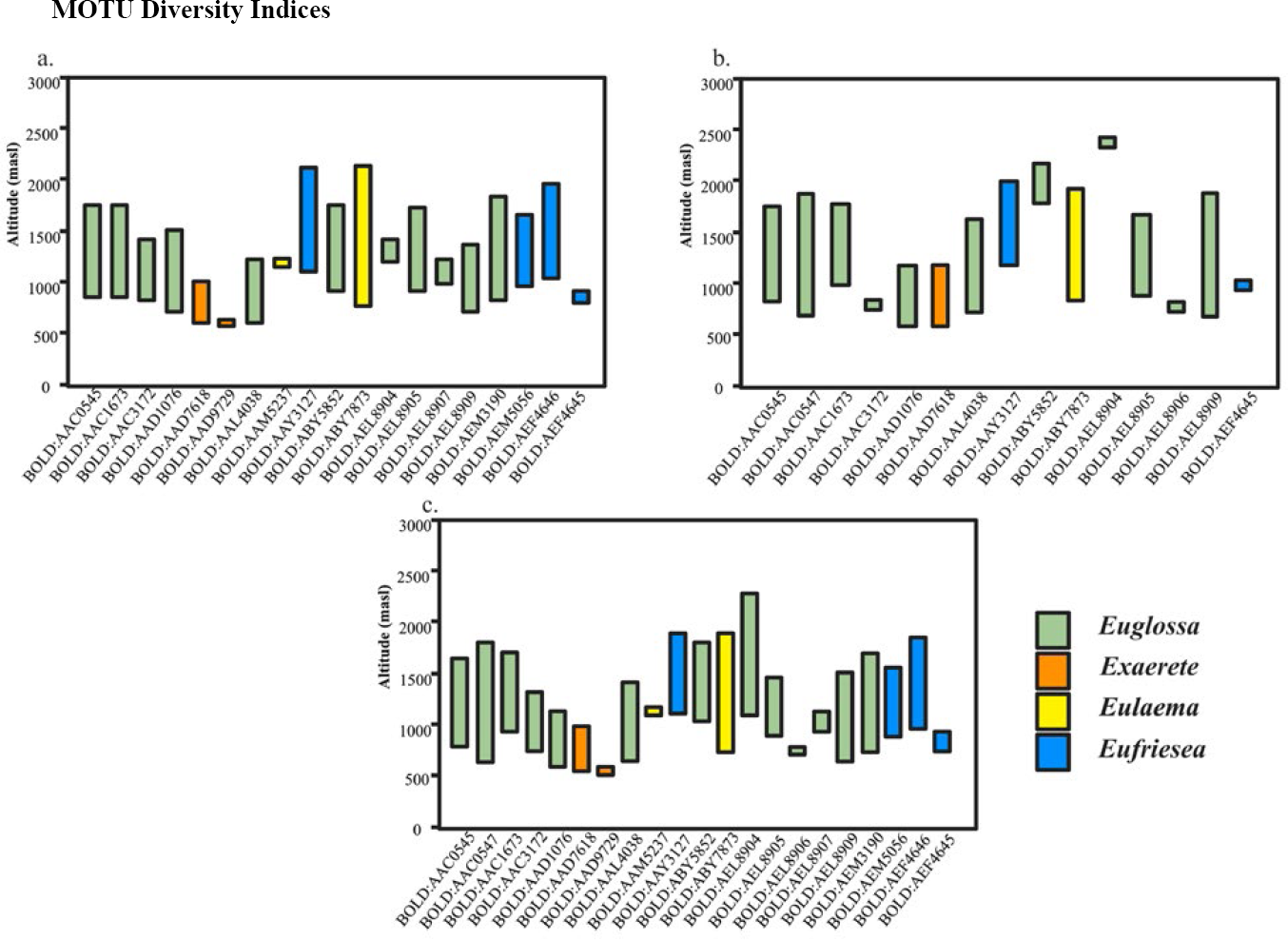
MOTUS observed distribution in the Sierra Nevada de Santa Marta (SNSM) and the Serranía del Perijá (SP). Presence/absence of MOTUs in each sampling site: a) Sierra Nevada de Santa Marta, b) Serranía del Perijá, c) Both mountainous configurations.
MOTU Diversity Indices
Rank-abundance curves were constructed to analyze the community structure of SNSM and SP. In the SNSM, Eufriesea and Euglossa species were broadly distributed along the altitudinal gradient, with key MOTUs such as BOLD: AEM5056 (Eufriesea, n = 159), BOLD: AAY3127 (Eufriesea, n = 118), BOLD: AAC1673 (Euglossa, n = 92), and BOLD: AAC0545 (Euglossa, n = 65) showing prominence.
Diversity indices (qD) and comparison graphs (Figure 3) along the SNSM gradient revealed high species richness (q0) at 800–1400 m a.s.l. This range corresponds to a transitional zone where lowland tropical forests (infratropical zones) begin shifting toward montane cloud forests (mesotropical zones).
Shannon diversity (q1) peaks at 800-1000 m a.s.l. and 2000 m a.s.l., suggesting that communities in these zones are more balanced. Similarly, Simpson diversity (q2), which emphasizes the most abundant species and reflects dominance within the community, also peaks at 800-1000 m a.s.l. (with species such as BOLD: AEF4646 (Eufriesea) and BOLD: AAC1673(Euglossa)) and 2000 m a.s.l. (represented by BOLD: AEL8905 (Euglossa)).
In SP, rank-abundance curves reveal BOLD: AAC0545 (Euglossa, n = 128), BOLD: ABY7873 (Eulaema, n = 50), and BOLD: AAC1673 (Euglossa, n = 43) as dominant species, indicating that Euglossa and Eulaema dominate along the altitudinal gradient. The highest species richness was observed at 600-1200 m a.s.l. (infratropical to lower mesotropical zones) and 1800-2000 m a.s.l. (upper mesotropical to supratropical zones). At 800 m a.s.l., Shannon diversity (q1) reaches its highest value, reflecting high species diversity combined with a more even community structure.
The q2 peaks at 800 m and 2000 m a.s.l., suggesting that dominant species, like BOLD: AAC0545, are primarily found at lower elevations, while the upper montane cloud forests within the supratropical zone experience harsh climatic conditions that favor a few highly adapted, dominant species as BOLD:ABY7873 (Figure 4).
Lower sampling completeness in SP, particularly at high elevations, may limit the complete detection of species diversity and beta diversity trends, emphasizing the need for further sampling in these areas.
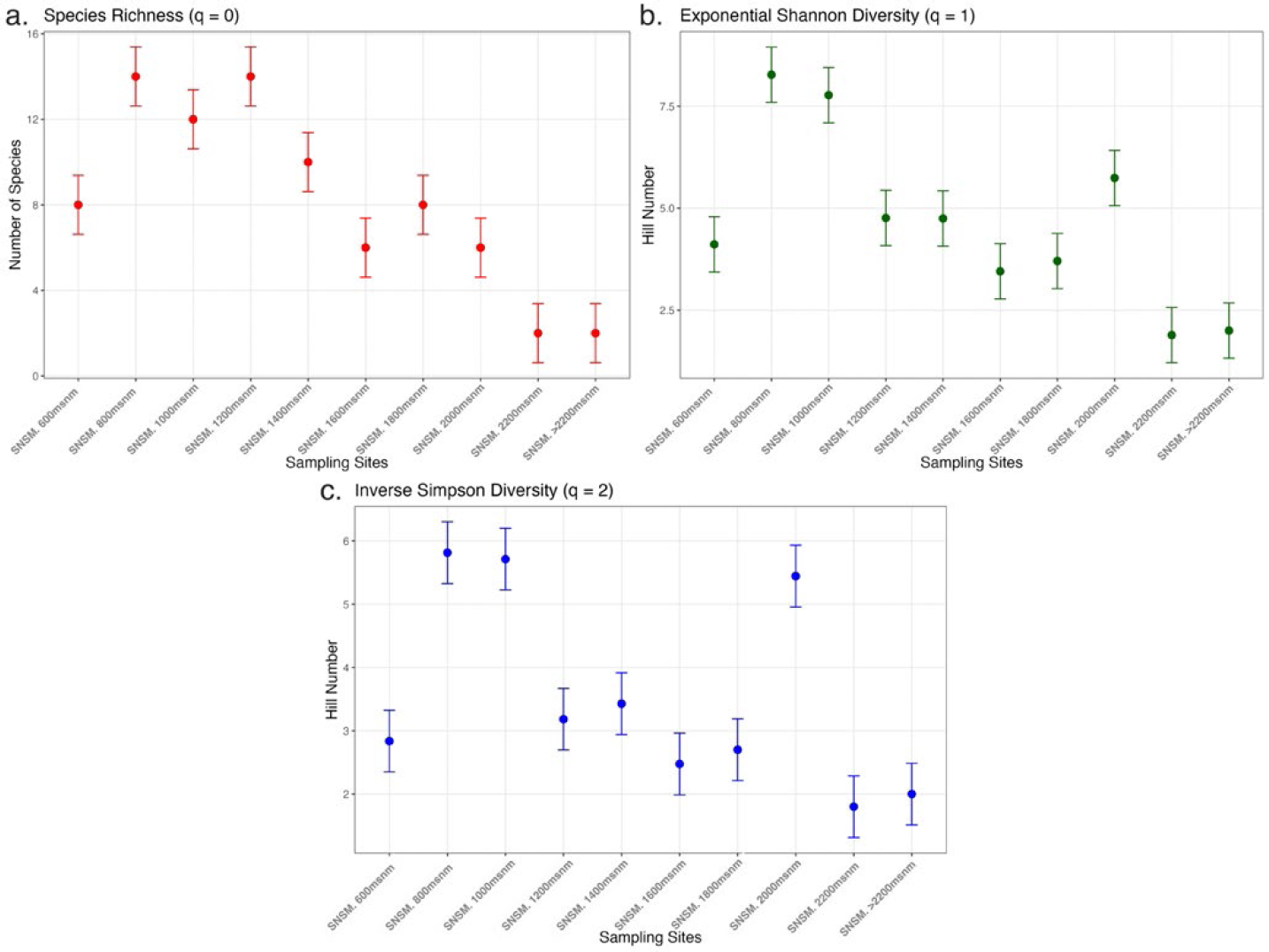
Comparison of Diversity Indices of Euglossini Communities in the Sierra Nevada de Santa Marta. a) Species richness (q0), b) effective number of equally frequent or common species (q1), and c) the effective number of dominant species (q2) are shown. Each point represents the estimated value of the respective diversity index for each sampling station.
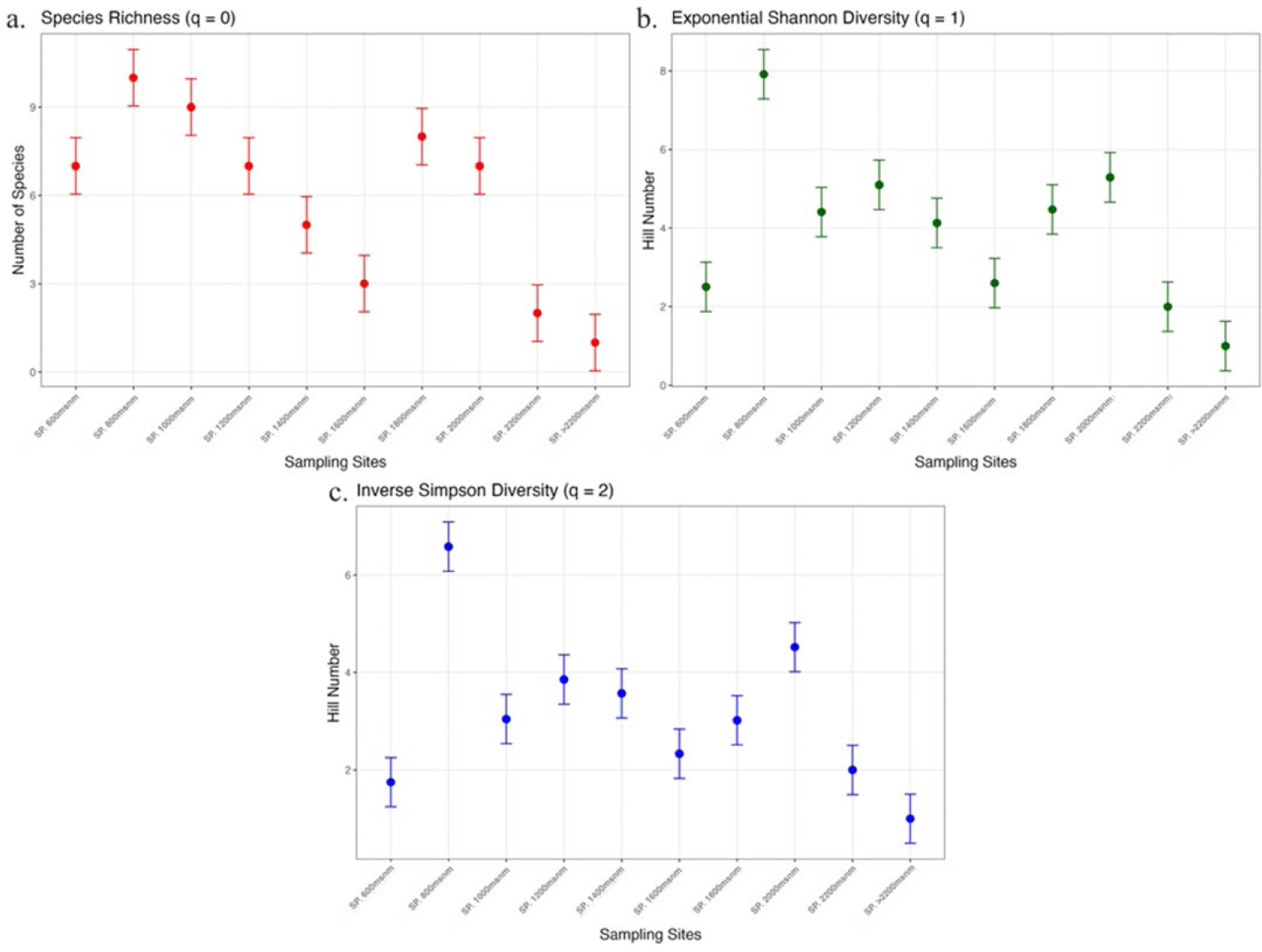
Comparison of Diversity Indices of Euglossini Communities in the Serranía del Perijá. a) Species richness (q0), b) effective number of equally frequent or common species (q1), and c) the effective number of dominant species (q2) are shown. Each point represents the estimated value of the respective diversity index for each sampling station.
Replacement of molecular taxonomic units with altitudinal gradients (β- diversity)
In the SNSM, the Jaccard index (IJ) and Sørensen's quantitative similarity coefficient (IS) were used to assess species richness and similarity among communities based on presence/absence and abundance (Villareal et al., 2004). The highest similarity in species presence was observed between communities at 1600 m and 2000 m a.s.l. (IJ = 0.67), while the most remarkable dissimilarity was between 600 m and 2200 a.s.l. (IJ = 0). Regarding species abundance, most substantial similarity was between 1200 m a.s.l. and 1400 m a.s.l. (IS = 0.73). This is due to the widespread presence and abundance of dominant species such as BOLD: AEF4646, BOLD: AEM5056, BOLD: AAY3127 (all Eufriesea), and BOLD: AAC0545 (Euglossa).
Dendrograms from IJ and IS analyses (Figure 5) show consistent clustering, separating higher elevation communities (supratropical zones, 2200 m a.s.l. and above) from those at lower and mid-elevations (infratropical to mesotropical zones, 800-1800 m a.s.l.). Communities at 1200 m and 1400 m a.s.l. (lower mesotropical zone), as well as those at 800-1000 m a.s.l. (upper infratropical to lower mesotropical zones) and 1600-2000 m a.s.l. (upper mesotropical to supratropical transition), share a high proportion of species.
In contrast, the community at 600 m (infratropical zone) is distinct, showing significant differentiation from higher elevation communities, with very low IS and IJ values (supratropical zone, 2200 m a.s.l.: IS = 0.0, IJ = 0.0; >2200 m: IS = 0.11, IJ = 0.2). These findings highlight clear habitat affinity patterns along the SNSM altitudinal gradient, reflecting transitions between tropical lowland forests and montane cloud forests as described by Josse et al. (2009).
The richness of SP communities, assessed using the IJ and IS indices, revealed distinct patterns. The highest similarity occurred between communities at 600 m and 800 m a.s.l. (infratropical zone, IJ = 0.55, IS = 0.71) and between 600 m and 1000 m a.s.l. (transition from infratropical to lower mesotropical zones, IJ = 0.60, IS = 0.75), largely due to the dominance of BOLD: AAC0545 (Euglossa). The greatest similarity was found between 1800 m and 2000 m a.s.l. (upper mesotropical to supratropical zones, IJ = 0.67, IS = 0.80), linked to the presence of BOLD: ABY7873 (Eulaema) and BOLD: AAC0545 (Euglossa).
In contrast, communities at 2200 m a.s.l. and above (supratropical zone) showed low similarity, likely due to limited species documentation.
The IJ and IS grouping trees (Figure 6) for SP revealed consistent clusters, with distinct elevation ranges. Intermediate elevations (800-1200 m a.s.l., upper infratropical to lower mesotropical zones) together, showing notable species turnover (IJ = 0.42, IS = 0.59). At higher altitudes (2200 m a.s.l. and above, supratropical zone), where few species are present, resource availability is a limiting factor.
A comparison of SNSM and SP communities assessed their similarity and species replacement. The results indicate that community similarity is largely preserved within each study area. SNSM communities exhibited greater shared species richness than SP communities, though rarefaction curves showed no significant differences between areas.
The highest similarity (IJ = 0.67) was observed at 600 m a.s.l. (infratropical zone), where species turnover is low. Similarly, the highest similarity in species abundance (IS = 0.8) between SNSM and SP communities was recorded at 600 m a.s.l., mainly due to Euglossa MOTUS BOLD: AAC0545 and BOLD: AAC1673. High similarity (IS = 0.8) was also observed within SP communities at 1800-2000 m a.s.l. (upper mesotropical to supratropical zones) and within SNSM communities at 800-1000 m a.s.l. (upper infratropical to lower mesotropical zones) (IS = 0.78).
The IJ and IS clustering trees (Figure 7) revealed consistent patterns, distinguishing altitudinal ranges. Communities above 2000 m a.s.l. (supratropical zone) and below 800 m a.s.l. (infratropical zone) displayed high similarity, while those at intermediate elevations (1000-2000 m a.s.l., mesotropical zones) were more distinct, indicating clear differences in community composition.
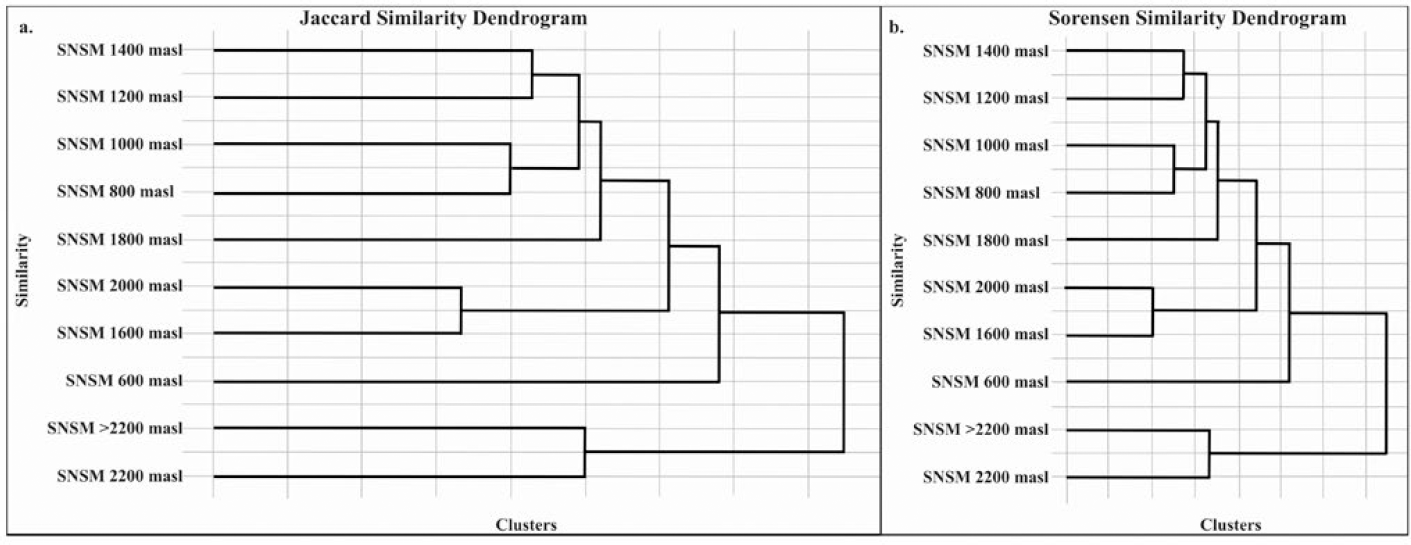
Grouping dendrogram for the Sierra Nevada de Santa Marta (SNSM) communities. a) Similarity between communities along the altitudinal gradient based on the Jaccard index (IJ). b) Similarity between the communities along the altitudinal gradient based on the Sørensen index (IS).
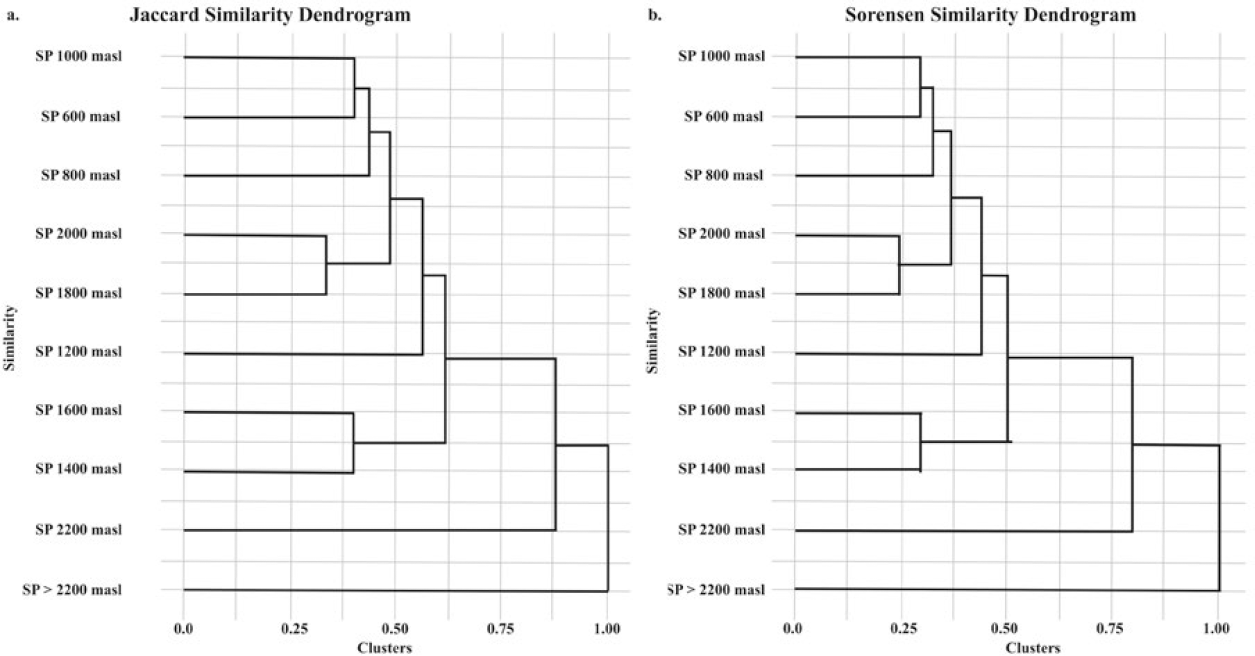
Grouping dendrogram for the Serranía del Perijá (SP) communities. a) Similarity between communities along the altitudinal gradient based on the Jaccard index (IJ). b) Similarity between the communities along the altitudinal gradient based on the Sørensen index (IS).
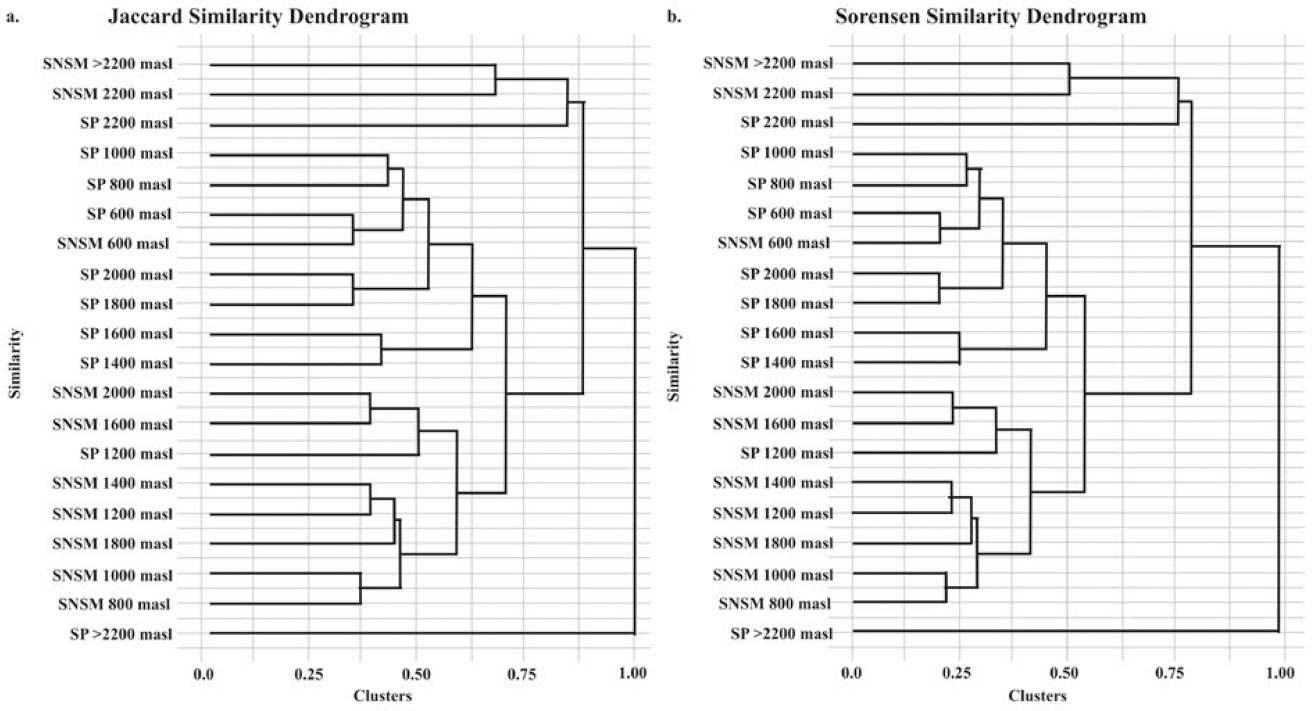
Grouping dendrogram for the Sierra Nevada de Santa Marta (SNSM) communities and the Serranía del Perijá (SP). a) Similarity between communities along the altitudinal gradient based on the Jaccard index (IJ). b) Similarity between the communities along the altitudinal gradient based on the Sørensen index (IS).
Discussion
Understanding the diversity of specialized pollinators, such as Euglossini bees, is crucial for effective conservation planning due to their importance in tropical ecosystems. By analyzing Euglossini diversity through Molecular Operational Taxonomic Units (MOTUs) along altitudinal gradients in the Sierra Nevada de Santa Marta and the Serranía del Perijá, we assessed their habitat preferences. We anticipated higher species diversity at lower elevations. This research, using MOTUs for the first time in the Colombian Caribbean, highlights the effectiveness of DNA barcoding in ecological studies. DNA barcoding clarifies species boundaries and uncovers hidden diversity and complements traditional methods, as noted by Mohammadi and Ahmadzadeh (2024). It also aids in documenting distribution patterns of these bees across altitudinal gradients.
Delimitation of MOTUs using BIN codes (Barcode Index Number)
The genetic distance analysis of Euglossini bees revealed significant variation among Molecular Operational Taxonomic Units (MOTUS), providing insights into species divergence and population structure across the Sierra Nevada de Santa Marta (SNSM) and the Serranía del Perijá (SP).
The MOTU BOLD:AEF4645 (Eufriesea) emerged as the most divergent, showing high interspecific distances compared to other taxa. This divergence likely results from limited gene flow influenced by geographic isolation, fragmented landscapes, and selective pressures across the mountainous regions of the Colombian Caribbean (Malpica-Topete, 2012; Stein et al., 2014). High-elevation zones, such as upper mesotropical, supratropical, and orotropical zones, with distinct climatic conditions may further exacerbate isolation, driving genetic differentiation and evolutionary divergence.
Conversely, BOLD:AEF4646 (Eufriesea) exhibited significant genetic variability, particularly across montane forests (800-2200 m a.s.l.) in the SNSM. The broader distribution and higher variability suggest a capacity for migration along the altitudinal gradient, potentially facilitated by the availability of floral resources and nesting habitats. This aligns with findings by Parra-H and Parra-Nates (2012), who reported that species adaptations to environmental gradients allow for broader occurrence patterns in complex montane systems.
These results underscore the role of geographic barriers and environmental selection in shaping genetic diversity within Euglossini bees. While geographic isolation drives genetic divergence in species like BOLD:AEF4645, environmental gradients promote intraspecific variability and population structuring in taxa like BOLD:AEF4646. Together, these patterns highlight the evolutionary processes operating within the fragmented landscapes of the SNSM and SP.
Molecular diagnostic characters recovered for most MOTUs further confirm the utility of DNA barcoding in distinguishing closely related species and identifying cryptic diversity. However, the absence of diagnostic characters in highly variable species, such as BOLD:AAC1673, suggests that additional data, including morphological traits, may be needed to resolve taxonomic ambiguities effectively. Integrating DNA barcoding with morphological analyses remains essential for refining species boundaries and understanding population structure (Ahmed et al., 2022 Kress; & Erickson, 2008; Will et al., 2005).
MOTU diversity indices
Alpha diversity analysis revealed clear patterns of species richness across the altitudinal gradients in both the Sierra Nevada de Santa Marta (SNSM) and the Serranía del Perijá (SP). Richness consistently peaked at mid-elevations (800-1200 m a.s.l.), corresponding to the upper infratropical and lower mesotropical zones, where favorable climatic conditions and high habitat heterogeneity promote species coexistence (Josse et al., 2009). These findings align with Josse et al. (2009), who reported that montane forests below 1500 m a.s.l. in the Northern Andes exhibit species richness levels comparable to those of lowland forests. The transitional nature of mid-elevations, which fosters overlaps between lowland and montane communities, likely explains the observed peaks in diversity.
In the SNSM, upper infratropical to lower mesotropical zones were dominated by species of the genera Eufriesea and Euglossa, with BOLD:AAC1673 (Euglossa) and BOLD:AEF4646 (Eufriesea) being particularly prevalent. This reflects a balanced community structure, consistent with findings by Noriega and Realpe (2018), who reported similar trends of high species diversity and compositional overlap at mid-levels before declining with increasing elevation. A comparable pattern was observed in the SP, where species like BOLD:AAC0545 (Euglossa) and BOLD:ABY7873 (Eulaema) strongly preferred preferences for upper infratropical to lower mesotropical zones. Support high richness due to their transitional nature, as observed in the results, offering optimal climatic conditions and sufficient floral and nesting resources to sustain diverse orchid bee communities (Abrahamczyk et al., 2011).
At higher elevations above 2000 m a.s.l. (the supratropical zone), species richness declined sharply in both regions. This reduction reflects the influence of harsh environmental conditions, including lower temperatures, reduced humidity, and limited resource availability, which impose strong ecological filters. Such constraints favor specialized taxa capable of adapting to these extreme environments, as evidenced by the presence of BOLD:AAY3127 (Eufriesea) in the SNSM (Josse et al., 2009; Noriega & Realpe, 2018).
In contrast, lower infratropical zone elevations (600-800 m a.s.l.) exhibited comparatively lower richness, likely due to increased competition, resource homogenization, and high temperatures and humidity. These factors may limit the distribution of certain orchid bee species, resulting in the dominance of fewer species such as BOLD:AAD7618 (Exaerete), which further reduces community evenness (Ramos et al., 2022).
Overall, the altitudinal patterns observed in both regions emphasize the role of elevation and environmental gradients in shaping species richness. Upper infratropical to lower mesotropical zones provide ideal conditions for the coexistence of diverse species, representing transitional areas, while higher elevations are defined by environmental filtering and specialization. The lower richness at low infratropical zone further highlights the influence of competition and resource distribution. These findings are consistent with broader trends reported for montane ecosystems in the Northern Andes (Josse et al., 2009) and underscore the importance of transitional areas in maintaining alpha diversity.
Replacement of molecular taxonomic units with altitudinal gradients (β- diversity)
Beta diversity patterns across the altitudinal gradients in the Sierra Nevada de Santa Marta (SNSM) and the Serranía del Perijá (SP) reveal clear species turnover and community differentiation driven by environmental gradients. Mid-elevations (800-1200 m a.s.l.) in both regions exhibit greater species overlap and compositional similarity, reflecting the transitional nature of these zones. The overlap likely results from the coexistence of lowland and montane species, supported by favorable climatic conditions and high habitat heterogeneity (Josse et al., 2009).
At higher elevations (above 2000 m a.s.l., supratropical zone), species turnover increases markedly, driven by the influence of environmental filtering. Harsh climatic conditions, such as lower temperatures, reduced humidity, and limited resource availability, impose constraints on community composition. These conditions favor specialized taxa like BOLD:AAY3127 (Eufriesea), which are adapted to survive in extreme environments. Similar patterns have been documented in other montane systems, where increasing altitude leads to reduced species similarity and greater compositional uniqueness (Josse et al., 2009; Nemésio & Vasconcelos, 2013; Noriega & Realpe, 2018).
In contrast, lower elevations (600-800 m a.s.l., infratropical zone) show reduced beta diversity and higher dominance of a few widespread species, such as BOLD:AAD7618 (Exaerete). This reduced turnover may result from resource homogenization and competitive exclusion, which limit species coexistence.
Geographical separation between the SNSM and SP further amplifies beta diversity, as the isolation of these two mountain systems has led to the development of distinct species assemblages. For instance, BOLD:AAD9729 (Exaerete) and BOLD:AAM5237 (Eulaema) were found exclusively in the SNSM, whereas BOLD:AAC0547 (Euglossa) and BOLD:AEL8906 (Euglossa) were unique to the SP. These patterns underscore the combined role of altitudinal gradients, environmental filtering, and geographical barriers in shaping species distributions and turnover across the two regions (Nemésio & Vasconcelos, 2013; Prieto et al., 2021).
Overall, beta diversity reflects the dynamic interplay between elevation-driven climatic constraints and geographic isolation. Mid-elevations emerge as areas of high species overlap, while higher altitudes promote uniqueness through strong ecological filtering. The observed trends align with broader findings in the Northern Andes, where environmental gradients act as critical drivers of species turnover and community differentiation (Josse et al., 2009; Nemésio & Vasconcelos, 2013; Prieto et al., 2021).
Conclusions
This study highlights the effectiveness of DNA barcoding as a tool for delimiting Molecular Operational Taxonomic Units (MOTUs) and assessing species diversity within the tribe Euglossini. Applying DNA barcoding along altitudinal gradients in the Sierra Nevada de Santa Marta (SNSM) and Serranía del Perijá (SP), 21 MOTUs were identified revealing complex patterns of species distribution, genetic divergence and population structure.
Alpha diversity peaks at upper infratropical and lower mesotropical zones (800-1200 m a.s.l.), where transitional conditions foster overlaps between lowland and montane species. Species richness declines at higher elevations (supratropical zone >2000 m a.s.l.) due to environmental filtering, favoring specialized taxa. Significant beta diversity was observed across altitudinal gradients and between mountain systems, driven by geographical isolation and environmental variation, promoting species turnover and evolutionary divergence.
The detection of significant genetic divergence, such as in BOLD:AEF4645 (Eufriesea), and high variability in taxa like BOLD:AEF4646, underscores the role of fragmented landscapes and environmental selection in shaping diversity. These findings highlight the importance of mountains as evolutionary hotspots and emphasize the need to conserve transitional and high-elevation habitats.
Our study advocates for integrative taxonomy, combining molecular techniques like DNA barcoding with traditional morphological methods, to improve species identification and ecological research. Expanding this approach to other regions and incorporating biogeographical analyses will further elucidate the evolutionary processes shaping Euglossini diversity.
In conclusion, this research enhances our understanding of Euglossini diversity and distribution, demonstrating the value of DNA barcoding in biodiversity studies and its potential to inform conservation strategies in tropical montane ecosystems.
Footnotes
Acknowledgments
We extend our gratitude to the Alexander von Humboldt Foundation under the Research Group Linkage Program (Evolution of the High Andean Insect Fauna project), which facilitated collaboration between Colombian and German teams, fostering new partnerships. We also thank all the entities that supported our research, including Finca La Victoria in Minca, Magdalena, and Mi Ranchito in Manaure Balcón Turístico del Cesar, Serranía del Perijá.
Annex 1.
Euglossini bees collected in the Sierra Nevada de Santa Marta and the Serranía del Perijá. The individuals collected are presented with their BIN code and information associated with the essence used and the location sampled. Where the species of each genus are recognized: Euglossa in green, Exaerete in orange, Eulaema in yellow and Eufriesea in blue.
Annex 2.
Neighbor joining tree for molecular species Sierra Nevada de Santa Marta (SNSM) and Serranía del Perijá (SP), for the Tribe Euglossini [A phylogenetic tree is displayed, showing the relationships between different Euglossini bee specimens based on their molecular data.]
Annex 3.
Molecular diagnostic characters Euglossini bees from the Sierra Nevada de Santa Marta and Serranía del Perijá. Color gradients indicate the type of character identified from red, showing diagnostic characters, to white, for those that do not provide relevant information for identification. [A character matrix is shown, highlighting nucleotide positions that are diagnostic for different BOLD species groups.]