
Abstract
The South American fruit fly, Anastrepha fraterculus (Diptera: Tephritidae), is an economically important pest of fruit production in Southern Brazil. In organically managed orchards the species has traditionally been controlled with oils, plant extracts, and solutions such as pyroligneous extract and lime sulfur. The objectives of this study were to examine the possible deterrent effect of pesticides with the highest electroantennographic bioactivity on fruit flies and to assess their effects on the viability of pupae in treated fruits. Antennae were exposed to pyroligneous extract (BioPirol7M®, 0.4%), lime sulfur solution (SulFertilizantes, 1%), neem (Organic Neem®, 0.5%), and rotenone (Rotenat®, 0.6%), taking into account fly sex, age and reproductive status. Pupal viability was assessed for larvae reared in papaya (Carica papaya var. Calyman) and guava (Psidium guajava var. Paluma) fruits treated with the pesticides that generated the strongest electrophysiological responses. The bioactivity of A. fraterculus antennae was highest when stimulated with pyroligneous extract and lime sulfur solution, for young and mated flies. Neither substance inhibited oviposition and larval development in treated fruits, a result that has important implications for A. fraterculus management in organic systems.
Introduction
Fruit flies rank among the most important pests in commercial orchards, because of the direct economic impact they have on fruit production and quarantine restrictions for fruit exports imposed by commercial patterns (Aluja 1994; Clark et al. 2005). Ovipositing flies puncture fruit that their larvae subsequently feed on, reducing their value or spoiling them altogether (Malavasi et al. 1994). Anastrepha fraterculus (Wiedemann, 1830) is common in citrus and rosaceous orchards in southern Brazil, where it outnumbers other flies in the same genus and the Mediterranean fruit fly, Ceratitis capitata (Wied. 1824) (Salles 1995).
In Brazil, fruit flies are mostly controlled with organophosphate insecticides. These are very toxic and not selective with regards to natural enemies (Kovaleski and Ribeiro 2003; Scoz et al. 2004; AGROFIT 2011). Full cover spraying is used in guava, stone fruit, and sweet passion fruit plantations, among others, while toxic baits are more commonly used in citrus orchards (Raga and Sato 2006).
In organic fruit plantations, pest control agents include plant oils and extracts such as neem (Azadirachta indica A. Juss, 1797) (Mordue and Nisbet 2000) and rotenone [Lonchocarpus utilis (Smith), Lonchocarpus urucu (Killip and Smith), Derris elliptica (Wallich) Benth and Derris malaccensis (Benth.) Prain] (Kathrina 2004; Wiesbrook 2004). Lime sulfur solution is also widely used in pest control (Bergamin Filho et al. 1995), as is pyroligneous extract (Azevedo et al. 2005; Morandi Filho et al. 2006; Kim et al. 2008). However, few studies have assessed the impacts of these substances on fruit flies (Gonçalves et al. 2005; Rupp 2005).
An electroantennography (EAG) bioassay was used to compare antennal receptivity to stimuli with neem, rotenone, lime sulfur and pyrolignous extract and to evaluate their potential as candidate substances for repelling A. fraterculus or deterring its oviposition on fruits. The results of EAG assays allow one to choose compounds perceived by the olfactory system of fruit flies and to discard those that are poorly perceived or not perceived at all.
The objectives of this study were to assess the electrophysiological activity of neem, rotenone, lime sulfur solution, and pyroligneous extract on the antennae of reared A. fraterculus of different sexes, ages, and reproductive status, and to assess the effects of the two latter substances on pupa viability.
Materials and Methods
Experiments were carried out with A. fraterculus individuals reared in the laboratory at the Universidade Federal do Rio Grande do Sul (UFRGS), southern Brazil, in 2010. Papaya (Carica papaya L. (Caricaceae) var. Calyman) fruits were used as the larval development substrate. Adults were fed on an artificial diet consisting of brown sugar, soy protein, and wheat germ at a 3:1:1 ratio. Insects were reared in an environmentally controlled chamber kept at 25 ± 2 °C, 70 ± 10% relative humidity, and a 12-12 L-D hour photoperiod.
Adult flies up to 24 hours of age were segregated into three groups: females, males, and both sexes together in 1.5L cages containing food and water, where they remained until they reached a suitable age for the bioassays. Electrophysiological responses to neem, rotenone, lime sulfur, and pyroligneous extract were observed in the antennae of 15 male and female A. fraterculus. It was accessed using flies from different ages [young (5 to 10 days old) and old individuals (25 to 30 days old)] and reproductive status (mated and unmated), totalizing 32 treatments. Couples kept together were considered mated. The tested substances were neem (Organic Neem®; 0.5%), rotenone (Rotenat®; 0.6%), lime sulfur solution (SulFertilizantes; 1%), and pyroligneous extract (BioPirol7M®; 0.4%), which were acquired from the manufacturers Dalquim, Natural Rural, SulFertilizantes, and BioCarbo, respectively. All substances were diluted in distilled water and prepared on the day the bioassays were performed at manufacturer-recommended concentrations.
The electroantennographic methods used in this experiment are similar to those described by Trimble and Marshall (2007), in which each antenna was attached to a two-filament silver electrode using conducting gel (Spectra 360, Electrode Gel-Parker). The analog responses of the signal (in millivolts) were captured, amplified, and processed with a data acquisition controller (IDAC-4, Syntech®), and subsequently recorded using EAG 2000 software (Syntech®). Antennae were stimulated with 5µL of each substance. Twenty-four hours before the electrophysiological tests were carried out, individual flies were placed into 500mL plastic cages with only distilled water.
The data related to the variables involved in the size of the EAG responses in millivolts (mV) was analyzed via a multiple-comparison General Linear Model followed by the Least-Significant Difference (LSD) test and expressed as the eta-squared (ηβ) index, using SPSS 17 software. Response sizes (mV) were compared with Kruskal-Wallis (α = 0.05) and Mann-Whitney tests using BioEstat 5.0 software.
Viability of A. fraterculus pupae was assessed in papaya (var. Calyman) and guava (var. Paluma) fruits that had been submerged for five seconds in lime sulfur solution (SulFertilizantes) (1%), pyroligneous extract (Biopirol 7M®) (0.4%), or distilled water (control). These substances were selected because they generated the strongest electroantennographic responses in the previous experiment.
A set of three fruits of the same species, each subjected to one of the treatments, was placed simultaneously on regularly spaced Petri dishes inside 350 cm³ plastic cages covered with voile. In each cage were placed dishes of water, food, and 15 mated female A. fraterculus that were 20 to 25 days old. The position of fruits within the cages was randomized for each of the 18 replicates per species. The bioassay was carried out in an environmentally controlled chamber under the same conditions as rearing.
The flies remained with the fruits for 48 hours. At the end of this period fruits were removed and stored in 500 mL containers that were 1/3 full of sterilized sand and covered with voile. After 20 days the fruits were removed, the sand sifted, and the pupae counted. Pupae were transferred to 500 mL containers with 2 cm of sterilized sand. The containers remained covered with voile for up to 30 days, during which time the number and sex of emerging insects were recorded.
Three guava and three papaya were stored in containers with sterilized sand, covered with voile, for 30 days, in order to determine the potential for prior infestation by fruit flies. Three other fruits (guava and papaya) were placed inside a rearing cage with approximately 150 pairs of A. fraterculus for 48 hours to ensure that the fruits were appropriate for insect development. These fruits were then transferred to 500 mL containers with sterilized sand and kept there for 30 days, at which time the sand was sifted and the pupae counted.
The numbers of pupae and emerged insects were square root-transformed and compared among treatments using the Kruskal-Wallis test (α = 0.05).
Results
Proportions of the variance in electroantennographic responses of Anastrepha fraterculus accounted for by treatments and their interactions, and their respective significances (P), according to a General Linear Model followed by a Least Significant Difference test, expressed as the eta-squared (ηβ) coefficient function.
Regardless of age, sex, and reproductive status, electroantennographic responses of adult A. fraterculus were significantly stronger when insects were stimulated with lime sulfur solution and the pyroligneous extract than with rotenone or neem (H = 77.183; df = 4; P < 0.0001) (Fig. 1). Males only showed significantly stronger responses than females for neem (H = 97.130; df = 9; P = 0.023), reflecting the low explanatory power (4%) of the association between pesticide and sex (Table 1).
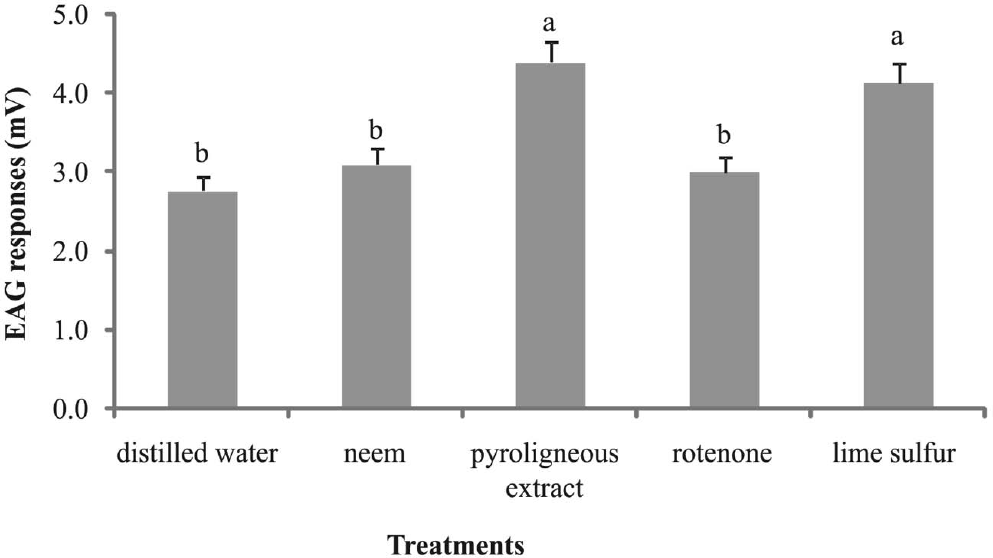
Mean electroantennographic responses of Anastrepha fraterculus to distilled water, 0.5% Neem (Organic neem®), 0.4% pyroligneous extract (Biopirol 7M®), 0.6% rotenone (Rotenat CE®), and 1% lime sulfur solution (SulFertilizantes). Columns with different letters differed significantly in a Kruskal-Wallis test (P<0.05) (n = 120).
Mated flies showed stronger responses than unmated flies (Z = 6.454; df = 2; P < 0.0001) and young flies showed stronger responses than old flies (Z = 6.282; df = 2; P < 0.0001). The eta-squared coefficient indicated that 4.5% of the variance in electroantennographic responses of A. fraterculus was accounted by the association between age and reproductive status, while 3% by reproductive status and pesticide (Table 1). Young mated flies showed significantly stronger responses than old mated flies, for all treatments (H = 104.76; df=9; P<0.0001) (Fig. 2). Young unmated flies only showed stronger responses than old unmated ones for pyroligneous extract (H = 45.167; df = 9; P = 0.025).
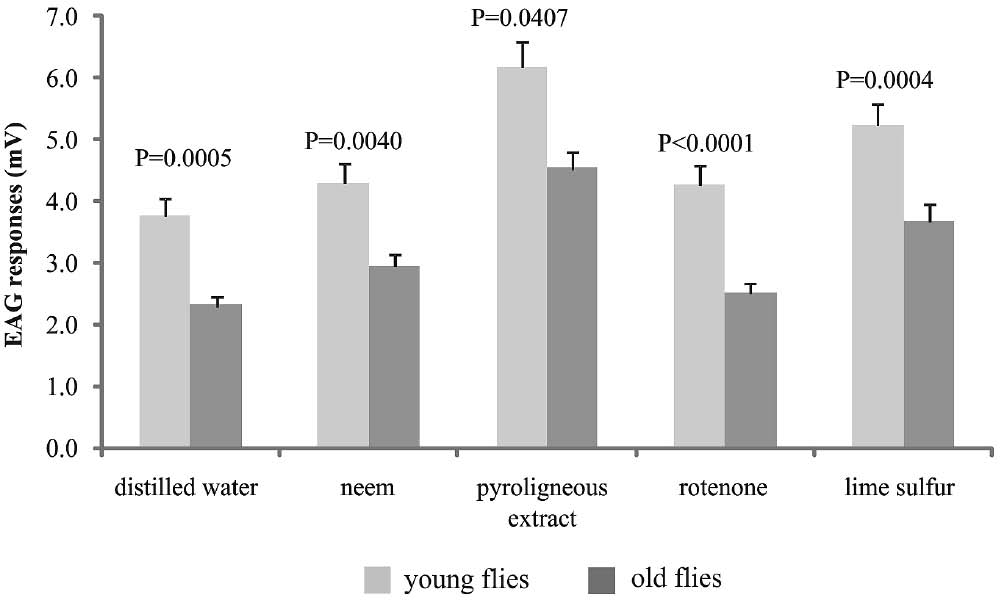
Electrophysiological responses of young and old mated Anastrepha fraterculus to distilled water, 0.5% Neem (Organic neem®), 0.4% pyroligneous extract (Biopirol 7M®), 0.6% rotenone (Rotenat CE®), and 1% lime sulfur solution (SulFertilizantes). P-values (P) are given above the bars. Kruskal-Wallis (P < 0.05) (n = 30).
No pupae were observed in the fruits stored in containers with sterilized sand. By contrast, pupae were recorded in the fruits stored in a rearing cage. These results were not statistically analyzed.
Discussion
Although female A. fraterculus responded selectively to pyroligneous extract and lime sulfur solution in the electroantennographic bioassays, the results of the oviposition test showed that these substances did not prevent egg laying and the subsequent development and emergence of A. fraterculus. Lime sulfur solution is a leaf fertilizer and fungicide traditionally used to repel certain species of insects. The elemental sulfur naturally present in the waxy cuticle of gymnosperms and angiosperms may play a role in plant defense mechanisms (Burow and Wittstock 2008), and can also induce the production of antifungal substances (Cooper and Williams 2004). Under field conditions, the toxic effect of lime sulfur solution on insects and mites is produced by the release of hydrogen sulfide (H 2 S) and sulfur colloids (Abbot 1945).
Sulfurous volatiles emitted by plants play a role in chemical defense (Rouseff et al. 2008). According to Rouseff et al. (2008), the dimethyl disulfide and trimethyl disulfide emitted by guava leaves may be primarily responsible for protecting against attacks by the psyllid Diaphorina citri Kuwayama, 1908 (Hemiptera, Psyllidae). By contrast, sulfurous compounds emitted by onion, Allium cepa L. (Liliaceae), attract Delia antiqua (Meigen) (Diptera, Anthomyiidae) (Matsumoto, 2008). In electroantennographic bioassays, Gouinguené et al. (2005) demonstrated that females of that species oviposited significantly more in the presence of n-propyl disulphide (Pr2S2).
In field conditions, Afonso et al. (2007) reported a decrease of 79.1% in infestations of the European peach scale, Parthenolecanium persicae (Fabricius, 1776) (Hemiptera, Coccidae), in vineyards treated with lime sulfur solution (0.5%). Likewise, Bellon et al. (2009) documented a 28.9% decrease in oviposition by Vatiga manihotae (Drake, 1922) (Hemiptera: Tingidae) in leaves of Manihot esculenta Crantz. According to Afonso et al. (2007) and Bellon et al. (2009), these species deposit their eggs on the host's cuticle, where the presence of sulfurous compounds may have a deterrent effect and inhibit oviposition. Such an effect, however, was not evident in A. fraterculus.
The stronger electroantennographic responses of A. fraterculus to Biopirol may be attributed to the presence of acetic acid in its composition. That compound is typically present in fruits, where it is a product of the fermentation process (IAEA 2003; Zhu et al. 2003). The same acid, in the form of vinegar, has been used in traps to monitor A. fraterculus populations (Salles 1999; Lemos et al. 2002; Monteiro et al. 2007) and has been characterized as an attractant for A. suspensa (Robacker et al. 1997; Robacker and Heath 1997; Robacker et al. 1998; Robacker et al. 2011) and C. capitata (Joachim-Bravo et al. 2001). Santos and Wansen (2006), however, noted that pyroligneous extract was ineffective at controlling A. fraterculus in organically managed apple orchards in Caçador, Santa Catarina, Brazil. Similarly, Morandi Filho et al. (2006) reported that the substance did not affect survival of Trichogramma pretiosum Riley, 1879 (Hymenoptera, Trichogrammatidae) in laboratory conditions.
In our study, while A. fraterculus showed electroantennographic responses to volatiles of lime sulfur solution and pyroligneous extract, we observed no deterrent effect on oviposition in fruits exposed to these substances. The lack of significant differences in pupae viability between fruits treated with water, solution, and extract may be explained by the fact that eggs are deposited in the fruit interior, away from the substances' potential insecticidal effects. Our results support those of Efrom et al. (2011), who also demonstrated that treating artificial fruits made of agar with both substances had an ineffective deterrent effect on A. fraterculus oviposition. Those authors also demonstrated that even the topical application of these substances on flies had no insecticidal action.
The fact that mated flies showed stronger electroantennographic responses than unmated flies might be related to physiological changes following mating. In Anastrepha ludens (Loew, 1873) (Diptera, Tephritidae) it has been observed that the olfactory perception of antennae changed after mating, in such a way that certain odor became more or less perceptible than others, including those involved in signaling attractiveness or repellence (Robacker et al. 1990). According to Metcalf and Metcalf (1992), mated females' greater olfactory sensitivity to host plant volatiles reflects the need for quick and selective orientation in finding the best oviposition sites, thereby favoring the survival of offspring.
Electroantennographic responses of young A. fraterculus were stronger than those of old individuals. A similar result was found by Kendra et al. (2005) for A. ludens when exposed to a bait of ammonium bicarbonate.
Our results suggest that, despite the traditional use of lime sulfur solution and pyroligneous extract in organic fruit orchards, these substances are not effective in reducing populations of A. fraterculus or in deterring oviposition and avoid damage in fruit.
Footnotes
Acknowledgements
The authors are grateful for the financial support provided by Brazil's National Council for Scientific and Technological Development (CNPq).