
Abstract
Mycotretus apicalis is an erotylid beetle that is commonly found in cultivations Pleurotus sajor-caju in Minas Gerais, Brazil and could potentially be a pest. The life cycle is studied for the first time by laboratory rearing using dried P. sajor-caju mushrooms as both a feeding resource and an oviposition substrate. M. apicalis species has a generation time of about three weeks, with the egg-adult period was about 22 days, four larval instars, and the average longevity for adults of 89.7 ± 32.89 days. These results suggest that the M. apicalis has a potentially high rate of population increase, indicating its great potential to be a pest insect.
Introduction
The family Erotylidae, with about 3,200 reported species, comprises exclusively mycetophagus insects that feed mainly on the fruiting bodies of basidiomycetes (Węgrzynowicz 2002). According to Skelley (1988), a wide variety of basidiomycete fungi serve as a host to the family; however, each species seems to be specific to a particular group of fungi. Some species have been reported to cause economic losses in commercial edible mushroom cultivation, since the larvae and adults consume the fruiting body (Valencia and Lopes 2005; Gaitán-Hernandez et al. 2006; Sato 2003).
The genus Pleurotus is one of the most edible mushroom cultivated in Brazil due to low cost, easy availability of substrates for its cultivation and high biological efficiency, adapting well to the condition of tropical climate (Azevedo et al. 2009; Souza Dias et al. 2003). Sciarid fly and coleopteran species are the major pests in Pleurotus cultivation.
Mycotretus apicalis Lacordaire, 1842 has been found to cause injuries and economic losses in commercial cultivations of Pleurotus sajor-caju (Fr.) Singer in southern Minas Gerais, Brazil (Moreira et al. 2009). Thus, data on the rearing and life cycle of this species may contribute significantly to the implementation of an effective program to control this pest insect. Moreover, the life cycle of M. apicalis is unknown so the results can contribute in further studies like systematic and ecology of erotylids.
Materials and Methods
The experiments were carried out under controlled temperature (25 ± 2°C), humidity (70%) and photoperiod (14 h) conditions at the Laboratory of Insect Pathology, Department of Entomology, Federal University of Lavras (UFLA), Lavras, MG, Brazil.
Thirty adults of M. apicalis were collected in the same cultivation area of P. sajor-caju and kept in a beaker (500 mL) closed with a filter paper disk, which contained pieces of fresh mushrooms and two filter paper disks at the bottom of the vial. After 48 h, the adults were removed with a glass tube, and the presence of eggs and larvae were observed.
All of the rearing were done in polystyrene Petri dishes (15 cm diameter) with holes at the top that were covered with moistened hydrophilic cotton pieces, and two filter paper disks, that were moistened with 3 mL of distilled water, to maintain humidity. About 4 g of dried basidiocarps of P. sajor-caju were used as a feeding substrate, as suggested by Sato et al. (1999). First instar larvae hatched were transferred to the dishes using a brush. The maintenance of the reared population was done every 48 h, with cleaning and provision new food. After their feeding, the larvae, and afterwards the pupae, were transferred to dishes containing only 2 mL of distilled water. Once emerged, adults were transferred to dishes containing dried mushrooms. Dishes that had eggs in the gills of the mushrooms were kept for up to four days in a B.O.D chamber (T: 25 ± 2°C, RH: 70% and 14L10D), then the larvae hatched were transferred to Petri dishes repeating the rearing process.
Upon reaching the fourth instar, each larva was transferred to 5-cm-diameter Petri dishes. The dishes had holes in their lids, which were covered with moistened cotton. Two filter-paper disks at the bottom, moistened with 200 μl of distilled water, also contained pieces of dried mushrooms. After pupation, they were transferred - as described above - to Petri dishes that had only two filter paper disks moistened with 150 μl of distilled water.
About seven days after the emergence of adults (as the species lacks dimorphism), they were marked and randomly separated into pairs so that the mating behavior could be observed. Copulating couples were then set aside in 5 cm-diameter Petri dishes with pieces of dried mushrooms, as described previously. The presence or absence of eggs was observed after 24 h. The adults on whose dish we found eggs were confirmed to be a couple. We obtained a total of 19 couples. The number of eggs laid by the female was observed daily. The adults were sexed by dissection after their death, and their longevity was then evaluated.
The life table of fertility was drawn using the obtained data, as suggested by Silveira Neto et al. (1976), where: x = the midpoint of age range; lx = the age specific survival rate during stage x; and mx = the age specific fertility rate, that is, the number of offspring produced per female at age x that originates females. The parameters of population growth, Ro (net reproductive rate), T (mean generation time), rm (intrinsic rate of increase) and λ (finite rate of increase), were calculated according to Maia et al. (2000).
Results and Discussion
These insects consumed large amounts of dried mushrooms during the larval stage. We also observed that at the end of the fourth instar, after the larvae had ceased their feeding, they gathered at the edges of the dishes. This behavior is commonly found in some members of the family Erotylidae, as cited by Robertson et al. (2004).
Despite consuming mushrooms in smaller quantities, adults use their lamellae for oviposition. According to Węgrzynowicz (2002), erotylid couples copulate near or on the fruiting body. The females prefer to deposit their eggs in dried mushrooms with intact gills, and this pattern is obtained when using mushrooms harvested before the beginning of the senescence process (margin of pileus plane). Sato et al. (1999) observed a similar behavior in the female of Dacne picta Crotch, 1873 (Erotylidae). According to these authors, females of D. picta can distinguish the quality of mushrooms, so they oviposit on the freshest mushrooms, which are more favorable for the development of their offspring.
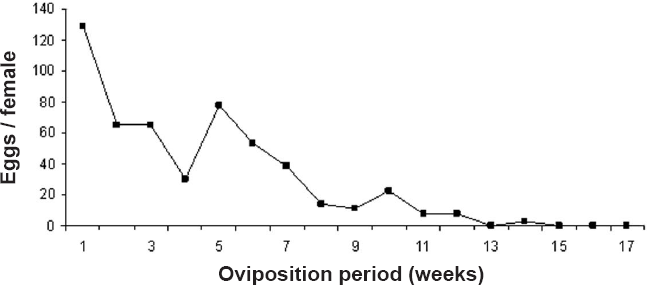
Average number of eggs per female during the oviposition period (weeks) for Mycotretus apicalis.
Duration of the stages of development (mean ± SE), survival (%) and mortality (%) of Mycotretus apicalis on dried Pleurotus sajor-caju mushrooms (T: 25 ± 2°C, RH: 70 ± 10%, LD: 14:10).
The longevity of adults was, on average, 89.7 ± 32.89 days. In 82% of the studied couples, the males showed greater longevity (102.95 ± 26.68) than the females (76.47 ± 33.8). As indicated by Townsend et al. (2006), the process of reproduction may have a high energy cost. Thus, one of the possible causes of lower longevity in the female is the high energy consumption involved in their oviposition process.
The biological parameters determined in this study indicate that the population of M. apicalis may increase 230.78 times (Ro) in each generation, and the mean generation time (T) is about three weeks. The population had a weekly increase of 46% [finite rate of increase (λ) = 5.46], and the intrinsic rate of increase (rm) was 1.69.
The highest rate of population increase, which corresponds to the meeting point of the age-specific fertility rate (mx) and age-specific survival rate (lx), occurred in the first week (Fig. 2).
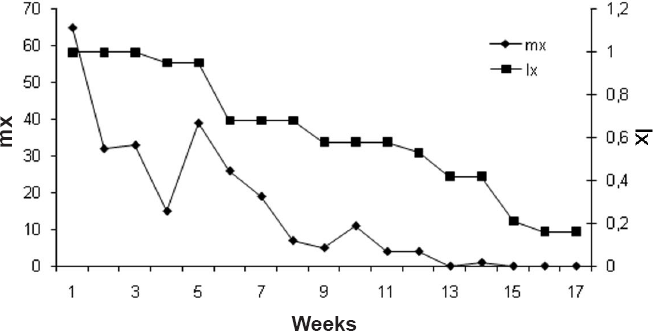
Midpoint of age range (mx) and age-specific survival rate (lx) of Mycotretus apicalis.
According to Price (1997), the intrinsic rate of increase (rm) can be used to compare growth rates among different species or growth rates of the same species under different conditions. Therefore, this datum will be of great importance, as it facilitates the selection of natural enemies to control this pest in Integrated Pest Management (IPM) programs.
It is important to note that these parameters of population growth were obtained under optimum conditions of humidity, temperature and the amount of resources for the development of the insect (similar to mushrooms house conditions). According to Odum (1988), under natural conditions, interactions within the population, as well as external environmental resistance, slow the growth rate, interfering with the determination of the form of population growth.
The results of this study show that M. apicalis has potential for a high rate of population growth, indicating its great potential as an insect pest of edible mushrooms. These data can be used to implement programs of integrated pest management and to contribute in further studies like systematic and ecology of erotylids.
Footnotes
Acknowledgments
The authors would like to thank to "Conselho Nacional de Desenvolvimento Científico e Tecnológico” (CNPq) for granting a scholarship to the first author, the “Fundação de Amparo à Pesquisa do Estado de Minas Gerais” (FAPEMIG) for their financial support and Sandro Carmelino for his help in the abstract translation.