
Abstract
To complement biological studies developed at CIAT on grass-feeding spittlebugs (Hemiptera: Cercopidae), a life table was established for the specific case of Zulia carbonaria (Lallemand). The cohort life table was conducted in the laboratory (27°C, 100% RH, continual darkness) for the eggs and in the screenhouse (19.5/29.5°C min./max., RH min. 56.3%) for the nymphs and adults. The survival exhibited by Z. carbonaria was 53.4% (egg-adult). The total mortality exhibited by the immature stages of Z. carbonaria was 46.4% divided in 10.1 and 36.3% for the eggs and nymphs, respectively, with a “killing-power” (kx) of 0.27. For the fertility table, the mean longevity of females was 29.3 days, with a mean fecundity of 125.8 ± 82.9 eggs/female and a fertility of 97.9%. The generational time (T) was 67.4 days, with a net reproductive rate (Ro) for (T) of 33.6 times. The intrinsic rate of natural increase (rm) showed that Z. carbonaria produces 0.05 females/day during the generational time. In addition, it was determined that the females presented a finite rate of multiplication (λ) of 1.05 females/day. This information is the first of its kind for spittlebug species in Colombia, and constitutes an appropriate tool to determine the quality of populations obtained from artificial colonies or under different habitat and host plant conditions.
Introduction
Según Lapointe y Miles (1992) el salivazo de los pastos es considerada la plaga más limitante en la actividad ganadera en América Tropical. A pesar de esta connotación es notoria la falta de estudios básicos sobre la biología y comportamiento del insecto que permitan entender su comportamiento en el campo, comprometiendo la efectividad de cualquier táctica de control. En el CIAT en los últimos años se han desarrollado investigaciones sobre la biología comparada del salivazo de los pastos. Estos estudios han cuantificado la variación a través del complejo en ciertos componentes biológicos. Por ejemplo, entre las especies predominantes en Colombia, el ciclo de vida varia entre 45 y 75 días (Peck et al. 2002a; Rodríguez et al. en preparación), y dependiendo de la región geográfica, el salivazo logra de dos hasta seis generaciones al año (Castro et al. 2005; Peck et al. 2002a; 2002b). Además, aunque la literatura reporta al suelo como el sustrato de oviposición más común para este grupo de insectos, algunas especies logran colocar hasta el 83% de huevos en otros sustratos como debajo la vaina o en la hojarasca (Rodríguez et al. 2003). Esta variación es obviamente relevante, al momento de implementar tácticas de manejo tendientes a disminuir el impacto del insecto en los diversos hábitats.
Los parámetros biológicos, así como los principales estadísticos vitales de una población de un insecto plaga, estimados con base en tablas de vida desarrolladas en condiciones de laboratorio, se constituyen en una herramienta básica para la implementación de estrategias de control (Carey 1993; Southwood 1978). Estas estimaciones han sido empleadas en estudios tendientes a evaluar resistencia de plantas (Trichilo y Leigh 1985) y además como un patrón para la selección de enemigos naturales (Janssen & Sabelis 1992). Las tablas de vida suministran información válida para la comprensión de la dinámica poblacional de una especie en particular (Sharov 1996; Carey 1993); éstas resumen la información esencial de una población con respecto a la tasa de mortalidad, supervivencia y esperanza de vida de la especie (Silveira et al. 1976). Según Carey (1993), las tablas de vida pueden ser completas o abreviadas; en una tabla de vida completa las funciones se computan para cada día de la vida.
Con base en lo anterior se planteó el presente trabajo con el propósito de establecer una tabla de vida específica para el caso de Zulia carbonaria (Lallemand) como un complemento a los estudios biológicos desarrollados en el CIAT para la misma especie (Rodríguez et al. 2002; Arango y Calderón 1981).
Zulia carbonaria es una de las 16 especies de salivazo de los pastos reportados hasta la fecha en Colombia asociadas con gramíneas forrajeras (Peck et al. 2004; Peck 2001). Actualmente esta especie se encuentra registrada en seis departamentos de Colombia asociada con Andropogon gayanus Kunth, Brachiaria brizanta (A. Rich.) Stapf, Brachiaria decumbens (Stapf), Brachiaria dictyoneura (Fig. & De Not.) Stapf, Brachiaria humidicola Stapf, Brachiaria plantaginea (Link) Hitchc., Brachiaria sp., Cynodon sp., Oryza sativa L., Pennisetum clandestinum Hochst. ex Chiov., y Saccharum officinarum L. Z. carbonaria es un insecto chupador que extrae su alimento del xilema (Fewkes 1969). Al igual que los otros miembros de la familia Cercopidae las ninfas se caracterizan principalmente por la construcción de una masa de espuma. Esta masa sirve de defensa contra enemigos naturales y protección contra microclima adverso (Whittaker 1970). Los adultos no poseen la capacidad de producir la masa de espuma; emplean como mecanismo de defensa colores aposematicos, y apoyados por una defensa secundaria denominada autohemorragia (Peck 2000).
Debido a la carencia de estudios relacionados con parámetros poblacionales en especies de salivazo, y para evaluar protocolos y empezar a establecer estos fundamentos para este grupo en Colombia, se escogió trabajar con una especie modelo Z. carbonaria. La biología básica y fluctuación poblacional de esta especie han sido descritas en una serie de estudios conducidos en el CIAT en el valle geográfico del Río Cauca (Castro et al. 2005; Rodríguez et al. 2002; Peck 2001; CIAT 1999; Castro et al. 1999). Esta información ofrece nuevas bases para el establecimiento de tácticas de manejo tendientes a disminuir el impacto del insecto en campo y adicionalmente, como una medida de control de calidad para las colonias implementadas para llevar acabo estudios de resistencia por el CIAT.
Materiales y Métodos
El trabajo se realizó durante el primer semestre del 2002 en condiciones de laboratorio y casa de malla (temperatura mínima promedio de 19,5°C, máxima promedio de 29,5°C, y humedad relativa mínima del 56,3%), en las instalaciones del Centro Internacional de Agricultura Tropical (CIAT), localizado a 3°31′N y 76°22'W, altura de 965 msnm y precipitación anual de 1.000 mm. La colonia para estos estudios se estableció a partir de hembras recolectadas en el Mpio. Santander de Quilichao, Departamento del Cauca a 1.060 msnm. Brachiaria ruziziensis CIAT 654 fue el material vegetal usado en cada fase de los estudios.
Mediante la observación diaria de los huevos se pudo determinar la emergencia de las ninfas (<12 horas de edad). Se retiraron de las cajas reticuladas para llevarlas al invernadero, donde fueron depositadas en potes plantados con el hospedero. Los potes fueron cubiertos con una tapa provista de un orificio para darle paso al material vegetal y contener a la ninfa. Esta metodología permitió determinar la supervivencia y mortalidad para cada uno de los instares ninfales (Rodríguez et al. 2002; Lapointe et al. 1992).
Basados en una cohorte inicial de 118 huevos se calcularon los parámetros de la tabla de esperanza de vida: (ax) número de individuos al inicio de cada grupo de edad, (lx) proporción de la cohorte que sobrevive a cada grupo de edad, (dx) número de individuos muertos durante cada grupo de edad, (qx) tasa de mortalidad específica por etapa de desarrollo y (kx) poder de mortalidad (Begon et al. 1990). Los datos demográficos se resumieron en una tabla de esperanza de vida, con los valores promedios de los datos observados. Para la configuración de la tabla de esperanza de vida de Z. carbonaria se empleó una estructura utilizada en otros estudios, pero adicionalmente se incluyó el poder de mortalidad (kx) como una medida más de la mortalidad para los estados inmaduros que a diferencia del (qx) pueden ser sumados y permitir comparaciones entre los distintos valores en las etapas de desarrollo del mismo ciclo (Manrique et al. 1998; Dent y Walton 1997; Sharov 1996).
(A) Hembras de Z. carbonaria confinadas para la obtención de huevos. (B) Montaje para la observación del desarrollo de los huevos (cohorte inicial).
Los cálculos se basaron en los valores promedios para una cohorte inicial de 30 hembras. Se determinó la fecundidad y la fertilidad de las hembras basados en el número total de huevos colocados por la cohorte. De igual forma se determinó la relación de sexos apoyados en evaluaciones y observaciones en estudios anteriores realizados por el CIAT. Con los valores promedios de la longevidad de las hembras, la tasa de oviposición y la proporción de sexos, se construyó la tabla de fertilidad basado en lo descrito por Birch (1948). Construida la tabla de fertilidad, se determinaron los siguientes parámetros demográficos. El tiempo generacional (T), fue entendido como el período medio entre el nacimiento de los padres y el nacimiento de los descendientes. La tasa neta de reproducción (Ro), representó el número promedio de los descendientes que una hembra dejó en una generación, resumiendo la supervivencia y la fecundidad. La tasa intrínseca de crecimiento natural de la población, o capacidad innata de crecimiento (r m ), fue entendida como el desarrollo potencial de una población de acuerdo con la cohorte inicial. Adicionalmente se calculó la tasa finita de multiplicación lambda (λ).

(A) Montaje empleado para establecer la mortalidad y supervivencia de ninfas de Z. carbonaria. (B) Montaje para establecer la fecundidad de las hembras Z. carbonaria.
Resultados
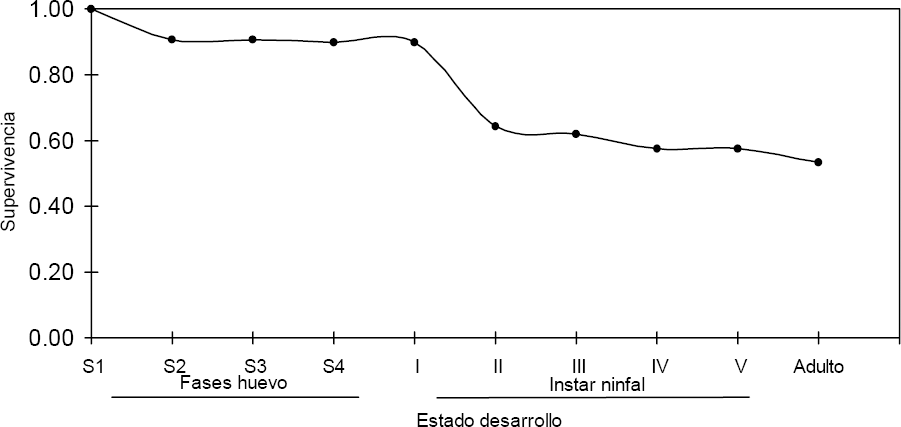
Curva de supervivencia de Z. carbonaria alimentadas sobre el hospedero B. ruziziensis CIAT 654 bajo condiciones de estudio.
Tabla de vida para valores promedios de los estados inmaduros de Z. carbonaria sobre B. ruziziensis.
ax = número de individuos al inicio de cada grupo de edad, lx = proporción de la cohorte que sobrevive a cada grupo de edad, dx = número de individuos muertos durante cada grupo de edad, qx = tasa de mortalidad, kx = poder de mortalidad
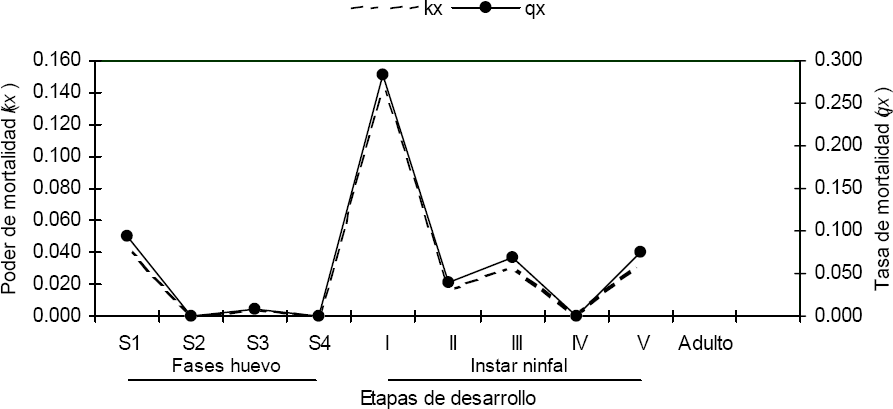
Tasa de mortalidad (qx=dx/x) y poder de mortalidad (kx=lnx-lnx+1) para los estados inmaduros de Z. carbonaria bajo condiciones de estudio.
Discusión
La tabla de vida de Z. carbonaria, exhibió la fase ninfal como la más susceptible en términos de supervivencia. Esta fase de desarrollo presentó 3,6 veces más mortalidad por grupo de edad (dx) que la fase de huevo. Para Z. carbonaria, la tasa de supervivencia (lx) fue 1,4 veces mayor en la fase de huevo, siendo la fase S1 la más susceptible con 11,6 veces más mortalidad por grupo de edad (dx) que la fase S3. El poder de mortalidad (kx) establecido para Z. carbonaria bajo las condiciones del estudio fue 2,1 veces menor al valor de la tasa de mortalidad. Esto confirma la utilidad del poder de mortalidad (kx) al ser un valor aditivo entre estados de desarrollo en un mismo ciclo de vida.
La longevidad exhibida por las hembras de Z. carbonaria, superó en 13,8 días la longevidad reportada para las hembras de esta especie en estudios biológicos anteriores (Rodríguez et al. 2002). Esta diferencia pone de manifiesto un posible efecto del ajuste en los métodos, representado por el suministro de material vegetal fresco cada tres días.
La fertilidad exhibida por las hembras de Z. carbonaria durante este estudio, fue igual a la observada para hembras traídas directamente de campo (Rodríguez observación personal). Esto confirma que los métodos implementados para el establecimiento de la tabla de vida de Z. carbonaria no tienen ningún efecto negativo sobre la primera generación, aunque se deba retomar la información para determinar el posible efecto sobre generaciones sucesivas.
La tasa neta reproductiva (Ro) conocida como tasa de reemplazo, muestra para las condiciones del ensayo, que Z. carbonaria multiplicó su población 33,6 veces en un tiempo generacional (T) de 67,4 días. El valor que pueda exhibir (Ro) en generaciones sucesivas, nos permite determinar la calidad de las condiciones artificiales empleadas para la cría de insectos, teniendo en cuenta que este parámetro es una característica innata de la población (García 2002; Nascimento 1996; Parra et al. 1995; Magrini 1993).
El tiempo generacional (T) para Z. carbonaria, fue muy parecido al establecido en condiciones de campo y laboratorio de 66,0 y 69,6 días de adulto-adulto, respectivamente (Castro et al. 2005; Rodríguez et al. 2002). A diferencia de Ro, la variación en los valores del tiempo generacional no son indicadores de la calidad de las condiciones de cría, debido a las variaciones que este parámetro puede sufrir por condiciones abióticas. El valor positivo de la tasa intrínseca de crecimiento (r m ) indica un aumento en la población de Z. carbonaria bajo las condiciones del ensayo, confirmándose que estas son adecuadas para el mantenimiento de una cría de Z. carbonaria. Adicionalmente, la tasa finita de multiplicación (λ) está relacionada directamente con la tasa intrínseca de crecimiento (r m ), y se interpreta como el número de individuos que se adicionan a una población por individuo y por unidad de tiempo (individuo/hembra/día).
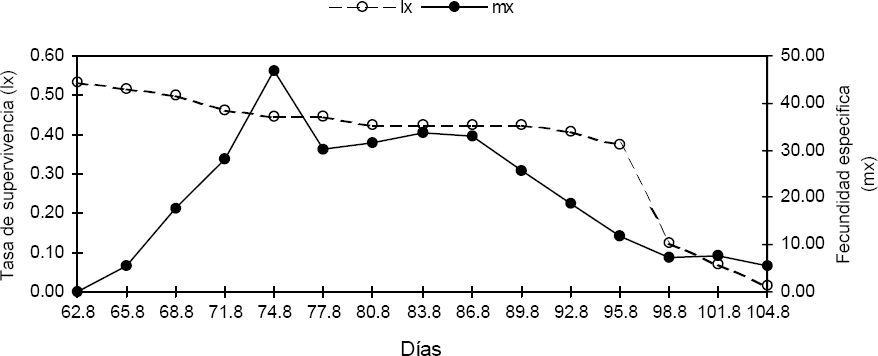
Curva de supervivencia de hembras (lx) y fecundidad específica (mx) de Z. carbonaria, alimentadas sobre B. ruziziensis CIAT 654 bajo condiciones de estudio.
El valor (Ro) para Z. carbonaria bajo las condiciones del presente estudio, fue 3,9 veces menor a lo reportado para Mahanarva fimbriolata (García 2002). De igual forma, la tasa finita de multiplicación (λ) fue 1,3% veces menor que la reportada para M. fimbriolata (García 2002). Al comparar los resultados obtenidos con Z. carbonaria con estudios preliminares, se logra establecer que la supervivencia de Z. carbonaria huevo-adulto fue 1,5 veces menor a la determinada para Prosapia simulans (Walker) bajo las mismas condiciones de estudio (CIAT 2003). El poder de mortalidad (kx) exhibido por Z. carbonaria, fue 2,5 veces superior al establecido para P. simulans. Además, el poder de mortalidad (kx) en Z. carbonaria fue superior para el estado ninfal, en contraposición a lo observado para el caso de P. simulans donde este valor fue mayor para el estado de huevo (CIAT 2003).
La fertilidad de Z. carbonaria determinada durante el estudio, fue similar a la de otras especies de salivazo estudiadas en Colombia como Mahanarva andigena y P. simulans con el 94,3 y 95,1%, respectivamente (CIAT 2002).
Conclusiones
Comparado con la fase de huevo y adulto, la fase ninfal sufre la mayor mortalidad, siendo el instar I el más susceptible. Subraya lo apropiado de tácticas culturales dirigidas a disminuir la calidad del hábitat para el desarrollo del insecto en el cultivo, en particular al momento de la eclosión de los huevos cuando el instar I establece en su sitio de alimentación.
Las variables evaluadas - fecundidad, fertilidad, tiempo generacional, tasa neta reproductiva, tasa intrínseca de crecimiento y tasa finita de multiplicación- son parámetros fundamentales para cuantificar la biología de Zulia carbonaria mediante una tabla de esperanza de vida y una tabla de fertilidad. Los valores obtenidos son comparables con los datos obtenidos en otros estudios de campo y cría. El éxito del protocolo abre perspectivas para conseguir y comparar datos cuantitativos entre diferentes especies de cercópidos a través de agroecosistemas contrastantes.
La construcción de una tabla de vida se constituye en una herramienta apropiada para evaluar la calidad de las crías de Z. carbonaria, tanto como otras especies plaga de salivazo, bajo condiciones semicontroladas.
Footnotes
Agradecimientos
Se agradece a Anuar Morales, Ulises Castro, Rosalba Tobón y Oscar Yela, por el apoyo para el desarrollo de este trabajo. Este trabajo se realizó gracias al apoyo financiero de la Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República.