
Abstract
The Pampa biome in southern Brazil has grassland areas with high biodiversity. Given the fast advance of
Introduction
The Pampa biome has vast areas of grasslands, covering the southern half of Rio Grande do Sul state, already the southernmost state of Brazil, filling approximately 176.496 km2 (IBGE 2004). Southern grasslands include areas with a high diversity of both plants and animals, with a long history of low impact pasture management under extensive cattle farming. However, its conservation has been threatened by the increased degradation caused by the inadequate use of exotic species, especially grasses, and especially by a recent economic interest in agriculture and silviculture (Pillar
Land use (agriculture, silviculture and pasture) can degrade natural environments, reducing biodiversity, mainly through loss of habitat (Wilcox and Murphy 1985; Primack and Rodrigues 2002). The establishment of exotic species silviculture has been a widely debated subject in southern Brazil, an activity that is known to generate strong negative impact in the original environments (Pillar
Many soil organisms, as most invertebrates, are directly affected by land use mode. Arthropods are an important component of the natural diversity in any habitat (May 1986), including native grasslands and silviculture. On the soil of both environments a complex system of organic matter cycling is established and leaf litter acts as food for a variety of arthropods, and as shelter for others, composing a particular food chain of detritivores and predators (Höfer
Few literature sources have studied the edaphic fauna of the eucalypt monoculture, comparing it to other environments, and mostly such comparisons address native forests (Ferreira and Marques 1998; Pellens and Garay 2000; Moço
Evaluations of diversity, richness and invertebrate species composition, especially arthropods, can help understand the dynamics of these introduced habitats. It is thus fundamental for wide scale planning of environmental management and conservation of all habitats composing a landscape. Given the advance of eucalypt silviculture activities over the grassland ecosystem of the Pampa biome, we aimed to evaluate the abundance, species richness, and foraging guild proportions and species composition for soil spiders comparing both environments.
Material and Methods
Study areas.
Sampling took place in three municipalities of the southern region of Rio Grande do Sul state of Brazil, within the Pampa biome. Five farms were sampled in the summer of 2008: in Cerrito municipality, Nossa Senhora do Guadalupe (NG) farm (31°77’56.57”S, 52°64’53.32”W; 208 ha of planted area; 440 ha of total area, sampled in 03/01/2008) and Pitangueiras (PI) farm (31°79’38.57”S, 52°53'50.51”W; 101 ha of planted area; 230 ha of total area, sampled in 24/01/2008); in Capão do Leão municipality, Ouro Verde (OV) farm (31°57’55.40'S, 52°51'42.73”W; 125 ha of planted area; 297 ha of total area, sampled in 15/01/2008); in Piratini municipality, Santa Izabel (SI) farm (31°56’50.95”S, 52°88'20.48”W; 145 ha of planted area; 330 ha of total area, sampled in 14/02/2008) and Santa Maria (SM) farm (31°56’58.01”S, 53°17’06.13”W; sampled in 21/02/2008). Silviculture was based on Eucalyptus saligna Smith planted in 2006. Grassland areas are managed lightly by releasing cattle infrequently for use as pasture.
Sampling method and design.
For each farm a native grassland and eucalypt plantation area were designated. From each area 25 samples were taken, distributed along a linear transect. To avoid edge effects, samples were at least 50 m far from the limits between the areas, and to guarantee a degree of independence between samples, there were also at least 50 m between consecutive sampling areas. For each sample all leaf litter and a small superficial fraction of the soil was taken from an area of 1 m2. The material was put through a field sieve, with the content transferred to closed nylon bags. These were taken to the lab where bag content was placed in Winkler extractors, where it stayed for 72h to collect the animals. Spiders were identified in the Laboratório de Aracnologia and deposited in the spider collection (curator: E. H. Buckup) of Museu de Ciências Naturais of Fundação Zoobotânica do Rio Grande do Sul, in Porto Alegre, Brazil.
Data analysis.
Diversity variables were compared using PASt (Paleontological Statistics 1.97, Hammer and Harper 2009), with environment (eucalypt plantation or native grassland) and site (farms) as factors. Alpha diversity (abundance, richness and equability) was compared at the family and species levels using two-way ANOVAs. To illustrate spider species composition comparisons two ordenations were plotted (Non-metric MultiDimensional Scaling - nMDS) using a qualitative similarity index (Simpson) and a quantitative one (Morisita). To test for statistical differences among spider assemblage composition for the above similarity indexes, we applied two one-way ANOSIM (Analysis of Similarities) with Bonferroni correction, one for each factor. A SIMPER Analysis (Percentage Similarity) was employed to rank species contributing more for dissimilarities among environments and sites (Clarke and Warwick 1994). Species accumulation curves and analytical species richness estimator Chao1 were calculated to verify sampling sufficiency, as suggested by Toti
Guild classification was based on Uetz
Results
Overall 1.301 spiders were found, mostly immatures (76.4%). Among adults, females were more common (77.5%) than males (22.5%) (a sex ratio of 1:3.5). A higher absolute abundance characterised the eucalypt plantation with 816 individuals, whilst grassland resulted in 485 spiders. Thus, spider densities were 6.53 (± 1.010 s.e.) individuals.m−2 in eucalypt, and 3.88 individuals.m−2 (± 0.731 s.e.) in grassland. Out of 125 samples, seven did not record spiders in eucalypts and 21 in the grassland; the highest number of spiders in a sample was 36 individuals for eucalypt and 27 for grassland.
Twenty-four spider families were captured, of which eleven were represented only by juveniles; the most abundant were: Salticidae (N = 276), Linyphiidae (230), Gnaphosidae (185), Theridiidae (124) and Lycosidae (116); the least abundant were Nemesiidae, Oonopidae, Senoculidae and Sparassidae, all with singletons. Of the five most abundant families, four were more abundant in eucalypt, the exception being Lycosidae, found more in the grasslands (Table 1). Nineteen families occurred in eucalypt and 21 in grassland. Three were exclusive to eucalypt, five to grassland. Six families were recorded from all sites and both environments (Table 1).
Analyses at the familial level (i.e. including young spiders) showed average abundance to differ significantly between environments, with more individuals found in eucalypt plantations (F1,4 = 13.83, P = 0.020). Surprisingly, differences in family density among sites are marginally significant (F4,4 = 6.00, P = 0.055; Fig. 1), indicating a possible founder effect. No differences in evenness among families were detected between environments or among sites.
Number of individuals (adults and young) per family and guilds found in the two environments (eucalypt plantations and grassland), in the Pampa biome, southern Brazil (ORB, Orb weavers; SPW, Space web sheet builders; HAS, Hunting - Ambushers/stalkers; HRU, Hunting - Runners; NG, Nossa Senhora do Guadalupe farm; OV, Ouro Verde farm; PI, Pitangueiras farm; SI, Santa Izabel farm; SM, Santa Maria farm).
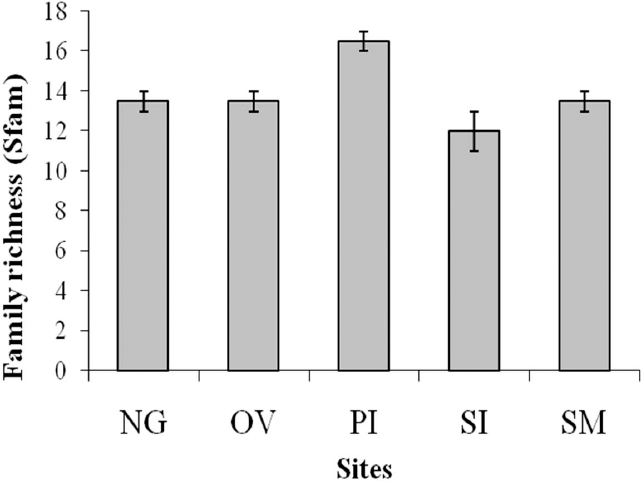
Family spiders richness (adults and young, ± s.e.) in the five farms of the Pampa biome, southern Brazil (NG, Nossa Senhora do Guadalupe farm; OV, Ouro Verde farm; PI, Pitangueiras farm; SI, Santa Izabel farm; SM, Santa Maria farm). Differences among farms are marginally significant (p < 0.055).
For adult spiders, 51 morphospecies were determined, the most abundant being
Each environment totalled 35 morphospecies, and thus no sample-based rarefaction was employed to distinguish environments. However, the Chao1 estimator indicated an expected 40.63 species for eucalypt, which would mean 86.1% of the spider fauna have been sampled for that environment. Figures for grassland were somewhat different with 47 species expected, meaning 74.5% of the fauna sampled. However, the estimated values do not differ significantly between environments (considering confidence intervals).
Adult abundance was marginally significant for environment (F1,4 = 7.50; P=0.052), with eucalypt areas having more spiders (Fig. 2A); no differences were found among sites. Species density (Fig. 2B) did not differ between either environments or sites, but evenness was significant for environment (F1,4 = 14.65; P = 0.019), being higher in grassland (Fig. 2C). Consequently, abundance distribution curves showed higher dominance by a few species in eucalypt with more rare species in grasslands as well. Curves for both environments significantly approximated the log-series model (eucalypt: α = 12.42,
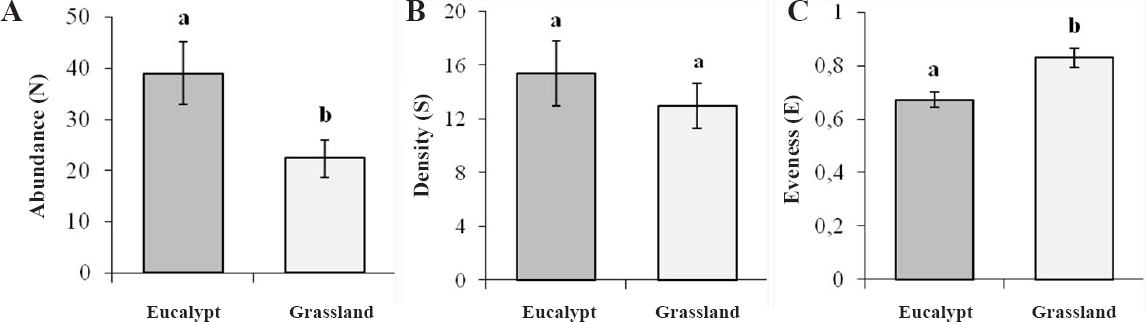
Adult spider diversity in two environments (eucalypt plantation and grassland) in the Pampa biome, southern Brazil. (A) Abundance. (B) Species Density. (C) Evenness (letters indicate significant differences, P < 0.05).
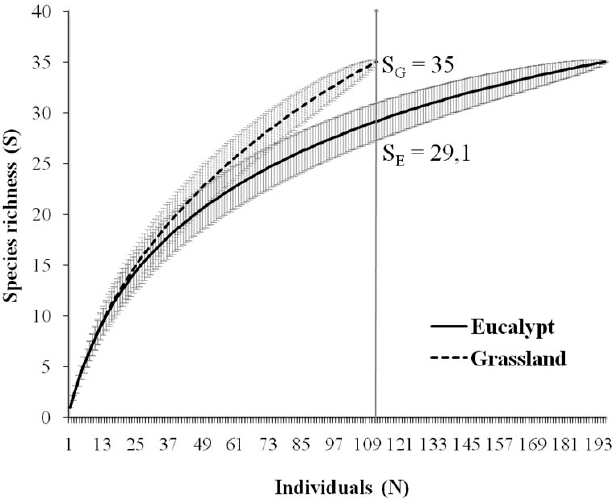
Individual-based rarefaction curves for the two environments (eucalypt and grassland) in the Pampa biome, southern Brazil (SG, grassland richness; SE, eucalypt richness; numbers indicate richness at the comparison point).
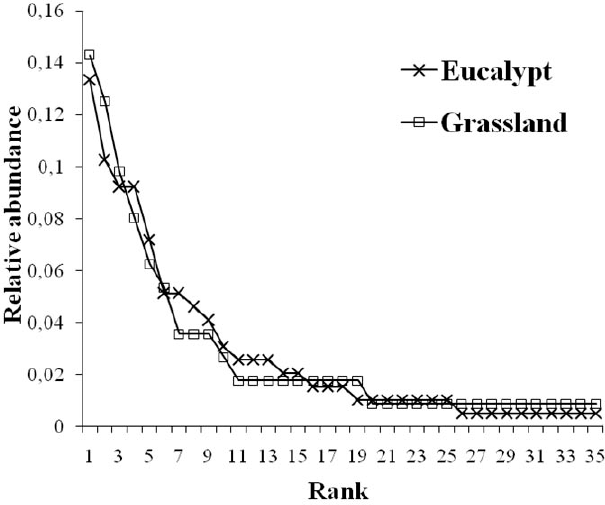
Species abundance distribution in two environments (eucalypt and grassland) in the Pampa biome, southern Brazil (relative abundances shown to allow visual comparison).
More than 37% of the species were common to both environments and the two most common species on each environment were the same. Sixteen species were exclusive to each environment, probably due to the large number of singletons and doubletons in the sample. This amount of exclusivity sets the two environments apart and thus differences in species composition are significant between the two environments for the qualitative index (Simpson, ANOSIM:
Number of individuals (adults) in spider morphospecies and diversity values for the two environments (eucalypt plantation and grassland) in the Pampa biome, southern Brazil (NG, Nossa Senhora do Guadalupe farm; OV, Ouro Verde farm; PI, Pitangueiras farm; SI, Santa Izabel farm; SM, Santa Maria farm).
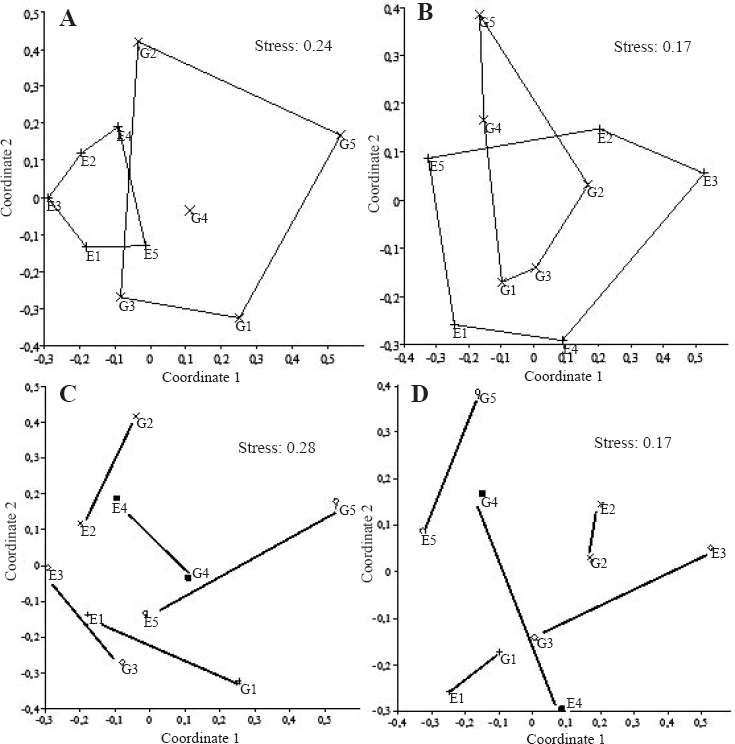
Ordination of spider faunal composition for environments and farms - Non-Metric MultiDimensional Scaling (nMDS, using qualitative and quantitative similarity indexes) - in the Pampa biome, southern Brazil.
The most abundant guild was the running hunters (N = 743), followed by irregular web builders (454) ambush hunters (77) and especially orbicular web builders (27) were relatively rare (Fig. 6). No differences were found in the proportion represented by each guild, either between environments (two-way ANOVA, interaction term: F3,32 = 1.547 p = 0.221) or sites (two-way ANOVA, interaction term: F12,20 = 0.789 p = 0.656).
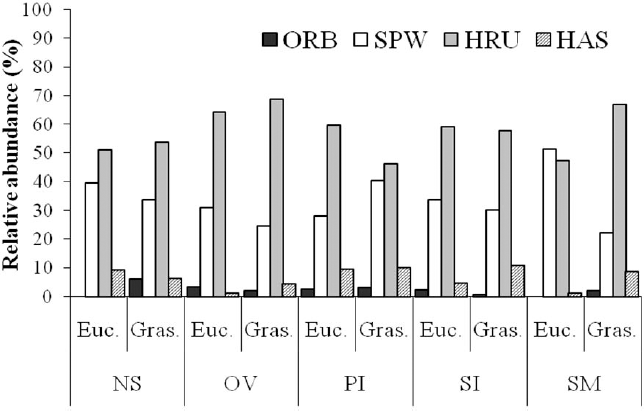
Spider feeding guilds for the two environments (eucalypt plantation and grassland) on each farm in the Pampa biome, southern Brazil (ORB, Orb weavers; SPW, Space web sheet builders; HAS, Hunting - Ambushers/stalkers; HRU, Hunting - Runners; NG, Nossa Senhora do Guadalupe farm; OV, Ouro Verde farm; PI, Pitangueiras farm; SI, Santa Izabel farm; SM, Santa Maria farm).
SIMPER analysis for the first ten species contributing more for dissimilarities between the two environments (eucalypt plantation and grassland) in the Pampa biome, southern Brazil.
species percentage contribution to dissimilarity;
cumulative dissimilarity between environments;
mean species abundance per transect on each environment.
Discussion
This is the first record of the spider fauna comparing eucalypt silviculture and grasslands in the Pampa biome of Brazil. Most of the studies compare eucalypt plantation to native forests; among such comparisons, generalizing for soil arthropod fauna, a lower species richness and/or abundance is found in silviculture (Ferreira and Marques 1998; Pellens and Garay 2000; Moço
The above sources differ somewhat from what we found in southern Rio Grande do Sul; a higher abundance was found in a monoculture (eucalypt) and species density was identical between eucalypt and grassland, although richness per se was higher in the latter. These two environments differ in other aspects as well, with grasslands showing higher heterogeneity in their spider assemblages and species composition quantitatively distinct. Higher structural vegetation complexity is known to lead to higher spider diversity, thanks to a larger number of microhabitats for web building and shelter (Souza 2007; Rypstra
In the eucalypt plantation the amount of litter can be higher due to a larger plant biomass. Litter can soften extreme abiotic factors (Uetz 1979; Höfer
At the litter level, the grassland environment also suffers stronger disturbances, as higher light intensity, higher wind speeds and direct rainfall, among others. Some or all of these abiotic factors could lead to a lower abundance in the grassland, either through stronger hygrothermal stress to spiders, damages to webs, or both. These effects in turn can influence not only spider survival and reproduction but also important behaviors as those involved in environment choice (Wise 1993).
Family composition showed dissimilarity with higher richness and more exclusive families recorded in the grassland. Rinaldi (2005) studied a eucalypt plantation in Botucatu, São Paulo state, Brazil, recording 17 families at different strata, finding only seven for soil; here we report 19. Six families were shared between theirs and our inventory, Caponiidae was exclusive to the Botucatu study and six were exclusive to southern Rio Grande do Sul. No Mygalomorphae was recorded for eucalypt silviculture in either inventory, but we recorded family Nemesiidae for grassland.
Comparing the araneofauna in this study to other work within the Pampa biome is all but impossible given the absence of publications with the same aim. In Rodrigues
Some species are worth mentioning for their habitat use.
Spiders are usually among the first organisms to occupy altered or recently formed habitats, actively participating in community succession processes (Uetz
Although it is known that distinct environments and microenvironments are selective in terms of spider hunting guilds as well as families and species (Uetz 1979; Uetz
Even a rapid spider diversity inventory can produce information on the main ecological aspects of such an assemblage. Data presented here can help direct future studies focusing on the changes brought to the grassland environment and the whole Pampa biome given the strong efforts to occupy these areas with exotic silviculture. Long term studies, especially, may permit solving the puzzles found here, to better understand the relationship between native and introduced environments, perhaps to help minimize the impacts brought by human disturbance.
Footnotes
Acknowledgements
To Erica H. Buckup (curator of the Araneae collection) and Maria A. L. Marques for access to spider material and bibliography and the direction of MCN by permitting the first author to develop part of this work at it dependences. This study was supported by CNPq (process 140586/2007-5 for E. N. L. Rodrigues from Programa de Pós-Graduação em Biologia Animal da Universidade Federal do Rio Grande do Sul).