
Abstract
Summary
The spider diversity in the forest understory was studied from a premontane forest of the basal slope on the cordillera oriental, in Toquiza's region, Medina's county, Cundinamarca. The main purpose was to describe the diversity, distribution and composition of spider communities on the remaining layer in interior-edge fragments from a premontane forest. We collected specimens using different methods at defined distances form the edge. The results showed a total 409 specimens, 86 morph species and 31 families included in two suborders. The estimators Chaol and Michaelis-Menten showed that the number of species ranged between 100 and 120. These species correspond to 84,21% and the 73,11% of the total observed, from which we concluded that the sampling was effective. The most effective method of capture was the pitfall traps. The most abundant families were Zodariidae, Ctenidae, Araneidae, Heteropodidae, Salticidae, Pholcidae and Liocranidae. Araneidae showed the highest number of morph species (15); the others showed values comparatively low. The open area and the forest interior evaluated showed significant differences in the distribution of individuals within stations; most of the species were found in the forest interior. Zodariidae, Ctenidae, Araneidae and Salticidae showed one tendency to the double-distribution (in both forest interior and open areas). The Hill's Numbers in the evaluated forest showed an increase in the spiders diversity within the interior stations, although the area outside the forest (a 30 meters fragment) did not have that tendency. This could be associated with the minor influence of the factors that define the edge effect; consequently, our study shows a remarkable edge effect and a reduction of the spider diversity in the external stations located on the edge and the 60 external meters from the transect.
Introducción
La diversidad biológica, una propiedad de los sistemas vivientes, está reducida hoy por acciones humanas que transfor- man sus paisajes y sus hábitats. En Co- lombia, esta reducción de la diversidad ha llegado a límites sin precedentes, con- llevando a la deforestación y reducción de los bosques andinos y a la destrucción de los ecosistemas naturales (Andrade 1993) generada por la expansión huma- na. Como consecuencia de la presión antrópica que se ejerce en los bosques andinos, los paisajes resultantes que se mantienen están fragmentados, razón por la cual han perdido su continuidad origi- nal y han generado los llamados efectos de borde (Murcia et al. 1993; Murcia 1995); se entiende como borde a una transición abrupta o nula entre dos ecosistemas o unidades de paisaje (Mur- cia 1995).
Los efectos de borde pueden influir sobre el bosque, penetrar a diferentes distancias e incluir la introducción en éstos de espe- cies generalistas, habituadas a las condi- ciones variables del exterior, generando bien sea la extinción local, el reemplazo o el desplazamiento de las especies de bos- que, habituadas a las condiciones del in- terior mucho más homogéneas en cuanto al comportamiento de los factores físicos. Estas especies con excelentes capacidades de dispersión, capaces de invadir y coloni- zar hábitats disturbados, son atraídas a los bordes y pueden penetrar en el centro de hábitats naturales que están conectados con otros bordes que tienen introducción de líneas de claros o de áreas abiertas (Andrews 1990).
Algunas arañas se encuentran dentro del grupo de especies pioneras, otras son pro- pias del interior del bosque (Gunnarsson 1990). Esta doble característica permite emplearlas como modelos para el estudio del efecto que ejerce la cercanía del borde sobre las características de la comunidad al penetrar hacia el interior del bosque (Merret 1978; Webb y Hopkins 1984). En este sentido, el objetivo del presente tra- bajo consistió en describir las variaciones en la comunidad de arañas de estrato ra- sante a lo largo de transectos borde-inte- rior de un bosque subtropical, con el ánimo de observar cómo se comporta la comuni- dad durante esta transición.
Las arañas pertenecen a la Clase Arachni- da, que se encuentra dentro del Phyllum Arthropoda, catalogado como el grupo más diverso y abundante de los ecosis- temas terrestres (Erwin 1991; Kremen et al. 1993). Las arañas de estrato rasante al igual que las tejedoras, cumplen un papel ecológico muy importante en el fun- cionamiento de los ecosistemas, ya que se encargan de regular las poblaciones de insectos e incluso alimentarse de pe- queños vertebrados (Wise 1993; Flórez 1996).
Materiales y Métodos
Esta investigación se realizó en la vereda de Toquiza, ubicada en la vertiente orien- tal de la cordillera oriental a los 4° 30′ 26" N y 0° 44′ 14" E, en jurisdicción del muni- cipio de Medina, departamento de Cundi- namarca, Colombia (Fig. 1). El sitio de estudio está ubicado en un gradiente altitudinal que va desde los 1.000 a los 1.200 msnm. El bosque evaluado perte- nece al bioma higrofítico bosque pluvial subtropical (bp-ST) (Holdridge 1967; Igac 1977).
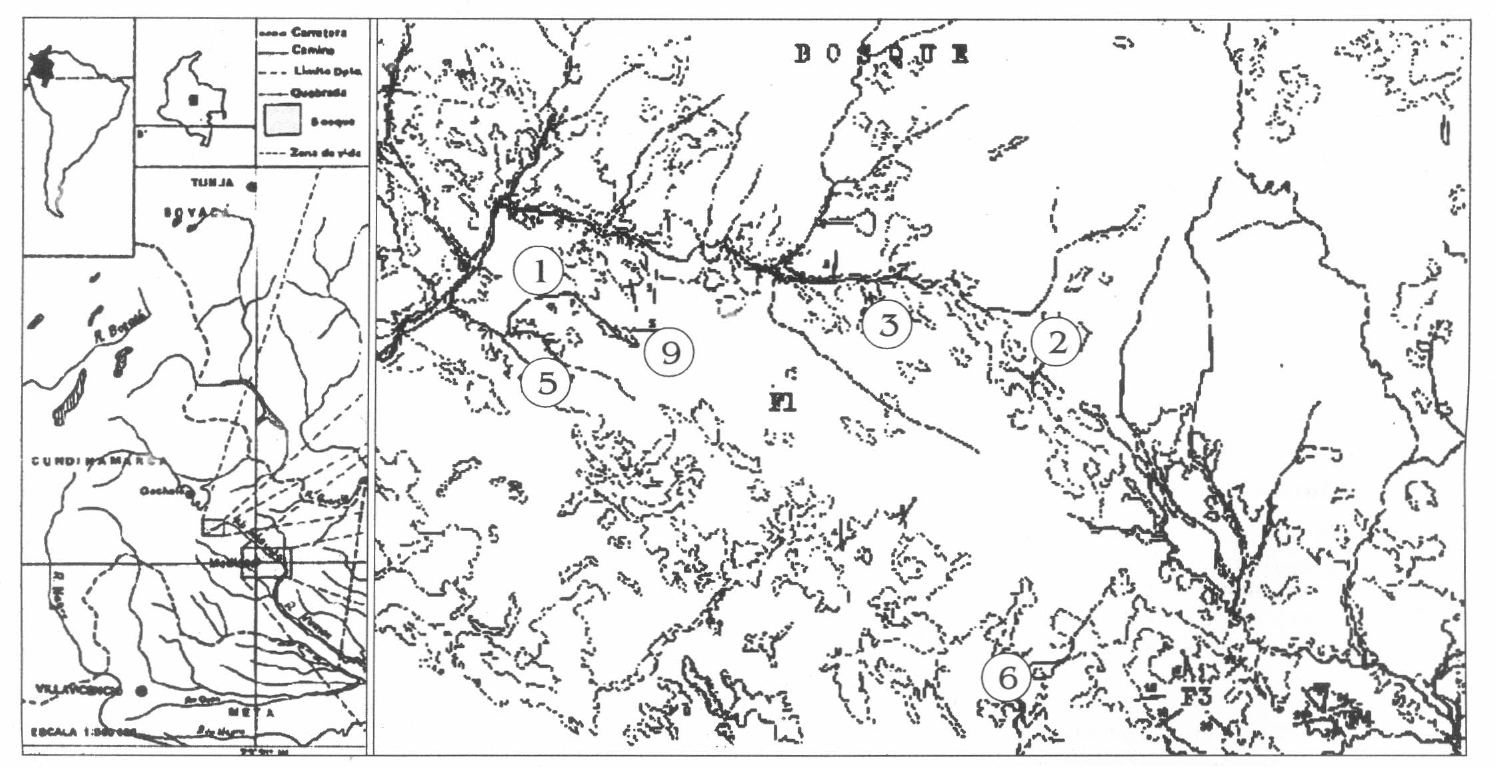
Área de estudio donde se realizó la investigación.
La vereda de Toquiza presenta un relieve escarpado y ondulado, formado por coli- nas y las vegas de los ríos Gazanore y Gazaunta, cuyas pendientes oscilan entre el 35 y el 50%. Presenta una cobertura bastante diversa que incluye bosques in- tervenidos (de los cuales se extrae madera de forma continua) y bosques poco inter- venidos.
La zona de investigación presenta tres es- tratos de vegetación: un estrato arbóreo, uno arbustivo y uno rasante o herbáceo (observación en campo). En el estrato arbóreo predominan plantas de los géne- ros y especies: Alchornia sp. (escobo), Persea coerulea (aguacatillo), Cupania sp. (tostao), Cecropia sp. (yarumo), Ochroma lagopus (balso), Inga edulis (guamo santafereño), Erythrina glauca (cámbulo), Inga densiflora (guamo machete), Loasa sp. (pringamosa), Calliandra sp. (carbone- ro), Clusia sp. (chagualo), Warscewiezia ciccinea (barba de gallo), Tabebuia chrysantha (guayacán rosado), Ficus sp. (caucho), Guadua angustifolia (guadua), Cynerium sagittatum (cañaflecha) y Cedre- la sp. (cedro). El estrato arbustivo está constituido por las siguientes especies: Rapanea sp. (chagualo), Juglans sp. (no- gal), Weinmania sp. (encenillo) y Abatia verbascifolia (duraznillo) (Igac 1995), en tanto que el rasante o herbáceo mantiene a: Begonia spp., Psychotria spp., Palicourea spp., Calathea spp., Heliconia spp., Peperomia sp., Piper spp., Anthurium spp., Xanthosoma sp., Asplundia sp., Cyperus spp., entre otros.
La investigación se desarrolló en tres fases (Fig. 2): La primera, fase preliminar, tuvo una salida con duración de ocho días, en ella se seleccionaron y se ubicaron las zo- nas de muestreo y se establecieron los transectos. En la segunda, fase de campo, se realizaron 3 colectas con salidas men- suales de 20 días en agosto de 1999 y enero - febrero y junio - julio de 2002, con una intensidad horaria comprendida en- tre las 7 y las 17:30 h. La tercera, fase de laboratorio, se llevó a cabo en el Labora- torio de Entomología de la Facultad de Ciencias de la Pontificia Universidad Javeriana en Bogotá, donde se procedió a la preservación de las arañas colecciona- das en campo, a su determinación taxonó- mica y por último a su separación por morfoespecies y a su posterior incorpora- ción en la base de datos. Finalmente, se analizaron los datos y se catalogaron los ejemplares en la colección de arácnidos e insectos del Laboratorio de Entomología de la Facultad de Ciencias de la Pontificia Universidad Javeriana (MUJ) en Bogotá.
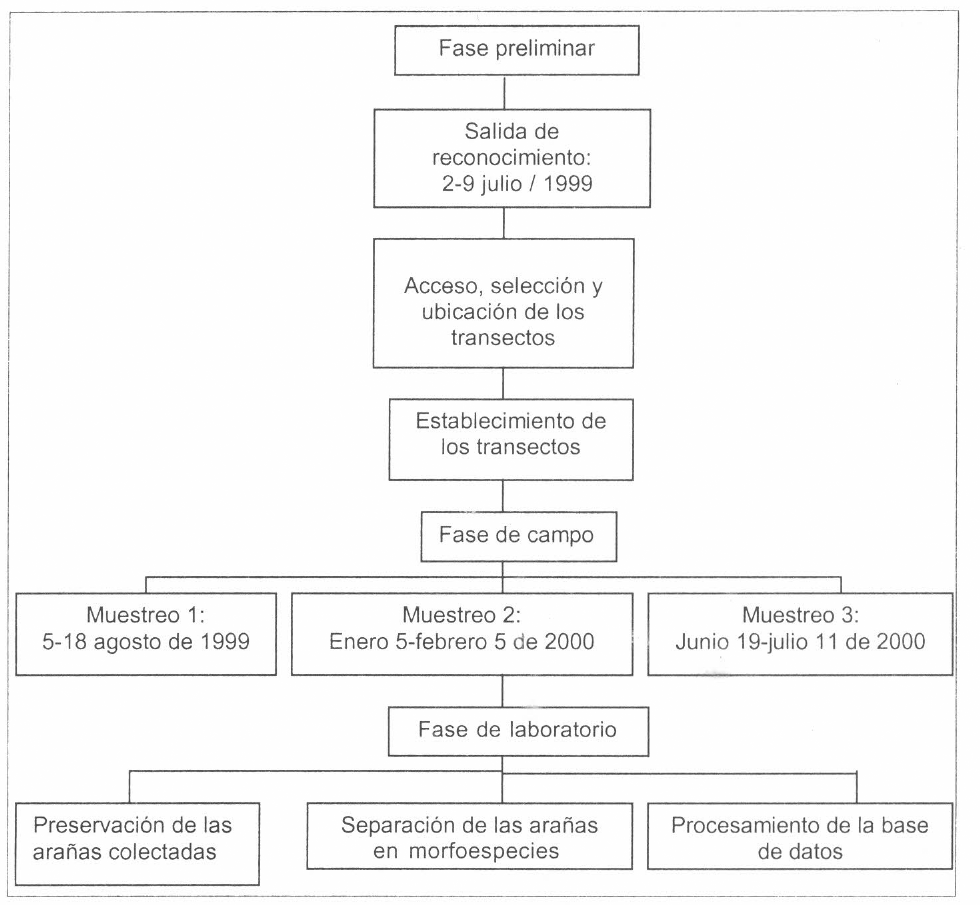
Esquema de las actividades realizadas en este estudio.
Durante la fase preliminar se conoció la zona en general, se seleccionaron rutas de acceso a las zonas de muestreo ubicadas en las veredas de Toquiza y San Antonio. Se establecieron los transectos de cada una de las zonas de muestreo y se realizó un premuestreo para discriminar familias preliminares de arañas presentes en la zona. Se determinó 0,1 ha como unidad física de muestreo. Se utilizaron dos mé- todos de captura: un método directo y uno indirecto. El método de captura directo consistió en la colecta manual de cada uno de los individuos y los métodos de captu- ra indirecta en las trampas pitfall y la reco- lección de hojarasca y su procesamiento mediante el método Winkler.
En la fase de muestreo se demarcaron seis transectos de 210 m perpendiculares al bosque. En cada uno de ellos se delimi- taron ocho estaciones separadas cada 30 m. En cada estación se colocaron 11 tram- pas de caída tipo pitfall, una cada 10 m (Fig. 3), por un intervalo de dos días. Unido a esto se realizaron colectas ma- nuales, teniendo en cuenta los datos res- pectivos como sustrato, altitud y la zona donde se colectó el ejemplar. En cada es- tación se colectaron, al azar, porciones de hojarasca mediante el uso de un cernidor, y luego se depositaron en las trampas selectivas tipo Winkler. Estas trampas se dejaron instaladas por dos días y el con- tenido se vertía posteriormente en fras- cos plásticos para muestras de orina de tamaño estándar conteniendo alcohol al 70%. Cada frasco se rotuló con una eti- queta que contenía los siguientes datos: zona de colecta, el respectivo transecto y número de estación donde se había reco- lectado la hojarasca.
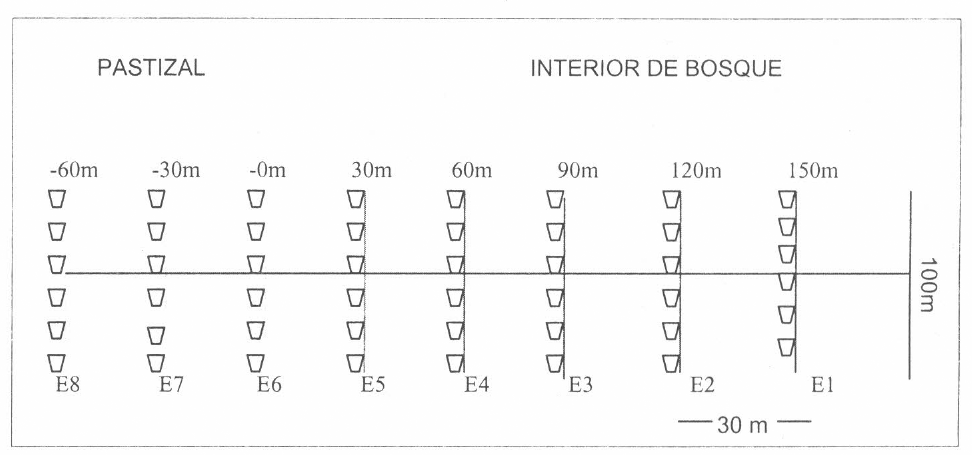
Disposición de los transectos borde interior.
Las colectas manuales se realizaron me- diante el uso de pinzas largas y pequeñas, teniendo cuidado de no dañar el ejemplar capturado. Se anotaron los datos del lu- gar exacto donde se había colectado el ejemplar y posteriormente, los ejemplares se depositaron en frascos plásticos para muestras de orina de tamaño estándar con una etiqueta que reseñaba la zona, el transecto, el sustrato y la estación donde se colectó.
Durante la fase de laboratorio, las arañas colectadas se separaron y se preservaron; posteriormente, se depositaron en frascos de vidrio con alcohol al 70%. En estos fras- cos se colocaron dos etiquetas con datos de colecta. Se realizó la separación de las morfoespecies, se determinaron taxonó- micamente los ejemplares hasta familia, mediante las claves dicotómicas de Flórez (1996), Kaston (1978), Dipenaar-Schoe- man y Jocqué (1997), Raven (1985) y los que se podían hasta el nivel de género o especie, mediante las claves de Levi (1976). Finalmente la información fue pasada a la base de datos.
Resultados
Se colectaron 409 arañas de estrato ra- sante durante la fase de muestreo. Se iden- tificaron 86 morfoespecies pertenecientes a 31 familias y dos subórdenes. Contem- plando todas las capturas en conjunto, las familias más abundantes fueron Zodariidae, Ctenidae, Araneidae, Hetero- podidae, Salticidae, Pholcidae y Liocra- nidae con abundancias entre 53 y 24 individuos (Fig. 4, abundancias respecto de los 409 individuos capturados), el res- to de familias presentaron valores meno- res. Las familias más ricas (asumiendo a las morfoespecies como equivalentes a es- pecies) fueron Araneidae con 15 morfo- especies; Ctenidae, Salticidae y Zodariidae con 6 morfoespecies (Fig. 5), las restan- tes presentaron menos de 6 morfo- especies.
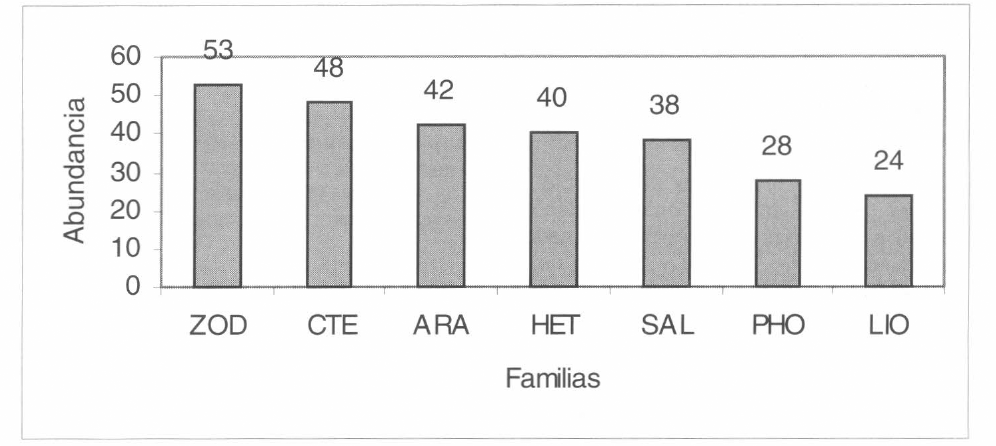
Familias más abundantes en el área de estudio. ZOD: Zodariidae, CTE: Ctenidae, ARA: Araneidae, HET: Heteropodidae, SAL: Salticidae, PHO: Pholcidae y LIO: Liocranidae.
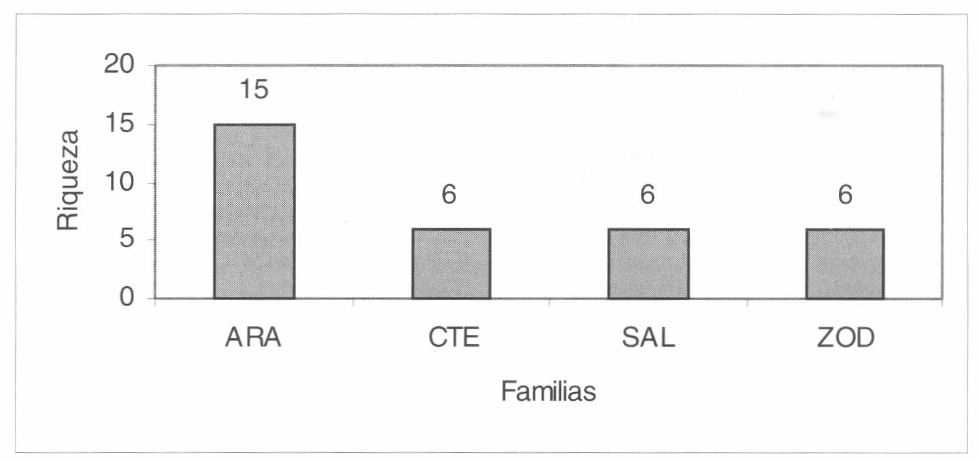
Familias más ricas que se registraron en el área de estudio. ARA: Araneidae, CTE: Ctenidae, SAL: Salticidae y ZOD: Zodariidae.
Las morfoespecies con mayor número de capturas en el área de estudio fueron Zodariidae M1, Araneidae M5, Pampho- beteus sp. 2 (Theraphosidae), Pholcidae M3, Oonopidae M1, Ctenidae M2 y Hetero- podidae M2, con valores de abundancia relativa entre el 5,12 y 3,17% (Fig. 6), las restantes especies presentaron valores inferiores.
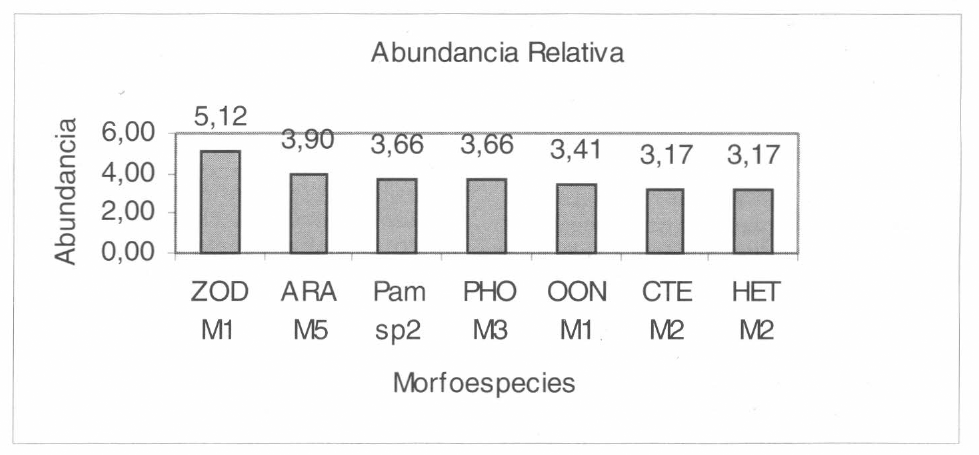
Grupo de morfoespecies más abundantes en los transectos evaluados. ZOD M1: Zodariidae M1, ARA M5: Araneidae M5, Pam sp2: Pamphobeteus sp. 2, PHO M3: Pholcidae M3, OON M1: Oonopidae M1, СТЕ М2: Ctenidae M2 y HET M2: Heteropodidae M2.
Las familias que se capturaron más con el uso de las trampas de caída tipo pitfall fue- ron Araneidae, Zodariidae, Theraphosidae, Salticidae, Liocranidae y Ctenidae (Fig. 7). La captura mayor de la familia Araneidae se observó en la estación 3 de interior, correspondiente a los 90 m. En cuanto a la familia Zodariidae, este método fue muy efectivo, ya que se capturaron ejemplares en todo el interior del bosque. Para la fa- milia Theraphosidae, la captura mayor se registró en la estación 5 (60 m); al igual que en la familia Zodariidae, las capturas por trampas de caída para Salticidae, Liocranidae y Ctenidae, fueron altas y se observaron en todo el interior del bosque (Fig. 7) y en las áreas abiertas (Fig. 8).
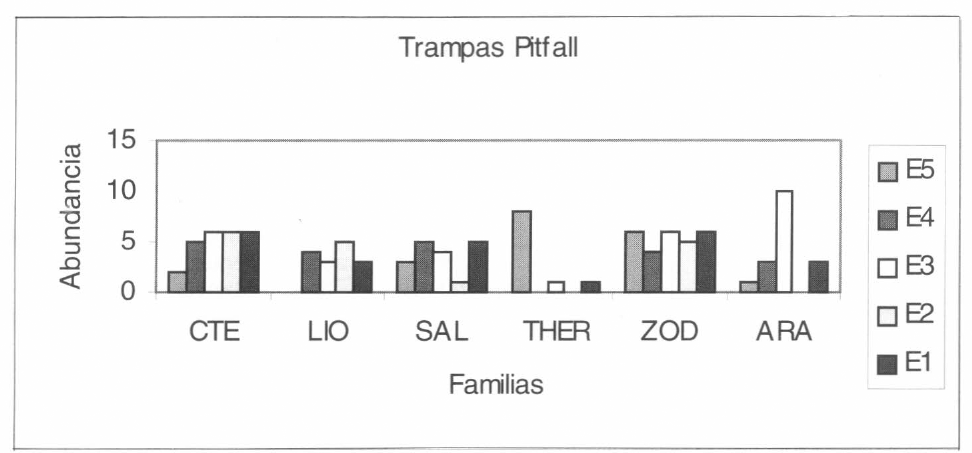
Grupo de familias que se registraron mediante la utilización de las trampas Pitfall en el interior del bosque. CTE: Ctenidae, LIO: Liocranidae, SAL: Salticidae, THER: Theraphosidae, ZOD: Zodariidae y ARA: Araneidae.
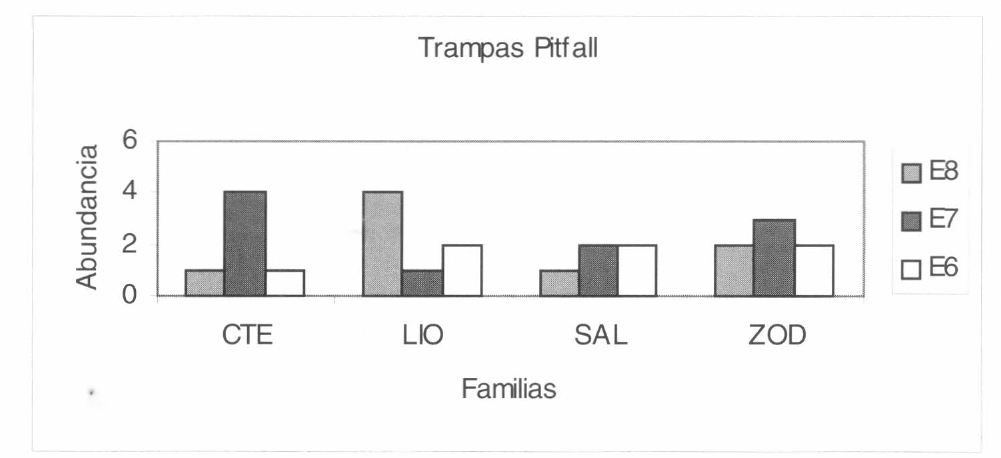
Grupo de familias que se registraron en los pastizales mediante la utilización de las trampas Pitfall. CTE: Ctenidae, LIO: Liocranidae, SAL: Salticidae y ZOD: Zodariidae.
Las familias más representativas mediante la utilización de las trampas Winkler en el interior del bosque fueron Zodariidae, Oonopidae y Hahniidae. Este método mos- tró alta efectividad en el interior del bosque para la familia Zodariidae y Oonopidae. En la familia Hahniidae, se vio una distribu- ción de abundancias en casi todo el interior del bosque. En la familia Heteropodidae se observó que el método sólo fue efectivo en las estaciones 2 y 3 correspondientes a los 120 m y 90 m, respectivamente (Fig. 9) y en los pastizales (Fig. 10).
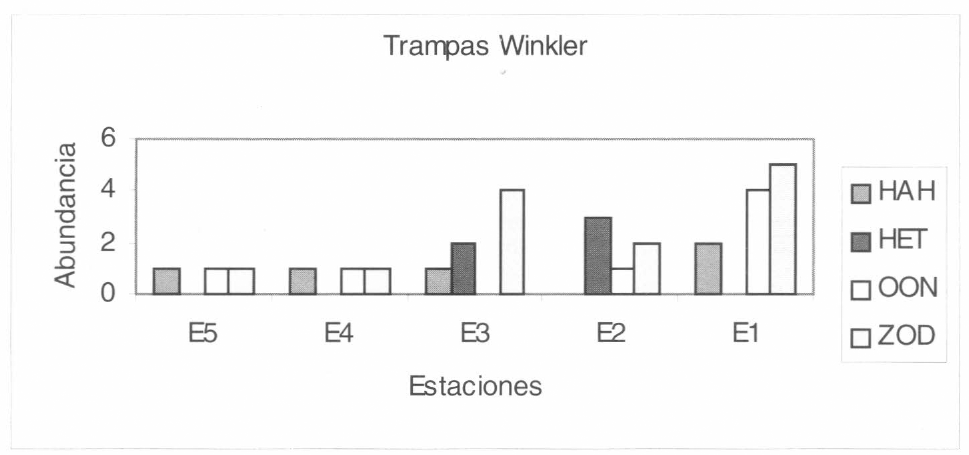
Grupo de familias más frecuentes que se registraron mediante la utilización de las trampas Winkler en el interior del bosque. HAH: Hahniidae, HET: Heteropodidae, OON: Oonopidae y ZOD: Zodariidae.
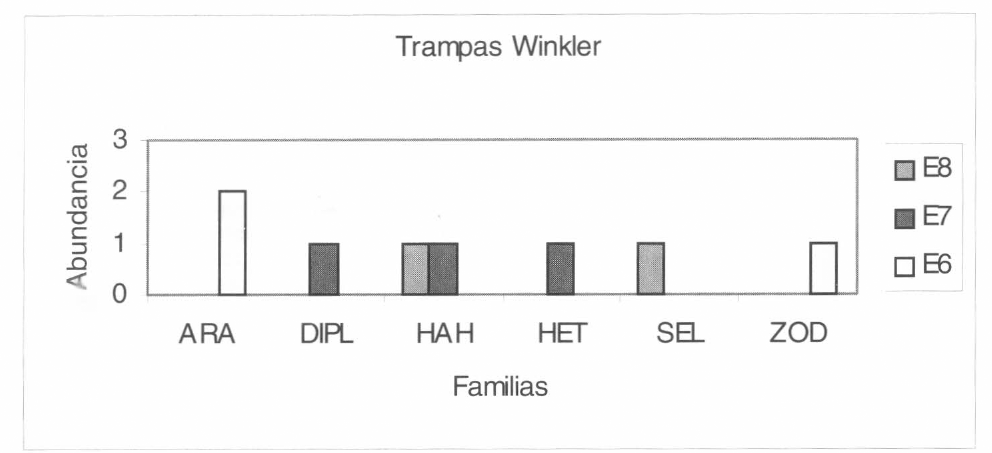
Grupo de familias registradas mediante la utilización de las trampas Winkler en los pastizales. ARA: Araneidae, DIPL: Dipluridae, HAH: Hahniidae, HET: Heteropodidae, SEL: Selenopidae y ZOD: Zodariidae.
La distribución de arañas de estrato ra- sante en los hábitats evaluados (interior de bosque y pastizal) se comparó con el coeficiente de disimilitud de Bray-Curtis que está basado en matrices de abundan- cias como también mediante el coeficiente de similitud de Dice, el cual está basado en matrices de presencia-ausencia.
Aplicando el coeficiente de disimilitud de Bray-Curtis, los resultados muestran altas diferencias entre las estaciones evaluadas y los hábitats a los que están asociadas (pastizal y bosque), observándose una di- similitud del 69% (Bray-Curtis = 0.69) para la comunidad de arañas de estrato rasante encontradas para los dos hábitats ante- riormente mencionados (Fig. 11). Con el uso del coeficiente de asociación de Dice, los resultados indican qué tan asociados están los hábitats evaluados, observándo- se una asociación del 39% (Dice = 0.39) para la comunidad de arañas de estrato rasante para los dos hábitats evaluados (Fig. 12), lo cual confirma el nivel de aso- ciación muy bajo entre estaciones que puede inferirse como nulo.
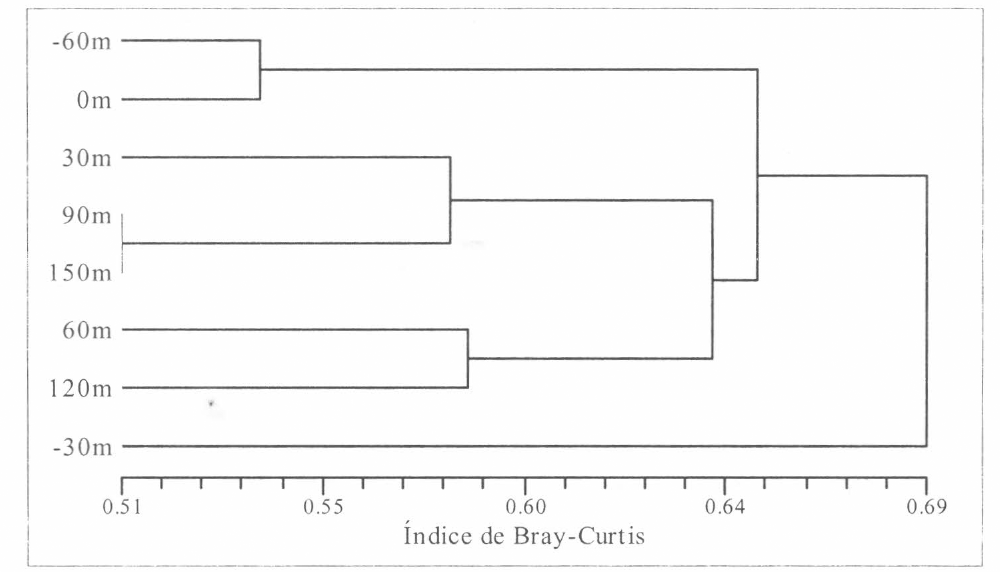
Coeficiente de Bray-Curtis para cada uno de los hábitats evaluados. -60 m: Estación 8,0 m: Estación 6, 30 m: Estación 5 (Interior de bosque), 90 m: Estación 3 (Interior de bosque), 150 m, 60 m, 120 m: Estaciones 1, 4 y 2 (Interior de Bosque) y -30 m: Estación 7 (pastizales).
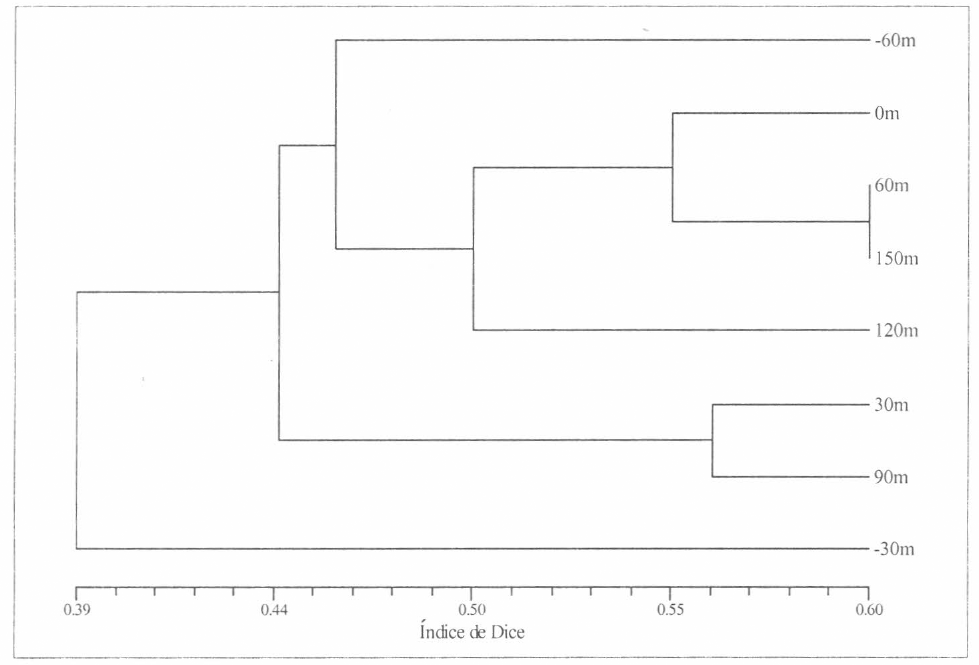
Coeficiente de Dice para cada uno de los hábitats evaluados.
Pese a la baja asociación observada, se des- taca la relación considerablemente mayor entre las estaciones de interior, lo que in- dica que la composición de las arañas de estrato rasante es más estable en el inte- rior del bosque. En cuanto al pastizal, se observa que no hay relación entre las es- taciones aunque la estación 6 (0 m) y la 8 (-60m) forman una unidad claramente de- finida. La estación 7 (-30m) se muestra como una unidad independiente dentro de las estaciones del pastizal, lo cual po- dría asociarse a que esta estación sea de distribución especial para un grupo espe- cífico de arañas de estrato rasante que pueda establecerse allí.
La riqueza esperada de arañas de estra- to rasante en el área de estudio se estimó mediante el programa "Estimates 5.01" (Colwell 1997) para el interior del bos- que y el pastizal. Según este programa y confrontando con los resultados presen- tados, el número de morfoespecies ob- servadas para todo el muestreo (Sobs: 86; incluyendo todo lo colectado en las estaciones) fue entre 84,21 y el 73,11% del esperado mediante los estimadores de Chaol y Michaelis-Menten, respecti- vamente. Estos resultados indican que la efectividad del muestreo general fue buena. En cuanto al número de especies con un sólo individuo "singletones" fue de 22 y el número de especies con 2 in- dividuos "doubletones" fue de 15 (Fig. 13). De acuerdo con este resultado, se consideró apropiado emplear índices de diversidad como descriptores de la co- munidad de arañas dado que la mayoría de las especies estaban representadas en la muestra.
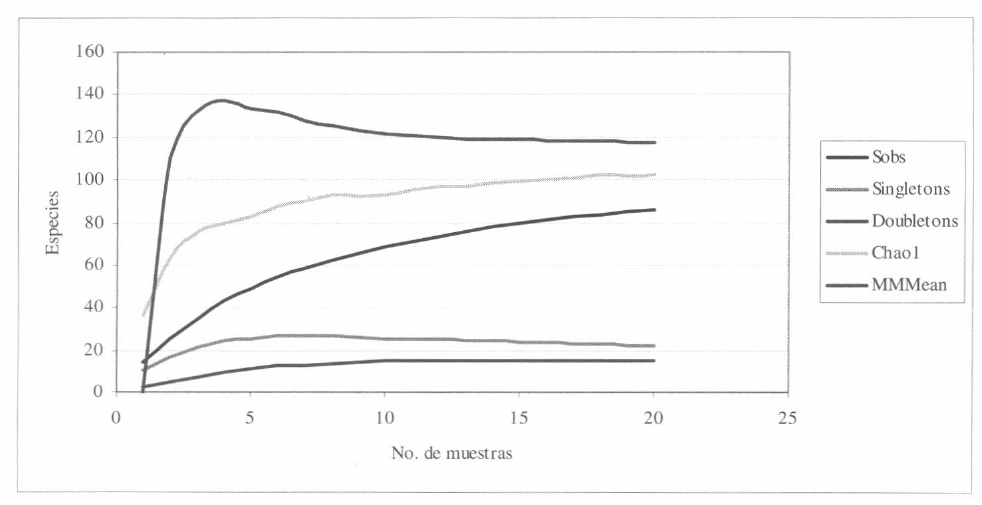
Curva de riqueza para las especies observadas (Sobs = 86) durante la fase de campo. Con un total de 20 muestras, se espera un total de 94 especies con Chaol y 120 especies con Michaelis-Menten (MMmean) en el piedemonte de la cordillera oriental en Toquiza, Cundinamarca.
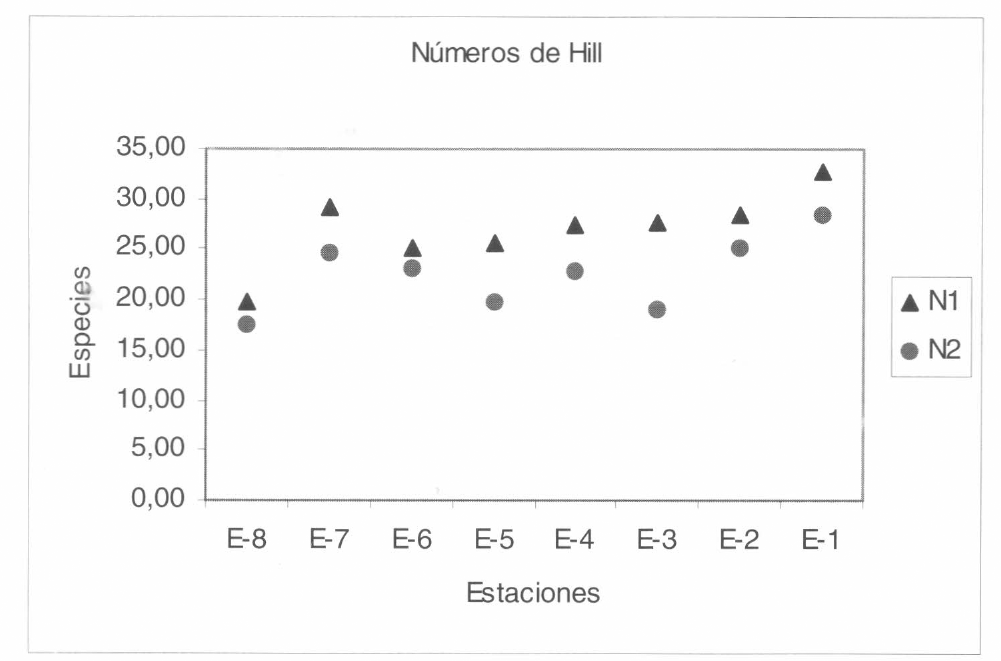
Variación de la diversidad de especies (expresada en los Números de Hill) para cada una de las distancias con respecto al borde de los hábitats evaluados.
Para cada distancia con respecto al borde, se estimó un valor de diversidad de arañas de estrato rasante relacionando los datos de riqueza y abundancia de las morfo- especies presentes en el bosque. A partir de estos valores, se estimó el índice de di- versidad expresado en los Números de Hill (Hill 1973) que muestran la relación entre el índice de Shannon y el índice de Simpson; así los Números de Hill reflejen aspectos diferentes de la diversidad de cada distancia (N, se refiere a las especies relativamente abundantes y N₂ se refiere a las especies dominantes). En la figura 14 se observa que la diversidad de arañas de estrato rasante expresada en N₁, presentó dos picos de diversidad alta claramente definidos en la estación 7 del pastizal (-30 m) y en la estación 1 del interior del bos- que (150 m). A partir de la estación 6 (0 m) y al ir ingresando al bosque se ve un aumento paulatino de la diversidad. La diversidad expresada en N₂ refleja tres pi- cos de dominancia claramente definidos en los 30 m externos y los 60 y los 120 m del interior del bosque.
Discusión
Se puede asociar la abundancia y riqueza de arañas de estrato rasante de las fami- lias Zodariidae y Liocranidae con la buena disponibilidad de lugares adecuados para la instalación de sus madrigueras en los sitios de estudio, tales como la hojarasca o bajo piedras, ambos abundantes de acuerdo con las observaciones de campo. Con respecto a Pholcidae, a pesar de que esta familia es tejedora, en campo ubica sus telas en cavernas que están al mismo nivel del estrato rasante, lo cual se corro- bora porque en el bosque evaluado se observan desniveles que se intercalan en- tre el piso de hojarasca y el estrato donde empiezan a crecer las raíces grandes y sóli- das de los árboles, lo cual concuerda con resultados similares obtenidos por Silva y Coddington en varios hábitats estudiados en Pakitsa, Perú (1996). También se puede atribuir a las diferencias entre la vegeta- ción rasante o herbácea con respecto a la vegetación arbórea y arbustiva presente en el bosque evaluado, la cual puede alber- gar un número potencial de microhábitats y de especies bioindicadoras, razón que se aplica en esta investigación dado que el efecto de borde presente en esta zona es bastante marcado. Este hecho también coincide con las apreciaciones de Fowler y Venticinque (1995), quienes detectan abundancias de arañas errantes y tejedo- ras asociadas al suelo y capturadas me- diante trampas pitfall en cuatro hábitats: bosque costero de tierras bajas, el suelo, el bosque atlántico y una plantación.
De Araneidae, por su hábito orbitelar, y Salticidae, por asociarse más con el follaje de herbáceas, no se esperaba una frecuen- cia alta en el estrato de estudio, ya que la distribución de Salticidae va desde el sue- lo hasta el dosel (Foelix 1996; Preston- Mafham y Preston-Mafham 1999) como también el no usar telas para la captura de sus presas y moverse entre el estrato ra- sante o herbáceo buscando a sus presas, lo cual concuerda con lo encontrado en Pakitsa, Perú por Silva y Coddington (1996). Con respecto a Araneidae hay que anotar que probablemente los machos en sus primeras incursiones en el hábitat bos- coso, cazan y acechan sobre el follaje y la vegetación rasante herbácea del bosque, lo cual posiblemente establecería una re- lación entre la complejidad estructural de la hojarasca y las abundancias de arañas tejedoras, ya que las hojas secas presentes en la hojarasca fomentan la presencia de telarañas y la abundancia de presas po- tenciales (principalmente insectos, sensu Uetz 1979) lo cual concuerda con la infor- mación presentada por Bultman (1981) у Bultman y Uetz (1982).
La familia Ctenidae, junto con las familias anteriormente mencionadas, está presen- te en la hojarasca confirmando lo señala- do por Levi y Levi (1993). Heteropodidae suele encontrarse en la hojarasca y en los cultivos de plátano o platanales (Levi y Levi 1993), cultivos que están presentes en el área de estudio, además de que otras zingiberales, como las Heliconia spp., fue- ron de las herbáceas más frecuentes en el área de investigación. Hay que anotar que el hábitat de hojarasca puede favorecer o afectar a estas familias de arañas por el suplemento de presas, reducción en las fluctuaciones microclimáticas, manteni- miento de la humedad como también la introducción de sustrato heterogéneo (hábitat modificado antrópicamente) y el aumento de espacio intersticial que existe entre la hojarasca, de acuerdo con las ob- servaciones hechas por Uetz (1975, 1979), Bultman y Uetz (1982) y Pelletier et al. (1998). También la heterogeneidad espa- cial en la hojarasca (Hatley y Macmahon 1980), puede influenciar la abundancia re- lativa de poblaciones de arañas por el incremento en las interacciones depre- dadoras que se presenten intra e interes- pecíficamente (Breymeyer 1966; Edgar 1969; Bultman et al. 1982). Entre otros factores, la estructura física de la hojarasca juega un papel muy importante en la dis- tribución y diversidad de estas familias de arañas, en el éxito reproductivo, actividad de forrajeo y presencia de depredadores (LaSalle y De La Cruz 1985; Brady et al. 1991).
Con base en los resultados obtenidos me- diante el índice de Bray-Curtis, se corrobo- ra la separación de las unidades del interior del bosque claramente diferencia- das de las estaciones del pastizal. Hay que resaltar que la estación 7 (-30m) se separa como unidad independiente de las esta- ciones 8 (-60 m) y 6 (0 m), con las que en teoría puede asociarse. Esta estación po- siblemente se comporta como una unidad de distribución especial, que marca la tran- sición abrupta entre microhábitats típicos de zonas abiertas, donde probablemente se dé una migración de las especies de arañas de estrato rasante entre este am- biente para llegar al interior del bosque.
Las 86 morfoespecies de arañas de estrato rasante registradas en el piedemonte cordillerano oriental constituyeron el 84,21% de lo esperado para Chaol y el 73,11% para Michaelis-Menten; es decir, se estiman entre 100 y 120 las morfoespecies para la zona de estudio, lo que corrobora la efectividad del muestreo realizado.
La diversidad de la comunidad de arañas de estrato rasante expresada en los Núme- ros de Hill (N, y N₂) presentó una tenden- cia a incrementarse hacia el interior del bosque para las dos variables, aunque fue más evidente para N,; esto se podría aso- ciar con una influencia creciente del efecto de borde al aproximarse hacia el borde, posiblemente determinado por el incre- mento drástico de las condiciones que imperan en estos ambientes (las variacio- nes en microclima, reducción en la dispo- nibilidad de presas, de refugios en las rocas, hojarasca o vegetación, e incremen- to de la exposición a depredadores). Hay que destacar que este comportamiento de la diversidad en la comunidad evaluada probablemente aumente con la compleji- dad estructural y la profundidad de la ho- jarasca, lo cual genera una gran cantidad de microhábitats estructurales posibles ha- ciendo que más especies de arañas coexis- tan en ellos (Uetz 1975, 1979), como también los cambios en diversidad que ocurren con los incrementos en las varia- ciones entre hábitats de áreas abiertas o pastizales y el bosque; además posiblemen- te las arañas asociadas al estrato rasante puedan responder a los cambios vertica- les de muy fina escala en la estructura de la hojarasca, con lo cual se espera que se genere al mismo tiempo un incremento en la riqueza y abundancia de las especies y en la profundidad de la hojarasca (Lowrie 1948; Huhta 1971; Anderson 1975, 1978).
Con la utilización de las trampas de caída, se encontraron cambios en la composición de especies, ya que se registraron familias como Zodariidae, Ctenidae, Salticidae, Araneidae, Liocranidae, Heteropodidae, Theraphosidae y Oonopidae. A diferencia de las trampas Winkler donde no se obser- varon cambios en cuanto a las abundan- cias, hay que anotar que éstas confirman y corroboran la efectividad de este método de captura según las pruebas hechas por Uetz y Unzicker (1976), en las cuales com- pararon la efectividad de muestreo de los cuadrantes y de las trampas de caída; ellos encontraron que los muestreos mediante las trampas de caída se correlacionan significativamente mientras que los de cua- drantes evidencian una relación no lineal con el número total de especies. Además, la técnica de los cuadrantes no muestra una fracción constante de especies, pero toma una fracción desproporcionadamente grande en diversas comunidades. Los da- tos que ellos presentan sugieren que las capturas que se hagan por el uso de tram- pas de caída, arrojan un estimado cercano al número total de especies en una comu- nidad, lo cual es de gran utilidad en estu- dios de diversidad de especies.
En cuanto a la utilización de las trampas Winkler, se observaron variaciones en la composición de las especies, información que corrobora su procedencia a partir de la hojarasca. Sus abundancias en el nivel de especies reflejaron un 26,92% total de lo colectado y observado, constituyéndo- se en un buen método de captura pero no el más efectivo, por lo cual se recomienda como un método complementario al de las trampas Pitfall.
Conclusiones
Las familias más abundantes en el área de estudio fueron: Zodariidae, Ctenidae, Araneidae, Heteropodidae, Salticidae, Pholcidae y Liocranidae.
El número mayor de morfoespecies se registró para la familia Araneidae.
Las morfoespecies más abundantes que se registraron en la región fueron: Zodarii- dae M1, Araneidae M5, Pamphobeteus sp. 2 (Theraphosidae), Pholcidae M3, Oonopi- dae M1, Ctenidae M2 y Heteropodidae M2.
La mayoría de las especies registradas se encuentran en el interior del bosque.
Las familias más abundantes en el inte- rior del bosque fueron: Zodariidae, Cteni- dae, Araneidae y Salticidae, mientras que en las áreas abiertas fueron: Ctenidae, Liocranidae y Zodariidae.
La mayor diversidad y distribución de especies de arañas de estrato rasante en el interior del bosque puede estar deter- minada por la mayor complejidad estruc- tural del hábitat, disponibilidad de presas y disponibilidad de sustratos en este hábitat.
La riqueza de especies de arañas de es- trato rasante, mediante los estimadores Chaol y Michaelis-Menten (MMmean), os- ciló entre 100 y 120 especies, valores co- rrespondientes al 84,21 y 73,11% de lo observado.
El método de captura más efectivo fue el de las trampas pitfall.
El pastizal y el interior del bosque eva- luados muestran claras diferencias en cuan- to a la distribución de individuos dentro de las estaciones.
Agradecimientos
A Eduardo Flórez Daza, Profesor del Insti- tuto de Ciencias Naturales de la Universidad Nacional de Colombia, por sus enseñanzas en la taxonomía de los diferentes grupos de arañas. A Diego Tobar, investigador del Instituto Alexander von Humboldt, por su asesoría en el análisis estadístico de los datos. Al Fondo "José Celestino Mutis" de la FEN-Colombia, por el apoyo financiero a este trabajo.