
Abstract
In order to understand the effect of the association of watermelon [Citrillus lanatus (Thumb) Matsum & Nakai] with corn (Zea mays L.), on Diabrotica balteata Lec. and Aphis gossypii Glover in watermelon, on Spodoptera frugiperda Smith and D. balteata in corn, and yields in the Ciénaga Grande de Lorica (Córdoba - Colombia), three associations were evaluated with watermelon (4.800 plants per hectare) with corn (10.000, 20.000 and 30.000 plants per hectare) and their respective monocultures, during the first semester of 2000. The experimental design used was a complete randomized block. Populations and the damage of the insects, yields, quality of the watermelon, profitability and the land equivalent ratio (ler) were quantified. Results showed statistically significance reduction in the population and damage of D. balteata and A. gossypii in associated watermelon with respect to the monoculture. In corn the incidence of S. frugiperda was less in 10.000 plants per hectare and statistical differences between treatments were not detected for D. balteata. The best yield and quality of watermelon were obtained in association with 10.000 corn plants per hectare: 21.941 kg/ha; and the least with 30.000 corn plants per hectare: 6.800 kg/ha; in the monoculture yield was 20.460 kg/ha. Corn yield varied between 1.280 and 2.813 kg/ha, proportional to the density of plants. Economically, the best treatment was watermelon with corn at the lowest density, with the highest index of 3,1 and a ler of 1,55; this was favored by a smaller population and incidence of the insects pests studied with a preference for monoculture over associated systems.
Introducción
Los cultivos múltiples o asociados son una alternativa de los sistemas de producción agrícola implementados principalmente por pequeños agricultores, dado que generan diferentes beneficios, como la diversificación de la producción en un mismo terreno y lapso de tiempo, mayores rendimientos agronómicos y mejor aprovechamiento de los recursos productivos. Otro beneficio de gran importancia es la contribución al manejo de problemas fitosanitarios como los insectos plaga, que disminuyen su incidencia por acción de una mayor diversidad biológica (Altieri 1976; Leihner 1983; Tobón 1997).
La contribución de los cuitivos múltiples al manejo de insectos plaga ha sido registrada por diferentes investigadores. Root (1973) definió las hipótesis de concentración de recursos y la de enemigos naturales, planteó que las plantas pueden adquirir resistencia a insectos por la asociación de cultivos, dada por las complejas interacciones de condiciones biológicas, físicas y climáticas del sistema. Altieri (1976) contrastó las bondades de la vegetación en condiciones naturales y la tendencia de mantener el equilibrio entre fitófagos y plantas hospederas. Algo similar fue expuesto por Perrin (1976) sobre la dificultad que tienen los insectos plaga para localizar las plantas hospederas en sistemas de cultivos diversificados, por la distorsión de estímulos visuales y olfativos de los insectos.
Altieri y Leibman (1986) expusieron la complejidad de los posibles efectos de los cultivos múltiples sobre insectos plaga, relacionaron una serie de aspectos, dentro de los cuales resaltan la interferencia entre el hospedero y la conducta de búsqueda de los insectos, como es el camuflaje, el contraste de fondo del cultivo, enmascaramiento o dilución de estímulos atrayentes y la repelencia química; otro efecto mencionado corresponde a la interferencia con el desarrollo y supervivencia de las poblaciones; así mismo, incluyeron las barreras mecánicas que dificultan la dispersión de los insectos, la ausencia de estímulos llamativos, las influencias microclimáticas y las influencias bióticas, como el alto grado de complejidad de enemigos naturales.
En la Ciénaga Grande de Lorica (Córdoba) durante la época seca (primer semestre del año) alrededor de 3.000 hectáreas son dedicadas cada año a cultivos transitorios de patilla, maíz, fríjol, pepino, habichuela, tomate y yuca, con tecnología tradicional en áreas de 0,5 a 2 ha por pequeño productor. Predomina la producción de patilla con cerca de 2.500 ha y rendimientos promedios de 11 t/ha; seguida por el maíz con unas 300 ha y rendimientos hasta de 1,6 t/ha. El principal limitante de estos cultivos son los insectos plaga, lo que ha conllevado a un alto uso de insecticidas y consigo a altos costos de producción (Secretaría de Agricultura 1999).
Para el cultivo de patilla, en el área mencionada, se han registrado como principales insectos plaga a Diabrotica balteata [Leconte (Coleoptera: Chrysomelidae)], Aphis gossypii Glover (Homoptera: Aphididae) y Bemisia tabaci [(Gennadius) Homoptera: Aleyrodidae] Secretaría de Agricultura (1999). D. balteata causa raspaduras y perforaciones irregulares en el follaje, también se alimenta de flores y yemas y su ciclo de huevo a adulto es de 30 a 40 días (Pulido y López 1973). A. gossypii ataca principalmente los brotes o yemas y por debajo de hojas jóvenes, algunas veces de forma generalizada, ocasiona arrugamiento, debilitamiento general y enanismo; excretan la “melaza” o “miel de rocío”, lo que genera el desarrollo de Capnodium sp. (hollín o fumagina); el estado adulto de este insecto es alcanzado alrededor de los 8 días (Vélez 1997).
La producción de maíz en el área de la ciénaga tiene como principal limitante a Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) (Secretaría de Agricultura 1999). Este insecto plaga actúa como gusano tierrero, trozador y cogollero; este último es el hábito más característico en gramíneas (García et al. 1999). El ciclo de huevo a adulto puede variar entre 20 y 35 días, con un estado de larva alrededor de 15 días, de acuerdo con las condiciones ambientales y el valor nutritivo del cultivo (Vélez 1997). Otro insecto plaga que afecta al maíz en el área mencionada, es D. balteata, se alimenta del follaje, espigas y estigmas o “cabellos” de las mazorcas; el estado de larva puede afectar el sistema radical (King y Saunders 1984).
Cuando se realiza el asocio patilla con maíz en el área de la Ciénaga Grande de Lorica, el pequeño agricultor, al comercializar la patilla, obtiene altos ingresos, mientras que el maíz lo utiliza para autoconsumo, siendo el interés principal del agricultor. En consecuencia, poco o nada se ha cuantificado el efecto de estos arreglos sobre la presencia de insectos plaga y sobre la eficiencia productiva de este sistema de cultivo. Por tal motivo, el objetivo del presente trabajo fue evaluar el efecto de la asociación patilla con maíz sobre la fluctuación poblacional y el daño de los principales insectos plaga, junto con el rendimiento de estos cultivos en la Ciénaga Grande de Lorica Córdoba. Se propuso cuantificar la fluctuación poblacional e incidencia de D. balteata en los cultivos de patilla y maíz, de A. gossypii en patilla y de S. frugiperda en maíz, así mismo medir la influencia del asocio sobre los rendimientos, la eficiencia en el uso del recurso suelo y analizar la viabilidad económica de este sistema productivo.
Materiales y Métodos
La presente investigación se realizó en el primer semestre del año 2000 (enero - abril) en un área de la Ciénaga Grande de Lorica - Córdoba, perteneciente al corregimiento de Sitio Viejo, 7 km al sur del municipio de Chimá (9° 15′ de latitud norte y 75° 3′ de longitud oeste); a una altura de 20 msnm, temperatura promedio anual de 27°C, precipitación anual de 1.250 mm, humedad relativa del 80%, brillo solar promedio de 7 horas día (Contraloría General 1999). El suelo con textura franco arcillosa, mostró un pH de 4,66, un alto contenido de materia orgánica (7,65%), una conductividad eléctrica de 0,15 mmhos/cm, niveles excesivos de S, Mg, Fe, y Zn, niveles altos de Ca y Mn, medios o moderados de Py Al y deficientes de K, Cu, y B, según el análisis realizado en el laboratorio de suelos de la Universidad de Córdoba Montería.
Se utilizó la variedad de patilla “Charleston Gray” y la variedad regional de maíz “Setentano”. La disposición espacial de los dos cultivos, de acuerdo con los tratamientos, se registran en la tabla 1. En la preparación del suelo se dejó una cobertura vegetal para protegerlo de las altas temperaturas y radiación (funcionando como mulch) y así mantenerlo con una humedad constante, dada la escasez de lluvias para la época (precipitación acumulada de enero a abril de 2000 igual a 40,9 mm, según estación 1307501 del IDEAM en Chimá - Córdoba); además esta cobertura funciona como cama para el fruto de la patilla, la cual contribuye con la calidad del mismo. El manejo agronómico fue uniforme para todas las unidades experimentales, que fue realizado de acuerdo con las condiciones del área y las exigencias de los cultivos.
Establecimiento en campo de los sistemas de patilla y maíz evaluados.
El diseño experímental fue de Bloques Completos al Azar (BCA), con cinco tratamientos (Tabla 1) Y cuatro repeticiones; cada bloque estuvo conformado por cinco parcelas de 10m de ancho por 12,5 m de largo para una área de 125 m? por parcela y 20 parcelas en total. El espaciamiento entre bloques fue de 2,5 m y entre parcelas de 1,5 m, resultando una área experimental de 3.360 m? y una área efectiva de 2.500 rn-. Las variables evaluadas se describen a continuación.
Población y daño causado por los insectos plaga
- Población e incidencia de D. balteata en patilla. El muestreo se realizó mediante observaciones en los seis sitios centrales de cada parcela: Muestreo inicial de todas las plantas, luego el tercio medio de las guías principales y finalmente se utilizó un cuadrado de 0,25 rn-. Se cuantificó la población (adultos) y daño fresco en cada sitio, donde la unidad de muestreo fue la hoja. Las evaluaciones se hicieron cada 4 días para un total de dieciocho.
- Población e incidencia de D. balteata en maíz. Las observaciones se realizaron en los diez sitios centrales. Se tomó como unidad de muestreo a toda la planta y se hicieron dieciocho evaluaciones, a intervalos de 4 días.
- Población y nivel de daño de A. gossypii en patilla. Se utilizó un cuadrado de 0,25 m’ para las observaciones en los seis surcos centrales; la unidad de muestreo fue la hoja. Se estimaron unos niveles de población de I a 4 (cero, de uno a diez, de once a veinte y veintiuno o más áfldos por hoja) y unos niveles de daño de 1 a 5 (hoja sin daño, hojas con bordes ligeramente entorchados, hojas ligeramente arrugadas y entorchadas, hojas arrugadas y entorchadas y por último hojas con presencia de miel de rocío). Para las dos variables se realizaron seis evaluaciones, con una frecuencia cada 4 días.
- Población e incidencia de S. [rugiperda en maíz. Se realizaron dieciséis evaluaciones, en las que se cuantificó el número de larvas y el porcentaje de plantas con daño fresco en el cogollo en 10 sitios de los surcos centrales de cada parcela. Las evaluaciones fueron realizadas cada 4 días.
Para las variables evaluadas de los insectos plaga se realizó un análisis estadístico mediante el programa de SAS/STAT, de acuerdo con el diseño de BCA, qué, por el sistema de evaluación de los insectos plaga, se constituyó en parcelas divididas en el tiempo, como lo sugieren Little e Hills (1976). Se realizó análisis de varianza por evaluación y para los promedios globales de cada una de las variables evaluadas y se aplicó la prueba de comparación múltiple de ‘fukey a la media de los tratamientos; realizando previamente las transformaciones necesarias para los datos de población con la formula (X+0,5)1/2 y para la incidencia o daño expresados en porcentaje arcosen (x/100)1/2 y de esta forma cumplir con los supuestos estadísticos de normalidad.
Componentes del rendimiento de los cultivos
Los componentes del rendimiento evaluados en patilla fueron: El número. y el peso en kg de frutos de primera, segunda y total por hectárea. El número de frutos de primera mayor de 6 kg, Y de segunda entre 4 y 6 kg. El área efectiva de cosecha por parcela fue de 37,5m2 correspondiente a los seis sitios centrales. El rendimiento (R) = [(Producción en cada parcela x 10.000m2)] / [(Área cosechada (37,5 m-j].
En maíz, se evaluaron el número de mazorcas por hectárea y el rendimiento en grano kg/ha. El área efectiva de cosecha fue de 24 m- correspondientes a los surcos centrales de acuerdo con la densidad de plantas. Para expresar los rendimiento en kg/ha se utilizó la fórmula propuesta por el CYMMIT (1985) donde R = [(kg por parcela x factor de corrección x 10) x (%Humedad x % desgrane)] / (área efectiva de cosecha).
Para determinar el efecto de los asocios sobre los rendimientos agronómicos se sometieron a análisis de varianza y a prueba de ‘fukey. Se aplicó además un análisis bivariado de varianza, donde se c nside ran dos variables simultáneas, esto implica análisis de varianza y covarianza en forma conjunta, siendo útil para no considerar individualmente la producción de un cultivo que ha interactuado con otro en la misma unidad experimental (Moreno 1990).
Parámetros económicos
Se evaluaron el costo total de producción en $/ha, el beneficio bruto en $/ha, el beneficio neto en s/ha y el índice de rentabilidad. Además, se estimó el uso equivalente de la tierra (uet). con el objetivo de conocer la eficiencia de cada sistema productivo en el uso del recurso suelo, donde uet = [(x1/Y1) + (x2/Y2)], x representa el rendimiento de cada especie cultivada en forma asociada; y representa el rendimiento de la misma especie en monocultivo.
En el análisis económico se tuvieron en cuenta los beneficios netos y la rentabilidad, comparados con los costos de producción de los sistemas evaluados, según lo propuesto por Leihner (1983), Y la evaluación de la eficiencia en el uso del recurso suelo con el uet, que representa el área relativa de tierra cultivada en monocultivo necesaria para obtener la misma producción que en la asociación, con base en lo propuesto por Soria el al. (1975), citado por Hart (1985).
Resultados y Discusión
Población y daño de D. balteata en patilla
Bajo las condiciones de investigación en el monocultivo, la población de adultos de D. balteata en patilla aumentó con el desarrollo del cultivo. La máxima población se registró entre los 36 y 44 días después de la emergencia (dde). donde se presentó la mayor población en patilla monocultivo (p(o)) y la menor en patilla por maíz 3 (pxm3) (Fig. 1). Las diferencias estadísticas se registraron en la evaluación a los 36 dde entre pro) con 4,7 adultos por sitio y pxm3 con 2,4 adultos por sitio (F = 4,37; df = 95 (modelo), 89 (error); P=0,04); a los 40 dde entre pro) y pxm3 (6,2 y 2,8 adultos por sitio respectivamente) (F = 8,53; df =95, 89; P=O,OI) y a los 44 dde entre pro), patilla por maíz 2 (pxm2) con respecto a pxm3 (5,5, 3,8, Y 1,8 adultos por sitio) (F = 15,2; df = 95, 89; P=0,01). Los promedios generales registraron valores de 1,3, 1,1, 0,9 у 0,7 adultos por sitio, para p(o), pxm1, pxm2 y pxm3, respectivamente, con diferencias estadísticas entre p(o) y pxm3 (F= 4,69; df= 1,727 * , 1,644 ** ; P=0,04).
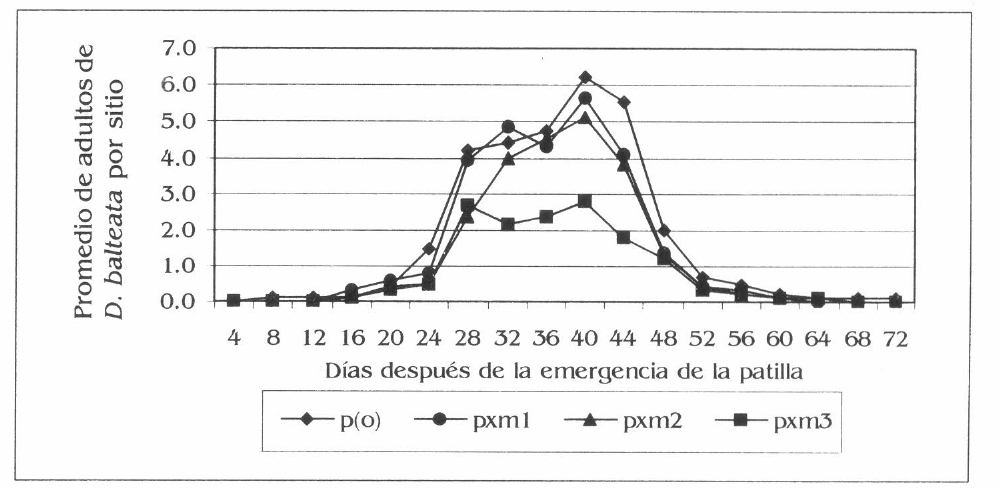
Fluctuación de la población de D. balteata en patilla, en la Ciénaga Grande de Lorica, Córdoba.
Grados de libertad (df) del modelo, incluye los df de los bloques (3), de los tratamientos (3) del error A (Bl.xTr.= 9). del número de lecturas (17), del tratamiento x lectura (51), del error B (Bl.x Lec). = 1,644.
Grados de libertad del error B.
El daño de D. balteata en patilla (porcentaje de hojas afectadas) presentó diferencias estadísticas a los 24 dde entre p(o) con respecto a pxm2 y pxm3, con promedios respectivos de 7,2, 4,6 y 4,3% de hojas afectadas (F= 4,14; df= 95, 89; P=0,01). A los 40 dde entre p(o) y pxm3 (19,0 у 13,8% de hojas afectadas, respectivamente) y entre pxm3 con patilla por maíz 1 (pxm1), con un porcentaje de hojas afectadas en este último de 17,55% (F= 4,59; df= 95, 89; P=0,01). P(o) y pxm3 también registraron diferencias a los 44 dde, el porcentaje de hojas afectadas fue de 16,5 y 9,2% respectivamente (F= 6,78; df= 95, 89; P=0,01) y a los 48 dde entre p(o) con 6,25% y pxm2 con 2,9% de hojas afectadas (F= 3,47; df= 95, 89; P=0,04).
Con base en los resultados descritos, se podría afirmar que la fluctuación del daño de D. balteata en patilla mantuvo una tendencia similar a la de la población, los promedios más altos en la mayoría de las evaluaciones se registraron en p(o) (Fig. 2). Los promedios globales reflejaron lo mostrado en las diversas evaluaciones, las diferencias se presentaron entre p(o) con respecto a pxm2 y pxm3, con valores de 6,9, 5,4 y 5,5% de hojas afectadas respectivamente, mientras que en pxm1 el promedio fue de 5,9% (F= 5,83; df= 1,727, 1,644; P= 0,04).
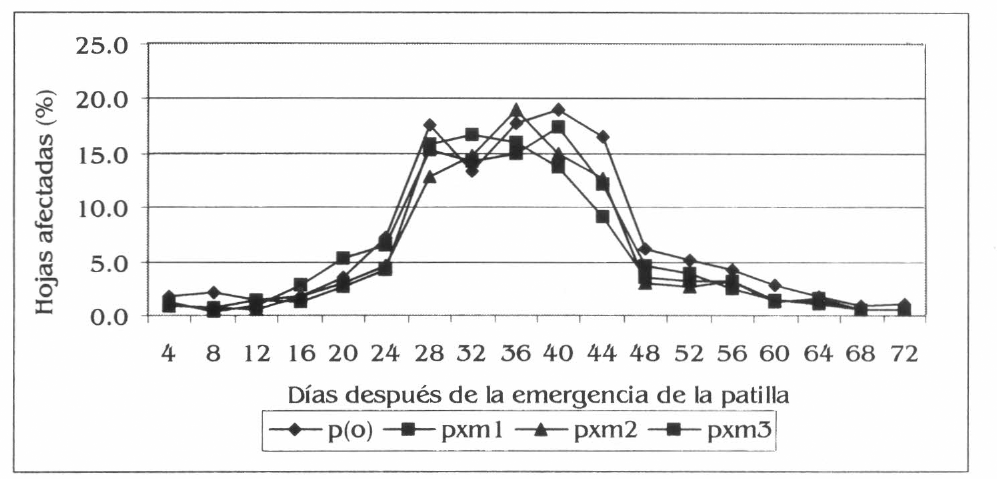
Daño promedio de D. balteata por hoja de patilla, en la Ciénaga Grande de Lorica, Córdoba.
Los resultados de la población e incidencia de D. balteata en patilla mostraron que los asocios con maíz tuvieron una influencia sobre este insecto plaga con respecto al monocultivo, lo que pudo ser debido a la mayor concentración de recursos, como lo propone Root (1973) y también por la formación de microclimas de preferencia o rechazo por parte del insecto (Altieri y Leibman 1986). La mayor presencia de D. balteata entre los 36 y 44 dde de la patilla se debió muy posiblemente a una nueva generación de la plaga y a una alta disponibilidad de alimento en el cultivo de patilla, con la mayor concentración de la población y del daño en el monocultivo. Por último, se presentó una correlación positiva significativa del 82% entre la población y el daño de D. balteata en patilla.
Población y daño de D. balteata en maíz
Las evaluaciones de la población y el daño de D. balteata en maíz solo presentaron diferencias estadísticas entre tratamientos a los 56 dde del cultivo. Para la población las diferencias se presentaron entre pxm2 con 1,8 adultos por planta y pxm1 con 0,7 adultos por planta (Fig. 3) (F= 3,8; df = 159, 153; P= 0,03). En los promedios generales de la población no se registraron diferencias estadísticas entre tratamientos, con valores de adultos de Diabrotica por planta para pxm2 de 0,2, m(o) con 0,17, pxm1 con 0,14 y pxm3 con 0,12 (F= 1,87; df= 2,879, 2,796; P= 0,12). Para el daño, las diferencias a los 56 dde fueron también entre pxm2 y pxm1 con valores respectivos de 33,3 y 19,2% de plantas afectadas (Fig. 4) (F= 2,67; df= 159, 153; P= 0,04). Los promedios generales del daño no registraron diferencias significativas, no obstante se presentaron valores de 4,9, 4,8, 4,7 y 3,7% de plantas afectadas en pxm2, m(0), pxm3 y pxm1, respectivamente (F= 1,74; df= 2,879, 2,796; P= 0,13).
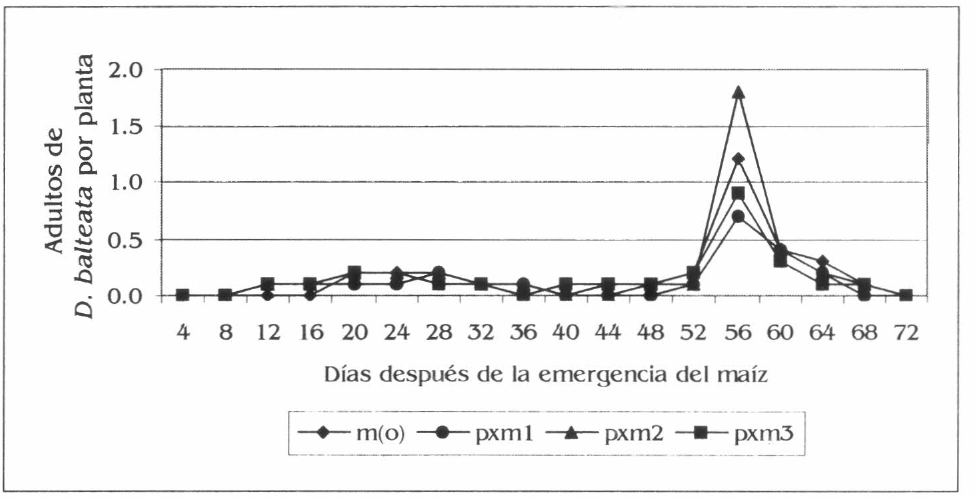
Fluctuación de la población de D. balteata en maíz, en la Ciénaga Grande de Lorica, Córdoba.
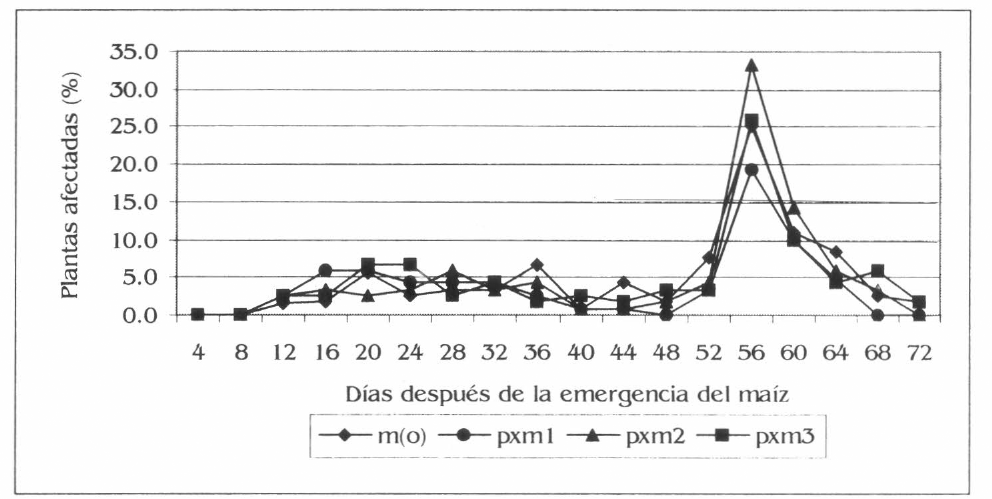
Porcentaje promedio de plantas de maíz afectadas por D. balteata, en la Ciénaga Grande de Lorica, Córdoba.
Al contrastar la población encontrada de D. balteata sobre plantas de maíz y la hallada sobre la patilla, se podría afirmar que este insecto plaga tiene una preferencia por el segundo cultivo, dado a la mayor presencia de adultos de este insecto que se registró sobre el cultivo de patilla, como se pudo observar en las figuras 1 y 3, descritas anteriormente. En maíz se presentó un aumento considerable de la población de D. balteata a los 56 dde, con respecto a las anteriores evaluaciones (Fig. 3), esto se debió a que en esta evaluación el maíz se encontraba en plena fase de floración, donde se observó que D. balteata tenia preferencias por las inflorescencias tanto masculinas como femeninas, esto explica la mayor presencia de adultos y de daño a esta edad del maíz. También se le podría sumar a esta situación, el inicio de la senescencia de la patilla, lo que ayudó a que el insecto encontrara una mayor disponibilidad de alimento fresco en el maíz.
Población y daño de A. gossypii en patilla
Los niveles de población de A. gossypii presentaron diferencias estadísticas en las primeras cuatro evaluaciones realizadas entre los 52 y 64 dde de la patilla. Las diferencias en las primeras tres evaluaciones fueron entre el monocultivo y los tres asocios; donde p(o) registró los niveles de población más altos así: evaluación a los 52 dde 2,0 (F= 11,16; df= 95, 89; P= 0,01), a los 56 dde 3,1 (F= 10,11; df= 95,89; P= 0,01) y a los 60 dde 3,8 (F= 8,65; df= 95, 89; P= 0,01); mientras que el menor valor en estas evaluaciones lo registró pxm3 con 1,0, 1,7 y 2,4, respectivamente. En la evaluación a los 64 dde la diferencia fue entre p(o) y pxm3, con valores correspondientes de 3,8 y 3,3 (Fig. 5) (F= 4,19; df= 95, 89; P= 0,04). En los promedios generales de los niveles de población de A. gossypii las diferencias se presentaron entre p(o) y los tres asocios con el mayor valor en p(o) (3,4) y el menor en pxm3 (2,5), mientras que pxm1 y pxm2 presentaron valores intermedios respectivos de 2,8 y 2,6 (F= 24.37; df= 540, 505; P= 0,01).
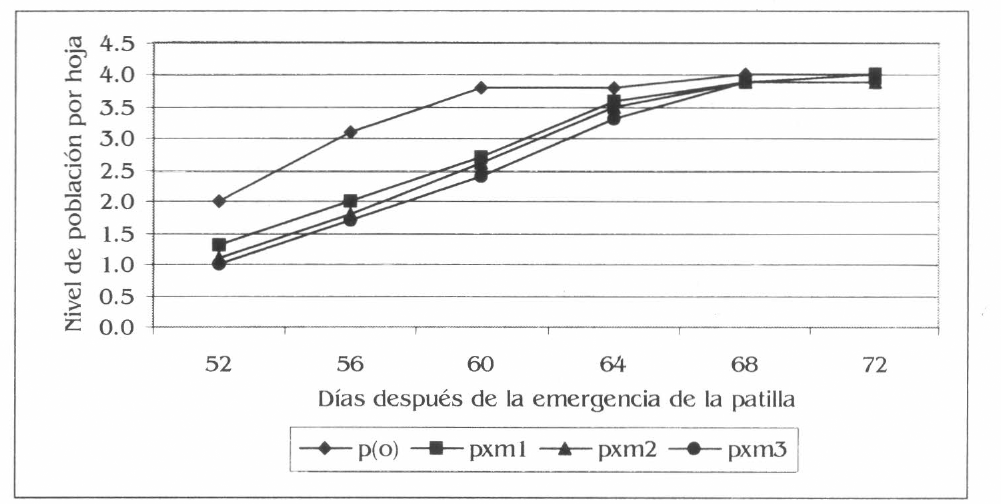
Fluctuación de niveles de población de A. gossypii por hoja de patilla, en la Ciénaga Grande de Lorica, Córdoba.
El nivel de daño de A. gossypii en patilla presentó diferencias estadísticas en cinco de las seis evaluaciones realizadas. En las primeras tres (52 - 60 dde) entre p(o) con los tres asocios evaluados (evaluación uno: F= 10,64; df= 95, 89; P= 0,01; evaluación dos: (F= 22,64; df= 95, 89; P= 0,01, у evaluación tres: F= 18,69; df= 95, 89; P= 0,01); en las dos siguientes (64 y 68 dde) las diferencias fueron especialmente entre p(o) y pxm3 (Fig. 6) (evaluación cuatro F= 7,75; df= 95, 89; P= 0,01), (evaluación cinco F= 2,74; df= 95, 89; P= 0,05). Los promedios generales del nivel de daño registraron diferencias entre p(o) y los tres asocios (F= 17,57; df = 540, 505; P= 0,01); p(o) con un valor de 3,4 fue el nivel de daño más alto, le siguió pxm1 con 5,2, pxm2 con 2,4 y el menor lo registró pxm3 con 2,2. La correlación entre el nivel de población y daño fue significativa y positiva, igual a 81%.
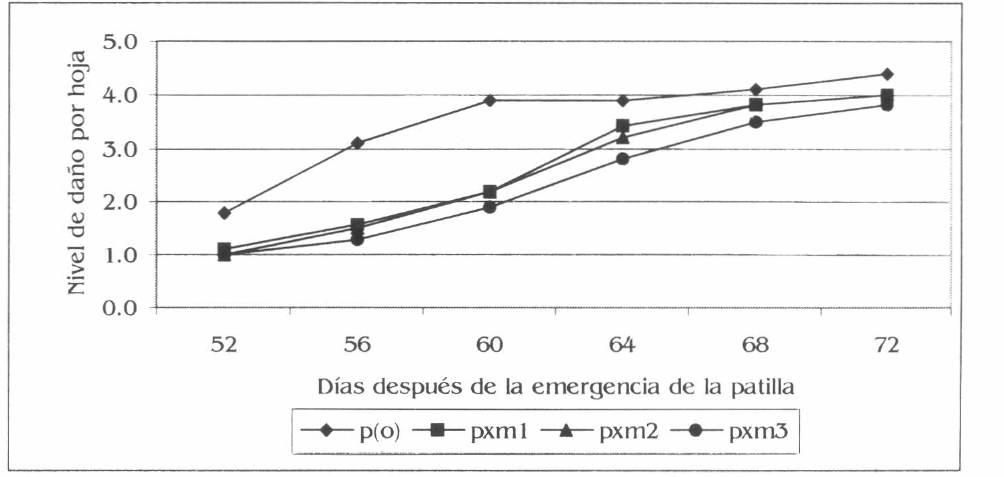
Fluctuación de niveles de daño de A. gossypii por hoja de patilla, en la Ciénaga Grande de Lorica, Córdoba.
La fluctuación de los niveles de población y de daño de A. gossypii en patilla, descritos anteriormente, muestran la influencia de los asocios sobre este insecto plaga, el cual registró los mayores valores para las dos variables en el monocultivo y disminuyeron en los asocios, principalmente en patilla con maíz a la mayor densidad (pxm3). Esto podría explicarse por la influencia que tiene el contraste de fondo de los cultivos sobre los áfidos, siendo atraídos por cultivos o plantas hospederas con un contraste de suelo desnudo, más que a un contraste de malezas, según lo expuesto por Altieri y Leibman (1986). En este estudio el contraste que pudo dificultar la llegada de los áfidos a la patilla fue el maíz, que también pudo interferir en el movimiento de los adultos alados limitando la colonización de nuevas áreas. Los resultados de la fluctuación de la población y del daño de A. gossypii en patilla pueden ser sustentados especialmente en las diferencias relacionadas con el hábito de crecimiento de los dos cultivos. El maíz con un crecimiento vertical y la patilla horizontal con respecto al suelo, conlleva a una estrecha asociación que da lugar a un contraste complejo entre estas dos especies de plantas. Esto estaría dificultando el movimiento de los áfidos (hembras con alas) que buscan colonizar otros sitios u otras plantas de patilla, las cuales, tendrían que superar la barrera física conformada por el maíz; además, los áfidos que vienen de otras áreas van a encontrar a la patilla camuflada por debajo del maíz, lo que distorsiona o dificulta la localización de su alimento preferido. Muchos otros aspectos biológicos y ecológicos de los áfidos estarían influenciados por los sistemas de cultivos implementados, entre ellos, los relacionados con los estímulos químicos y/o olfativos, que intervienen en la localización del alimento por parte del insecto, no obstante, sería necesario realizar estudios más específicos de la influencia de los cultivos asociados sobre los diferentes aspectos relacionados con el comportamiento de los áfidos o de algún insecto plaga en un área determinada.
Población y porcentaje de plantas afectadas por S. frugiperda en el cultivo de maíz
La población de S. frugiperda (larvas por planta) presentó diferenciasísticas entre tratamientos, desde los 20 hasta los 44 dde del maíz excepto a los 32 dde. A los 20 dde entre m(o) y pxml (F= 5,94; df 159, 153; P= 0,01); 24 dde entre m(o) y pxm2 (F = 3,27; df= 159; P<0,042); 28 dde entre m(o), pxm3 con respecto a pxm2 y pxm1 (F= 12,61; df= 159, 153; P= 0,01); 36 dde entre m(o) con pxm2 y pxm1 (F= 3,85; df= 159, 153; P= 0,05); 40 dde entre m(o) y pxm1 (F= 3,30; df= 159, 153; P= 0,05) y a los 44 dde entre m(o) con pxm2 y pxm1 (F= 7,51; df= 159, 153; P= 0,01).
El análisis de varianza para el porcentaje de plantas con daño fresco presentó diferencias estadísticas entre tratamientos a los 20 y 44 dde, así: 20 dde las diferencias fueron entre m(o) y pxm2 (F= 6,15; df= 159, 153; P= 0,01); 24 dde entre m(o), pxm3 con respecto a pxm1(F= 2,06; df = 159, 153; P= 0,05); 28 dde entre m(o) con pxm2 y pxml (F = 12,59; df = 159, 153; P= 0,01); 32 y 36 dde entre m(o) y pxm1 (F= 3,40; df = 159, 153; P= 0,05) (F= 3,47; df = 159, 153; P= 0,05) y a los 44 dde entre m(o) con pxm2 y pxm3 (F= 7,39; df 159, 153; P= 0,01).
La población y el daño de S. frugiperda aumentó con el desarrollo del maíz, con una disminución a los 24 dde (Figs. 7 y 8) debido a una precipitación de 12,5 mm un día antes de esta evaluación. Esto corrobora lo registrado por García et al. (1999), quienes afirman que las lluvias son un mecanismo de control de este insecto plaga. Los promedios generales mostraron diferencias estadísticas para la población y el daño de S. frugiperda en maíz; diferencias dadas entre m(o) y pxm3 con respecto a pxm1; para cada tratamiento m(o), pxm3, pxm2 y pxm1 la población registró promedios de 1,9, 1,6, 1,1 y 0,9 larvas por planta, mientras que el daño fue de 25,19, 20,98, 17,24 y 13,32% de plantas afectadas en forma respectiva (F= 3,65; df = 2,560, 2,485; P= 0,05) (F= 4,23; df 2,560, 2,485; P= 0,05). En las condiciones de la investigación, S. frugiperda registró una reducción de la población en un 55% y del daño en un 47% para el asocio pxm1 con respecto a m(o). Esto sugiere la preferencia de este insecto por el monocultivo de maíz, posiblemente al contrasté en forma, color o textura de los asocios, dado que la diversidad de los hábitats influye sobre S. frugiperda, como lo registra Altieri (1976) en policultivos de maíz asociado con fríjol y diversificados con malezas, la población de este insecto se redujo en un 23% y el daño en un 38% comparado con el monocultivo.
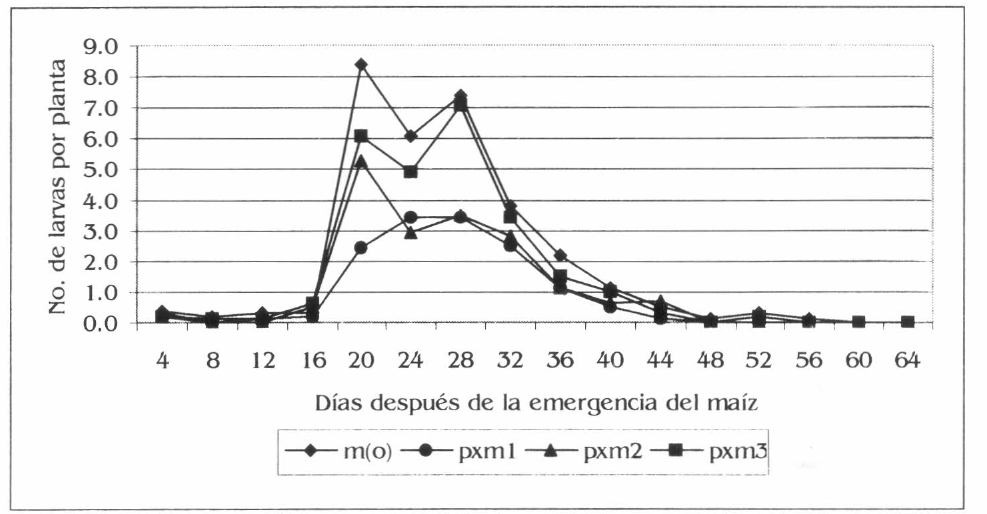
Fluctuación del número de larvas de S. frugiperda por planta de maíz, en la Ciénaga Grande de Lorica, Córdoba.
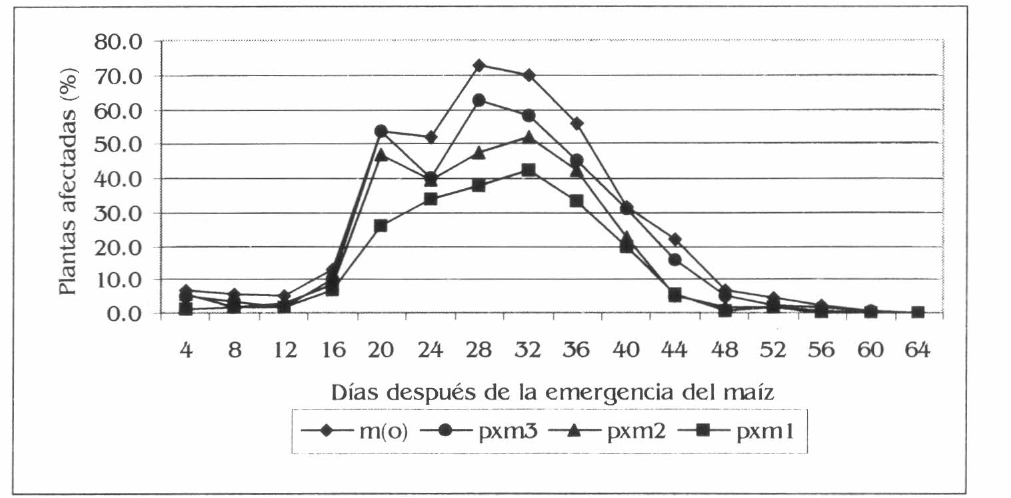
Fluctuación del porcentaje de plantas afectadas por S. frugiperda en maíz, en la Ciénaga Grande de Lorica, Córdoba.
Producción de la patilla
Para el número y peso de frutos de patilla de primera, las diferencias estadísticas se registraron entre pxm1 con pxm2 y pxm3; la producción fue de 1.667 frutos por hectárea= 13.614 kg/ha; 933 frutos por hectárea- = 6.960 kg/ha y 533 frutos por hectárea= 3.560 kg/ha, respectivamente (Figs. 9 y 10) (F= 14,04; df= 15, 9; P= 0,01) (F= 65,75; df=15, 9; P= 0,01).
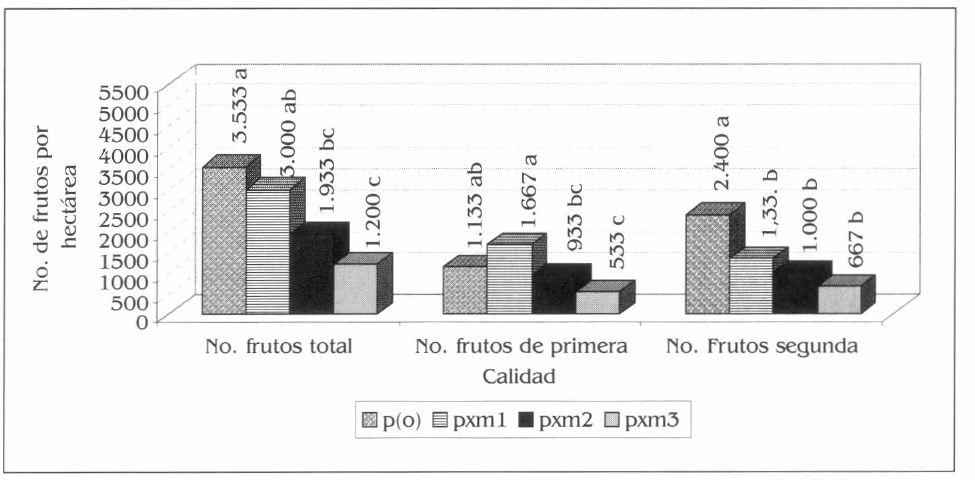
Producción promedio de patilla en número de frutos por hectárea para las diferentes calidades, en la Ciénaga Grande de Lorica, Córdoba. Promedios en columnas seguidos por la misma letra no son significativamente diferentes, prueba de Tukey 5%.
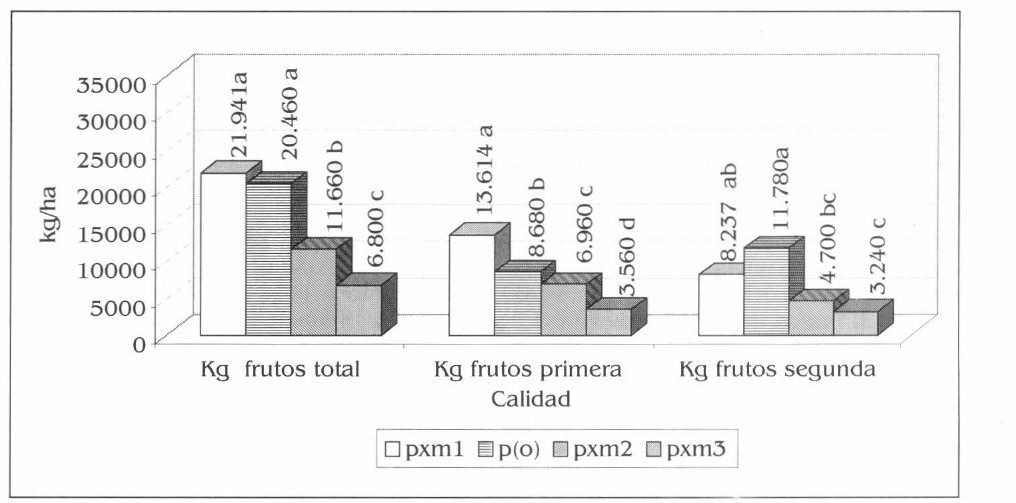
Producción promedia de patilla en kg/ha para las diferentes calidades, en la Ciénaga Grande de Lorica, Córdoba. Promedios en columnas seguidos por la misma letra no son significativamente diferentes, prueba de Tukey 5%.
El número y peso de frutos de patilla de segunda por hectárea presentaron diferencias estadísticas entre p(o) (2.400 frutos por hectárea = 11.780 kg/ha) con los tres asocios evaluados, la menor cantidad fue en pxm3 (667 frutos por hectárea = 3.240 kg/ha) (Figs. 9 y 10) (F = 21,15; df = 15, 9; P= 0,01) (F= 15,40; df= 15, 9; P= 0,01).
El número total de frutos de patilla por hectárea registró diferencias entre p(o) con respecto a pxm3 y pxm2, también entre pxm1 y pxm3 (F= 15,66; df= 15, 9; P= 0,01). El mayor número de frutos por hectárea se obtuvo en p(o) con 3.533 y el menor en pxm3 con 1.200 frutos por hectárea (Fig. 9). El peso en kg/ha mostró diferencias estadísticas entre pxm1 (21.941 kg/ha) con pxm2 y pxm3 y entre p(o) con estos dos últimos (F= 45,75; df= 15, 9; P= 0,01). El menor rendimiento fue el de pxm3 (6.800 kg/ha) (Fig. 10).
El rendimiento de la patilla en los asocios evaluados con la mayor densidad de maíz (pxm2 y pxm3) fueron menores con respecto al monocultivo, no obstante pxm1 registró los mayores rendimientos con el mayor número de frutos de mejor calidad, mientras que p(o) produjo el mayor número de frutos de segunda, contrastando esto con los rendimientos más bajos que fueron registrados en pxm3 (Figs. 9 y 10). Estos rendimientos estuvieron influenciados por cada sistema productivo, se podría afirmar que los sistemas de patilla con mayor densidad de maíz ocasionan una disminución en la captación de luz y de la capacidad fotosintética de la patilla, reflejada dicha situación en la producción baja de frutos y la calidad de los mismos, como se presentó en el sistema de pxm3. La mayor producción de frutos de segunda y la disminución en cierto grado de los rendimientos de p(o) con respecto a pxm 1 (Fig. 10), posiblemente se relaciona con la mayor exposición a factores limitantes como los insectos plaga estudiados, especialmente A. gossypii, que alcanzó rápidamente los máximos niveles de población y de daño en el monocultivo de patilla (Figs. 5 y 6), como se relacionó anteriormente.
Producción del maíz
Se registró un aumento en el rendimiento (número de mazorcas por hectárea y kg/ha en grano) a medida que se aumentó la densidad de plantas por hectárea, lo que muestra la poca o mínima influencia del cultivo de patilla sobre el rendimiento del maíz. La mayor producción se obtuvo en m(o) con 20.667 mazorcas por hectárea= 2.813 kg/ha en grano y el menor en pxm1 con 9.778 mazorcas por hectárea= 1.280 kg/ha en grano (Fig. 11). Se presentaron diferencias estadísticas entre tratamientos para el número de mazorcas por hectárea (F= 19,39; df= 15, 9; P= 0,01) y el peso del grano en kg/ha (F= 18.29; df= 15, 9; P= 0,01).
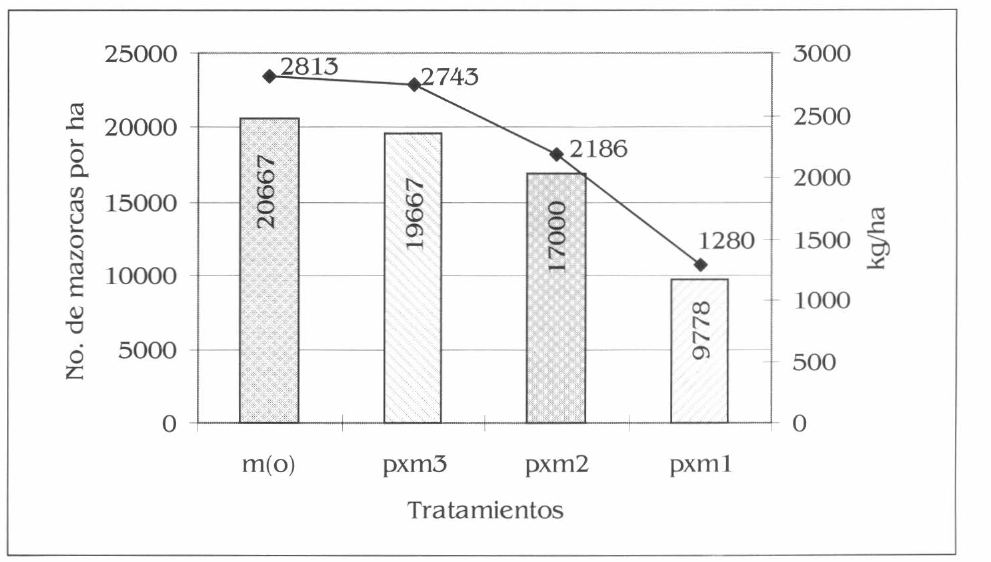
Producción promedio de maíz en No. de mazorca y kg por hectárea en la Ciénaga Grande de Lorica, Córdoba.
El análisis multivariado para los rendimientos en kg/ha de patilla y maíz en los tres asocios (pxm1, pxm2 y pxm3) mostraron diferencias altamente significativas entre cada uno de los tratamientos (F= 17,57; df= 11, 6; P<0,01) (siginificancia con base en el criterio de Hotelling) (Fig. 12). Es interesante anotar que el resultado de las pruebas de F en el análisis bivariado es consistente con el análisis por separado de los rendimientos de cada cultivo. En consecuencia, se podría afirmar que los asocios evaluados influyeron sobre la producción de los dos cultivos, especialmente sobre los rendimientos de la patilla que disminuyeron a medida que se aumentó la densidad de maíz en los sistemas asociados, mientras que en maíz la influencia pudo ser muy baja, dado que los rendimientos variaron de acuerdo con las densidades de siembra en cada tratamiento.
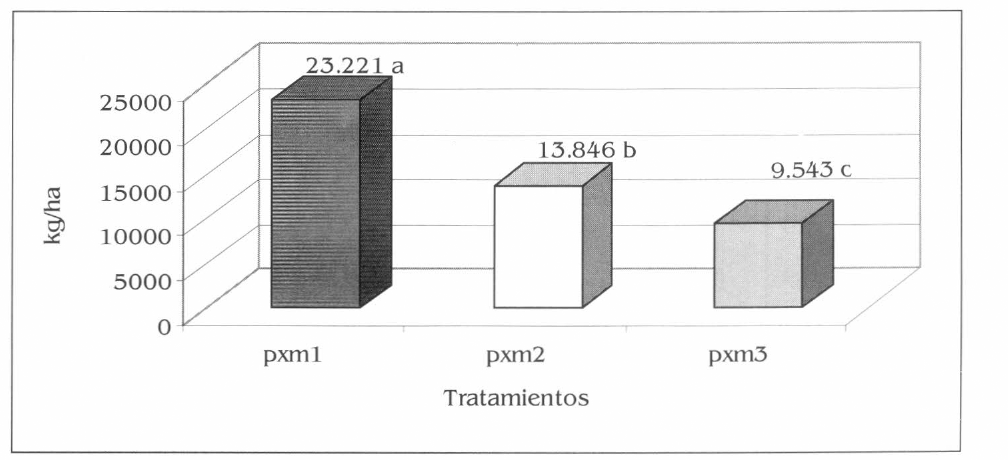
Rendimientos conjuntos totales de patilla y maíz en kg/ha, en la Ciénaga Grande de Lorica, Córdoba. Promedios en columnas seguidos por la misma letra no son significativamente diferentes, prueba de Hotelling al 5%.
Análisis económico
Se tuvieron en cuenta los costos de producción, los beneficios netos y la rentabilidad de cada sistema de patilla y maíz evaluado, como también para los monocultivos tradicionales en el área de influencia del experimento. El mejor tratamiento evaluado fue pxml, con unos beneficios netos por hectárea de $2.598.680 y un índice de rentabilidad de 3, 1, mientras que el menor fue m(o) con un beneficio neto de 656.380 $/ha y un índice de rentabilidad de 1,0. En patilla tradicional se registraron unos beneficios netos de 1.169.980 $/ha y un índice de rentabilidad de 1,3, superado por p(o), pxm1 y pxm2 evaluados (Fig. 13). En el índice de rentabilidad se presentaron diferencias altamente significativas entre pxm1 con los demás tratamientos evaluados junto a patilla y maíz tradicional en monocultivo (F= 17,57; df= 19, 12; P<0,01). Es de anotar que los costos de producción en patilla en monocultivo tradicional se incrementan con el uso de insecticidas (insumo no utilizado en el experimento) y en la posible falta de optimización de las diferentes prácticas agronómicas del cultivo, al igual que en el maíz. Esto corrobora la aplicabilidad comercial de los asocios pxm1 y pxm2, como también del sistema de p(o) evaluado.
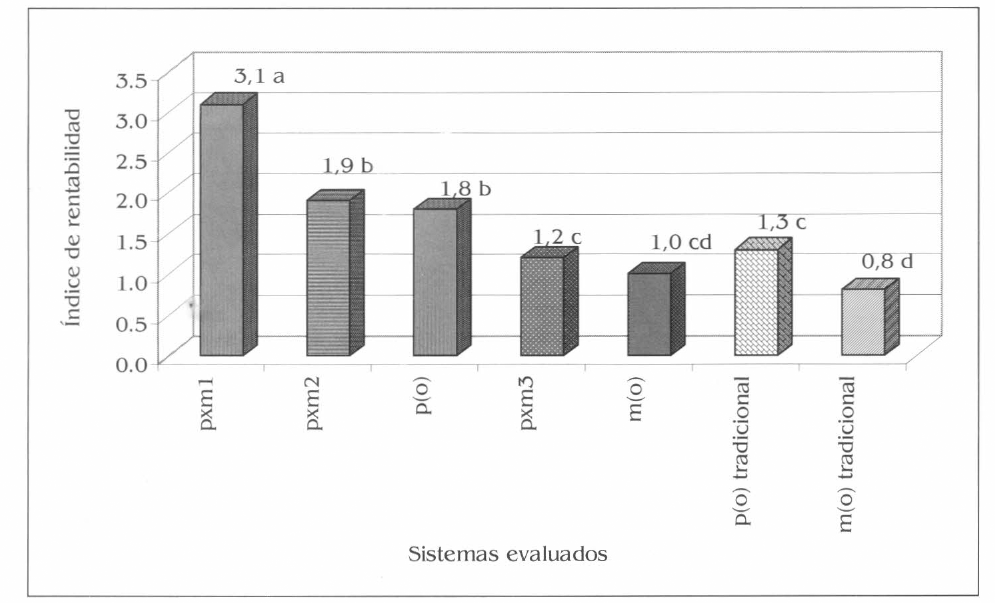
índice de rentabilidad de los sistemas de patilla y maíz evaluados y de los monocultivos tradicionales de patilla y maíz, en la Ciénaga Grande de Lorica, Córdoba. Promedios con la misma letra no son significativamente diferentes, prueba de Tukey al 5%.
Uso equivalente de la tierra (UET)
Los tres asocios evaluados fueron más eficientes en el uso del recurso suelo que los monocultivos (Fig. 14), se presentaron diferencias altamente significativas en pxm1 con respecto a pxm3, p(o) y m(o) (F= 30, df= 15, 9; P<0,01). Pxm1 fue un 55% más eficiente en el uso del recurso suelo en comparación con p(o) y m(o), por tanto se necesitarían 0,55 ha más de espacio o terreno al sembrar la patilla y el maíz en monocultivo. Este índice muestra la viabilidad de implementar la asociación de patilla con maíz en cultivos comerciales, en especial pxm1 donde se logra una óptima utilización del recurso suelo, recurso limitado en el área de influencia del estudio, en la que el sistema de asociación le genera al productor altos ingresos con la comercialización de la patilla y un bien básico para la alimentación como lo es el maíz.
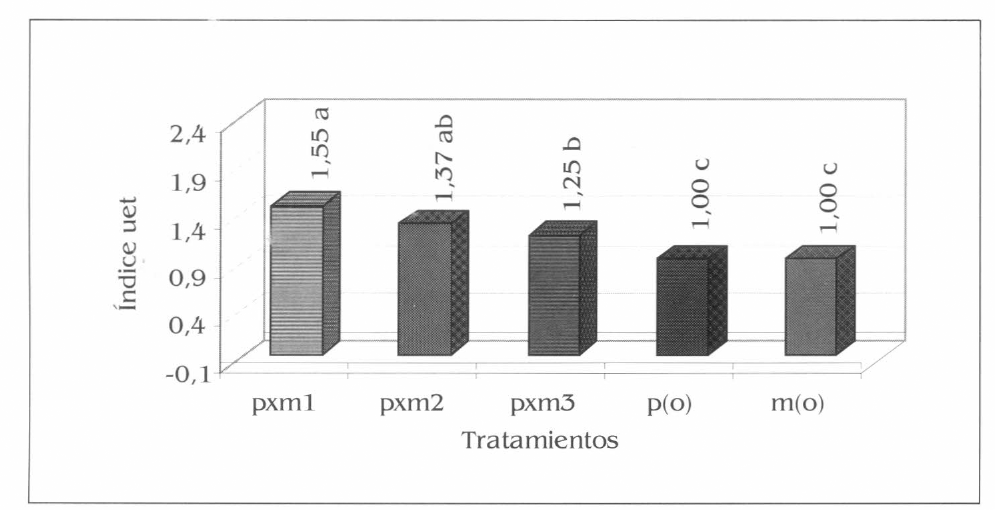
Uso equivalente de la tierra (uet) para los sistemas de patilla y maíz evaluados en la Ciénaga Grande de Lorica, Córdoba. Promedios en columnas seguidos por la misma letra no son significativamente diferentes, prueba de Tukey al 5%.
Conclusiones
Los diferentes asocios patilla con maíz estudiados afectaron la fluctuación poblacional y el daño de D. balteata y A. gossypii en patilla, así mismo a S. frugiparda en maíz y en menor medida a D. balteata en este último.
De los asocios estudiados el que contribuyó con la disminución de la población y del daño D. balteata y A. gossypii en patilla fue pxm3, no obstante pxm2 y pxm1 presentaron esa tendencia con respecto al monocultivo.
El asocio que mantuvo la población y el daño de S. frugiperda más bajo en maíz fue pxm1 con una reducción en un 55% de la población y en un 47% del daño fresco en este asocio con respecto al monocultivo; mientras que en pxm2 y pxm3 la población y el daño causado por este insecto presentaron valores similares al monocultivo.
Los rendimientos mayores de patilla se presentaron en pxm1 con 21.941 kg/ha, seguido por p(o) con 20.460 kg/ha y el menor rendimiento en pxm3 con 6.800 kg/ha, entre los cuales la mayor cantidad de frutos de primera se obtuvo en pxm1 y la mayor cantidad de frutos de segunda la registró p(o).
Los rendimiento en maíz aumentaron en forma proporcional a las densidades de siembra en cada tratamiento, con el mayor promedio en m(o) con 2.813 kg/ha y el menor en pxm1 con 1.280 kg/ha.
El sistema de cultivo pxm1 registró los mayores beneficios netos con un índice de rentabilidad de 3,1 en comparación con el monocultivo de patilla que mostró un valor de 1,8; pxm1 también fue el más eficiente en el uso del recurso suelo con un uet de 1,55. Económicamente éste fue el mejor tratamiento, superó ampliamente a p(o) y m(o) evaluados y a los cultivos tradicionales de la zona de estudio, además que contribuyó con el manejo de los insectos plaga evaluados.