
Abstract
Most studies on the family Noctuidae focus on very few species that cause damage to crops; however, there is little information on the biology and ecology of the remaining species of noctuid moths. This work examines the relationship of the larvae of Cucullia calendulae with their main food plant Calendula arvensis. The results showed that the larvae of this species feed on the fruiting bodies, reaching an overall incidence of ca. 30%. In addition, there was a differential distribution of larvae according to their size. Early stages were found in the reproductive structures while latter stages were found on the stems, which should be due to a defensive strategy. Furthermore, from seven characteristics of the patch taken into account, only the height of the plant explained a greater number of larvae per patch.
Introduction
Cucullia calendulae Treitschke, 1835 is included in the family Noctuidae, one of the most diverse families of the order Lepidoptera (Bretherton et al. 1979; Fibiger 1990). This species has the widest distribution of the genus in the Mediterranean basin, from southern Greece to the Iberian Peninsula, and it is characterized by having a single generation per year such that the first imago appears in October and the flight period lasts until April-May (Ronkay and Ronkay 1994). The larvae of C. calendulae feed mainly on the genus Calendula (Compositae), but also on species of the genera Ormenis, Archillea and Anthemis (Bergmann 1954; Merzheevskaya 1989; Yela 1992). The imago has a low dispersal ability and females lay from 50 to 150 eggs (Pérez-Guerrero 2001). There are numerous studies on the biology and ecology of species relevant to agroforestry (see revisions in Fitt 1989; Showers 1997) due crop damage and the associated economic costs. Generally, these species feed on determinate parts of the plants, including leaves, stems and reproductive structures (Cayrol 1972). In addition, there is a differential distribution of the larvae, based on size, on different parts of the plant (Eckel et al. 1992). Little information is available, however, about noctuid moths that are not considered pests (Rejmánek and Spitzer 1982; Yela 1992). In fact, this information is almost exclusively limited to zoogeographic, taxonomic and phenological data (Ronkay and Ronkay 1994). The aim of this work was to evaluate larval impact to the plant, describe the larval distribution on the plant at different development stages and describe the patch characteristics that determine a differential larvae density.
Methods
The study was conducted in Cordoba (Spain), in the south of the Iberian Peninsula (37°54’N, 4°42’W), in an area surrounded by olive orchards and wheat crops. A total of 19 randomly chosen patches of 1 m2 were scouted in an area where Calendula arvensis predominated, over the period of 2-22 March, 2001. For each patch, all larvae of C. calendulae were measured in mm and their location in the plant was recorded. We distinguished six categories according the location of larvae in the C. arvensis plant: 1) on the flowers, 2) close to the flower bud, 3) on the stem, 4) on the leaves, 5) on the fruits and 6) on neighbouring plants. Moreover, height in the center of each patch was registered. All plants included in the patch were collected and C. arvensis individuals were separated to be counted from the rest of neighbouring plants. Afterwards, fresh weight of C. arvensis and the remaining plants was measured with a dynamometer. In addition, the number of flowers, flower buds, and fruits of C. arvensis present in each patch were quantified. To assess the damage caused by C. calendulae, the number of damaged flowers, buds and fruit were also counted. Because C. calendulae does not eat leaves (personal observation), these were not evaluated.
The statistical treatment of data was carried out with Statistica v7.0 (StatSoft). Log-transformations were used to improve parametric conditions (Zar 1984). In order to analyze the distribution of stages over the plant, an analysis of variance (ANOVA) was used. Since some plant locations were occasional and not related to feeding behaviour, four larvae found on fruits of C. arvensis and two on neighbouring plants were excluded from the analysis. To analyze the influence of patch characteristics on larval density, a forward step multivariate regression analysis was carried out. Patch traits selected were number of flowers, number of buds, number of fruits, number of C. arvensis plants, dry weight of C. arvensis, dry weight of neighbouring plants and height of C. arvensis plants measured in the core of the patch. Plants were dried at 70°C for at least 48 h prior to recording their dry-weight.
Results and Discussion
An average (mean ± SE) density of 5.05 ± 0.46 larvae per square meter was found. The greatest number and percent of damage was found in buds (n = 807, which represents 38.95% of total), followed by flowers (n = 280, 16.14%) and finally the fruits (n = 255, 13.33%). Thus 30.01% of reproductive structures were damaged. The degree of damage is comparable to that produced by other noctuid species on crops, such as Helicoverpa armigera (Hübner, [1808]) in tomato, whose damage reaches 60% of the fruits (Montmany 1993), or Earias insulana (Boisduval, 1833) on cotton, whose damage reaches 40-50% of the bolls (Duran et al. 2000).
All larval instars were represented in the samples, with the intermediate size being most abundant. ANOVA showed differences in larval plant location according to size (F3,90 = 13.817; P < 0.001). Tukey post hoc tests revealed larger larvae to be more frequent on stems, and smaller larvae on flowers, buds and leafs (P < 0.01; Fig. 1). Because the largest larvae are easier to detect, their location on the stem may be a strategy for protection from predators as their coloration at this age is cryptic with the stems of C. arvensis (personal observation). In addition, the feeding larval activity may imply an increased risk of predation (Bernays 1997). With regard to patch selection, regression analysis showed that patches with the highest stems of C. arvensis had a greater number of larvae (R2 = 0.47; P = 0.002; y = -0.368 + 0.019*x); plant height was the only variable related to the density of larvae per patch. This may be due to greater detectability of plants by females during the laying period. The results obtained in this study are consistent with those obtained by other authors in relation to other noctuid pest species such as Helicoverpa zea (Boddie, 1850) (Eckel et al. 1992) and Spodoptera exigua (Hübner, [1808]) (Belda et al. 1994).
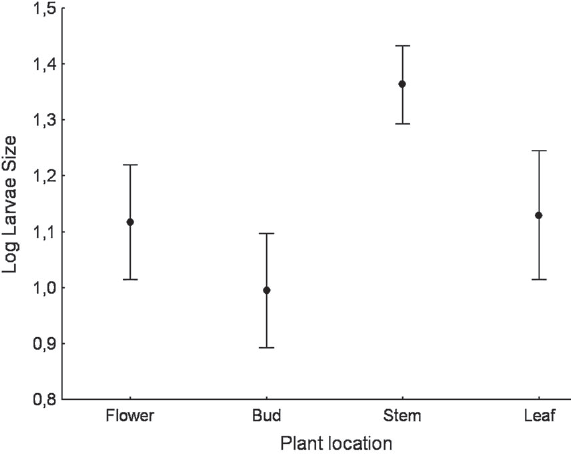
The size of C. calendulae larvae with respect to location on the plant (F(3, 90) = 13.817; P < 0.01).
Footnotes
Acknowledgments
We are grateful to J. L. Quero, Saskia Grootemaat and Enrique Doblas for help in translation and comments on the manuscript. Olmo Linares helped with the field samplings.