
Abstract
Spodoptera frugiperda (J.E. Smith) is a lepidopteran insect which is considered a pest in a great variety of crops in our country like cotton, rice, corn and shorghum. In the American continent it is widely distributed, that it is why some studies have been carried out developing some biological control strategies based on the natural enemies of the pest. In the present work it is evaluated the biopesticide activity of native isolates of Beauveria spp. and Metarhizium spp. against second instar larvae of S. frugiperda, in order to bring out the knowledge of the biopesticide potential of some entomogenous fungi native strains. A great variability in terms of the biopesticide activity was observed when it was evaluated 30 isolates from different host insect and geographic places. Of the 30 isolates, only 18 developed mycosis when they were passing through second instar larvae of the target insect. Those strains were tested in a bioassay by using 1×108 conidies.mL−1 against second instar larvae of S. frugiperda. In this phase, the variation in terms of mortality for the different isolates was between 0 and 93%. Seven of those isolates presented more than 50% of mortality, with a LT50 value between 1.41 and 6.31 days. In the second phase the LC50 was evaluated to the entomogenous fungi isolates with the lower LT50. The LC50 values were between 8.2×106 and 1.1×108 conidies.mL−1. The Beauveria spp. isolates IBUN-B-59, IBUN-B76 e IBUN-B-146, presented the higest percentage of mortality, the lower values of LT50 and LC50 and were very stable in terms of its biopesticide activity during the different evaluation phases, that is why it is suggested some field studies with those isolates.
Keywords
Introducción
Los hongos entomopatógenos, a diferencia de otros microorganismos empleados como estrategia de control biológico, no requieren ser ingeridos para infectar a sus hospederos, ya que su mecanismo de acción es la penetración directa a través de su integumento (Hall y Papierok 1982, St Leger 1993). Esta característica les confiere ventajas sobre otros entomopatógenos, como la capacidad de infectar estadíos que no se alimentan, o estadíos avanzados que no son tan susceptibles a agentes entomopatógenos de mecanismo de acción por ingestión, y la posibilidad de controlar especies de insectos picadores o chupadores que no pueden ser controlados por insecticidas cuyo principio activo debe ser ingerido por la larva. Además de lo anterior, a estos microorganismos se les atribuyen ventajas sobre los plaguicidas químicos como inocuidad ambiental, capacidad de producir epizootias y especificidad a hospederos.
Beauveria bassiana y Metarhizium anisopliae han sido los hongos entomopatógenos más utilizados a nivel mundial para el control de plagas agrícolas (Feng et al. 1994; Roberts y Yendol 1971), debido principalmente a su amplio espectro de acción y su fácil multiplicación. Diversas especies de estos géneros han sido observadas atacando larvas de S. frugiperda en diferentes regiones de Colombia (Rodríguez 1984; Pérez y Vergara 1998; Vargas et al. 1998). Sin embargo, muy pocos estudios han sido realizados en nuestro país evaluando cepas de estas especies para su control (Peréz y Vergara 1998), a pesar de que actualmente se encuentran varios productos comerciales disponibles en el mercado, a base del principio activo de Beauveria spp. y/o Metarhizium spp., para el control supuestamente de insectos del género Spodoptera o simplemente para el control de lepidópteros, como sugieren las etiquetas de presentación de dichos productos.
S. frugiperda o falso gusano ejército, es una de las plagas polífagas de mayor impacto económico en cultivos de gran importancia para el país como maíz, arroz, sorgo y algodón, por los daños que causa y los altos costos para su control (Vélez 1997; URPA 1998). La gran incidencia de este lepidóptero sobre los cultivos arriba mencionados, ha llevado a una sobre-utilización de insecticidas de origen químico, lo cual ha estimulado el desarrollo de resistencia a todos los grupos de insecticidas con que ha sido tratado (Zenner y Borrero 1996). Esta situación ha llevado al desarrollo de estrategias de Manejo Integrado de dicha plaga, los cuales involucran agentes de control biológico de diferente tipo, como la utilización de parasitoides, bacterias entomopatógenas como Bacillus thuringiensis y los mismos hongos entomopatógenos, los cuales presentan ventajas en algunas condiciones de cultivo gracias a su mecanismo de acción por contacto como ya se mencionó (García et al. 1999).
Aprovechando el gran potencial de los géneros de Beauveria y Metarhizium para el control de Spodoptera frugiperda, la importancia económica de este insecto plaga y la necesidad de generar alternativas para su manejo, el presente trabajo tuvo como objetivo identificar aislamientos nativos de hongos entomopatógenos de los géneros Beauveria y Metarhizium, que presentaran una alta patogenicidad sobre larvas de S. frugiperda de II ínstar en condiciones de laboratorio, mediante los parámetros de mortalidad acumulada, tiempo letal medio (TL50) y concentración letal media (CL50).
Materiales y Métodos
Cepa entomológica
La cepa entomológica de S. frugiperda provino de larvas colectadas en cultivos de algodón de los departamentos de Córdoba y Tolima. La cría del insecto se llevó a cabo en un cuarto climatizado, a una temperatura de 25 ± 2 °C, humedad relativa de 65 ± 5% y un fotoperíodo de 12 horas. Se siguieron procedimientos convencionales para el mantenimiento del pie de cría en insectario (López 1981).
Colección micológica
Se realizó una selección preliminar de 30 aislamientos (Tabla 1) del cepario del IBUN, teniendo en cuenta sus orígenes en cuanto a sitio geográfico y hospedero, procurando tomar grupos heterogéneos a fin de observar una posible asociación entre estas características y la virulencia de los aislamientos. Los aislamientos se mantuvieron en los medios selectivos para hongos, SDA® y/o Agar Trigo (AT: infusión de 30g de germen de trigo, 15g de agar-agar para 1L de agua destilada).
Procedencia y clasificación de los aislamientos nativos seleccionados
Aislamientos que no desarrollaron micosis en la prueba de reactivación, los cuales fueron descartados para las pruebas de patogenicidad.
Todas las zonas corresponden al departamento de Cundinamarca excepto la zona correspondiente al aislamiento 22.
Protocolo para bioensayo
Preparación de los insectos
Las larvas para bioensayo se tomaron a partir de una postura de por lo menos 100 huevos, para obtener material entomológico lo más homogéneo posible. Al momento de su emergencia, se llevaron a cámaras húmedas con dieta natural conformada por trozos de hoja de higuerilla (Ricinus communis) previamente lavada con agua destilada. Cuando las larvas llegaron a II ínstar, lo que ocurrió al 4° día después de la emergencia, se realizó el montaje del bioensayo.
Obtención de biomasa fúngica
Reactivación de aislamientos
Los aislamientos seleccionados (Tabla 1) se reactivaron sobre larvas de S. frugiperda. Para este propósito, las larvas se desinfectaron superficialmente por medio de inmersión en hipoclorito de sodio al 1% durante 2 minutos, posteriormente se realizaron tres enjuagues con agua destilada estéril (ADE). Las larvas así preparadas se depositaron en una caja de Petri con cada aislamiento previamente esporulado, se dejaron caminar por 5 minutos y posteriormente se llevaron en forma individual a cámara húmeda estéril sin alimento. Las cámaras húmedas se incubaron durante 10 días a 25 ± 2°C, 60 ± 5% de humedad relativa, y un fotoperíodo de 12 horas. Cultivos esporulados se obtuvieron a partir de los cadáveres de larvas con micosis, sembrando estos últimos en medio SDA. Tales medios fueron denominados Cultivos Madre. El cultivo de trabajo, el cual fue empleado para realizar las pruebas de actividad insecticida, consistió en el primer pase obtenido a partir del cultivo Madre, lo cual permitió evitar pérdidas de patogenicidad por efecto de repiques sucesivos (Daoust y Roberts 1982; Uribe 1995).
Obtención de la suspensión conidial madre
En primer lugar se verificó la viabilidad de ias conidias de los cultivos de trabajo mediante el procedimiento descrito por Goettel e Inglis (1996); posteriormente, éstas fueron depositadas en 5 mL de una solución de Tween 80 al 0.03%, la cual fue llevada a agitación en un agitador eléctrico (Multi wrist, Lab-Line, modelo 3589) por 15 minutos a velocidad de 7. La concentración de la suspensión conidial se determinó con un hemocitómetro (cámara de Neubauer mejorada), ajustándola posteriormente a la concentración deseada.
Infestación de las larvas
Tres grupos de 15 larvas de S. frugiperda de II ínstar se colocaron en recipientes plásticos estériles por separado. Un mL de la suspensión conidial fue vertido sobre cada grupo de larvas, las cuales permanecieron en contacto con la suspensión por dos minutos, bajo agitación suave; posteriormente, las larvas fueron vertidas sobre un tamiz, de donde pasaron de forma individual en condiciones de cámara húmeda a vasos plásticos de 2 onzas con tapa, en cuyo interior se colocó un trozo de papel servilleta empapado en agua destilada y un trozo de hoja de higuerilla. Las larvas así tratadas se llevaron a las mismas condiciones de incubación descritas arriba. El registro de mortalidad y micosis de los insectos se realizó diariamente durante 10 días.
Con el procedimiento de bioensayo descrito anteriormente se efectuaron las fases de evaluación de actividad insecticida, las cuales se describen a continuación:
Primera fase: Selección inicial
Para esta fase, los aislamientos de trabajo (Tabla 2), se evaluaron a una concentración conidial de 1 × 108 conidias.mL−1. Se realizaron registros diarios de mortalidad y micosis, y se estimó el TL50 para aquellos que superaron el 50% de mortalidad.
Adicionalmente, dos cepas aisladas de productos comerciales pertenecientes a Beauveria brongniartii (B. br), recomendada para el control de Spodoptera y otros insectos plaga y M. anisopliae (M.a) recomendada para controlar lepidópteros, se incluyeron como controles positivos de actividad entomopatogénica, junto con un aislamiento evaluado por Pérez y Vergara, (1998) sobre larvas de S. frugiperda de 10 días de edad.
Segunda fase: Concentración letal media (CL50)
Se hicieron cinco concentraciones seriadas establecidas como una sucesión geométrica de progresión 10, fueron hechas a partir de la suspensión conidial madre (desde 109 hasta 105 conidias.mL−1), para estimar la CL50. Los cultivos de trabajo en esta fase, se crecieron en el medio AT.
Diseño experimental
La variable a medir fue la mortalidad diaria de las larvas, confirmando su muerte, por micosis. Se utilizó un diseño completamente aleatorizado (DCA) con 3 repeticiones de 15 individuos cada una. Se utilizó la prueba de comparación múltiple Tukey con un α = 0.05, para observar diferencias estadísticas entre tratamientos (aislamientos). El Tiempo Letal 50 (TL50), se obtuvo por regresión polinómica estimada por el método de mínimos cuadrados, con transformación de Ln para los datos de porcentaje de mortalidad. La CL50 se obtuvo por análisis Probit con transformación de raíz cuadrada del porcentaje de mortalidad. Las larvas control para cada bioensayo, se trataron con solución acuosa de Tween 80 al 0.03%, sin esporas. Los análisis se implementaron bajo el software estadístico SAS.
Resultados
Reactivación
De los 30 aislamientos seleccionados inicialmente (Tabla 1), 12 fueron descartados para las pruebas de patogenicidad, por no desarrollar micosis; los 18 restantes (reactivados) se evaluaron en la fase de selección inicial.
I Fase: Selección inicial
Se observaron diferencias de actividad insecticida entre los 18 aislamientos evaluados en esta fase, encontrándose aislamientos sin actividad como el IBUN-MO de la especie M. anisopliae; con baja actividad (entre 4 y 30% de mortalidad), como los aislamientos IBUN-B-14, IBUN B-21, IBUN-B-23, IBUN-B-30, IBUN-B-37, IBUN-B-90, IBUN-B-93 e IBUN-M2; actividad intermedia (entre 31 y 60%) para los aislamientos IBUN-B-10, IBUN-B-19 e IBUN-B-29 y por último, aislamientos con actividad alta (mortalidades superiores al 60%), IBUN.B-16, IBUN.B-48, IBUN.B-59, IBUN.B-76, IBUN.B-79 e IBUN.B-146 (Tabla 2). Merece igualmente destacar que todos estos últimos aislamientos (excepto IBUN-B-16), poseen los porcentajes de micosis más altos, los cuales oscilan entre 55% y 81%.
Mortalidad en la primera fase de evaluación de actividad insecticida (1 × 108 conidias.mL−1), de los 18 aislamientos nativos reactivados y los aislamientos control positivo, con supuesta actividad contra S. frugiperda
Mortalidad acumulada al décimo día, promedio de tres repeticiones, las letras que acompañan el promedio de mortalidad pertenecen a la prueba Tukey a un a=0.05 (promedios con letras iguales no son estadísticamente diferentes).
Porcentaje de micosis presentada en las larvas muertas a lo largo del bioensayo.
Viabilidad de los aislamientos 24 horas antes de realizar el bioensayo, crecidos a 25°C.
El aislamiento IBUN 48 llegó al 100% a las 48 h de incubación.
La figura 1 muestra el promedio de tres repeticiones de la mortalidad acumulada hasta el décimo día, para cada uno de los aislamientos nativos evaluados en esta fase. Adicionalmente, se muestran dos cepas aisladas de productos comerciales pertenecientes a Beauveria brongniartii (B. br) y M. anisopliae (M.a) recomendadas para el control de Spodoptera y otros insectos plaga, las cuales fueron incluidas como controles positivos de actividad entomopatogénica, junto con un aislamiento evaluado por Pérez y Vergara (1998) sobre larvas de S. frugiperda de 10 días de edad, a quien los autores registraron con una m rtalidad del 85%. Sin embargo, estos aislamientos no mostraron acitividades superiores al 31% de mortalidad en las larvas de II ínstar. B. brongniartii, también evaluado como producto comercial, alcanzó solamente un 2.2% de mortalidad.
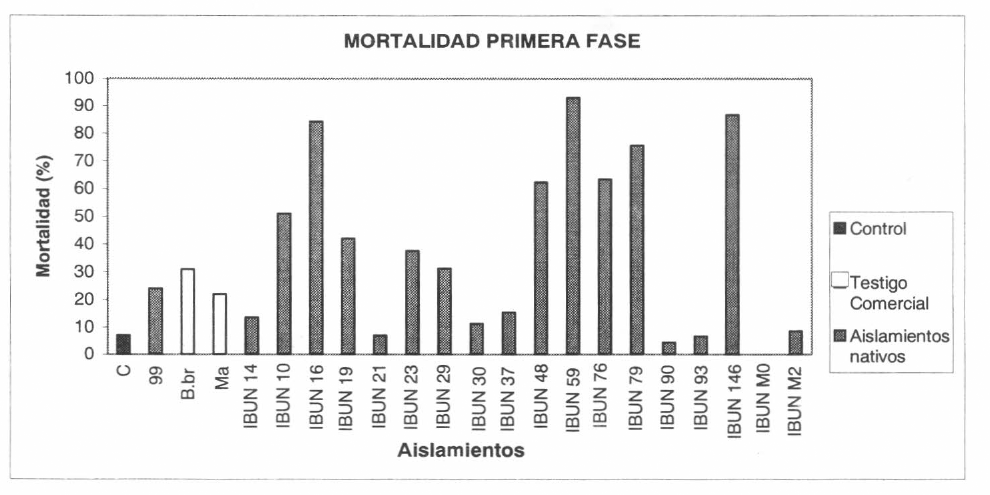
Mortalidad acumulada al décimo día de los aislamientos evaluados en la primera fase utilizando una concentración única de 1 × 108 co.mL−1.
A los aislamientos que superaron el 50% de mortalidad se les estimó el TL50 como se observa en la Tabla 3.
Valores obtenidos de TL50 para los aislamientos que superaron el 50% de mortalidad en la primera fase, con sus fespectivos límites de confianza a un α = 0.05
R2 = coeficiente de determinación. CV = coeficiente de variación. S2 = varianza.
En la figura 2 se muestra la distribución de la mortalidad en función del tiempo, observándose diferencias de actividad insecticida entre los aislamientos que alcanzaron el 50% de mortalidad; aquellos que obtienen los porcentajes de mortalidad más altos, son los que obtienen los menores TL50 y viceversa. Como se observa en la figura 2 y la tabla 3, los TL50 más bajos en orden ascendente los obtienen los aislamientos IBUN-B-59, IBUN-B-146, IBUN-B-79, IBUN-B-76, IBUN-B-16, IBUN-B-48 e IBUN-B-10.
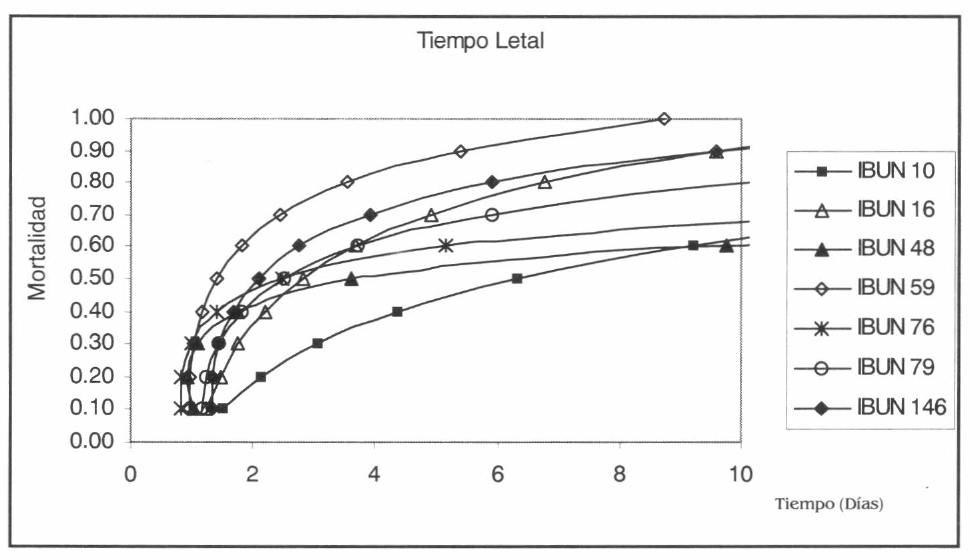
Distribución de la mortalidad en función del tiempo para los aislamientos que superaron el 50% de mortalidad en la primera fase, al ser evaluados a una concentración de 1 × 108 co.mL−1.
II Fase: Estimación de la CL50
Los aislamientos que superaron el 60% de mortalidad en la fase de selección inicial y que obtuvieron los TL50 más bajos, se evaluaron en esta fase, descartando el aislamiento IBUN 10 por presentar un TL50 muy alto y una mortalidad inferior al 60% (Tabla 3).
La figura 3 muestra los valores promedio de mortalidad acumulada (%), de tres repeticiones para cada concentración evaluada; también aparece la mortalidad promedio del tratamiento control para cada bioensayo de CL50 (C). Se observa el incremento de la mortalidad a medida que aumenta la concentración. Los aislamientos IBUN-B-59, IBUN-B-76 e IBUN-B-79 alcanzaron el 100% de la mortalidad con la máxima concentración empleada, mientras que el aislamiento IBUN-B-16 presentó el porcentaje de mortalidad más bajo (81%) en esta concentración.
En la tabla 4 se presentan los valores de CL50 para el décimo día de los aislamientos evaluados en esta fase, donde se observa que los límites de confianza para los aislamientos IBUN-B-59, IBUN-B-76 e IBUN-B-146 son muy estrechos, mientras que para el aislamiento IBUN-B-79 son un poco más amplios y aún mayores para los aislamientos IBUN-B-16 e IBUN-M-48.
CL50 al décimo día para aislamientos seleccionados, con sus respectivos límites de confianza
(α = 0.05)
Discusión
La colección de aislamientos nativos de hongos entomopatógenos proviene de aislamientos obtenidos en zonas de cultivos agrícolas donde no se ha aplicado previamente ninguna clase de producto biopesticida (cultivo de café), y de zonas de bosques silvestres (Uribe et al. 1998), donde existe poca o ninguna intervención del hombre en la introducción de agentes microbianos de este tipo. Las características mencionadas arriba permiten asegurar que ninguno de los aislamientos evaluados proviene de cepas de productos comerciales, algunos de los cuales son ampliamente utilizados en el país, para el manejo de insectos plaga de diversos cultivos. Por otra parte en tales zonas existe poca probabilidad de encontrar insectos plaga típicos de monocultivos, como son las larvas de S. frugiperda, que si bien son insectos altamente polífagos tienen sus preferencias por gramíneas (Vélez 1997). Estas características son de destacar ya que autores como Reithinger et al. (1997), Prenerová (1994), Lopéz (1994) y Prior (1992), al evaluar la actividad de hongos entomopatógenos contra Phlebotomidos, Cephalcia abietis, Bombix mori y langosta, respectivamente, han propuesto que el mayor potencial biocontrolador en hongos entomopatógenos debe provenir de aislamientos obtenidos del insecto blanco o especies estrechamente relacionadas; sin embargo, en este trabajo los aislados obtenidos a partir de cadáveres de lepidóptero (IBUN-B-19 e IBUN-B-23, Fig. 1) no presentaron actividad significativa, mientras que los aislados obtenidos a partir de coleópteros (IBUN-B-59 e IBUN-B-146) y homóptero (IBUN-B-76) presentaron alta actividad entomopatogénica sobre S. frugiperda (Fig.1, Tabla 3), sugiriendo que fuentes con potencial biocontrolador alto pueden encontrarse en órdenes de insectos diferentes al del insecto blanco, lo cual concuerda con los trabajos de Jiménez (1992), Varela y Morales (1996) y Wraight et al. (1998).
La primera fase de selección incluyó la evaluación de los 18 aislamientos que lograron ser reactivados en larvas de S. frugiperda, procedimiento que es llevado a cabo para lograr la preadaptación de los aislamientos hacia el estilo de vida patogénico (St Leger et al. 1991). A pesar de dicha reactivación se encontró un amplio rango de mortalidad entre los aislamientos, el cual estuvo entre 0 y 93% (Fig. 1, Tabla 2). Estos resultados sugieren que la actividad de los géneros Beauveria y Metarhizium contra S. frugiperda dependen de características intrínsecas de cada aislamiento, tales como origen geográfico, hospedero de origen, grado de esporulación, capacidad y tiempo de germinación, actividad de las enzimas asociadas al proceso de infección de los aislamientos, viabilidad de los conidios al momento de realizar el bioensayo entre otros, como lo sugieren los trabajos de Daoust y Roberts (1982), Jiménez (1992) y Bernal et al. (1994).
De acuerdo con los trabajos desarrollados por Kershaw et al. (1999), existen dos grandes estrategias de virulencia en los hongos entomopatógenos. La primera que consiste en aislamientos que producen niveles altos de toxinas, las cuales llevan rápidamente a la muerte de la larva, permitiendo que otros microorganismos oportunistas ocupen los espacios del hemocele al interior del insecto; tal estrategia termina en un crecimiento reducido del hongo entomopatógeno dentro del hospedero. Por otra parte, están los aislamientos que producen crecimiento del hongo en la hemolinfa, los cuales finalmente son reconocidos como aislamientos que ocasionan niveles altos de micosis. En la Tabla 2 se muestran niveles diferentes de micosis desarrollados por los aislamientos evaluados. Vale la pena destacar la micosis obtenida por los aislamientos que presentaron los niveles de mortalidad más altos, como el IBUN-B-16, que con un porcentaje de mortalidad del 84.3% mostró un porcentaje de micosis del 15.8%, mientras que el aislamiento IBUN-B-59 con un porcentaje de mortalidad del 93.3% obtuvo un porcentaje de micosis del 81%. Estos datos sugieren la presencia de diferentes estrategias de virulencia en los aislamientos analizados, siendo deseable la estrategia donde se desarrolle un mayor crecimiento micelial del hongo dentro del hospedero, en aras de aumentar las posibilidades de desarrollo de epizootias en campo.
Además de las diferencias mencionadas, se observaron variaciones en los TL50 entre los aislamientos que lograron valores superiores al 50% de mortalidad. El tiempo letal medio presentó un rango de 1.4 – 6.3 días (Tabla 3), tiempo que de acuerdo con el ciclo de vida de S. frugiperda, no permite el paso de las larvas a estados de pupa y adulto. Para los aislamientos nativos altamente patogénicos, los TL50 estuvieron entre 1.4 y 2.5 días (Tabla 3), tiempo que además impide el paso del insecto al siguiente ínstar de desarrollo larval, logrando que no escape del ataque fúngico por pérdida del inóculo en su exuvia. Aislamientos como el IBUN-B-59, que presentaron altos porcentajes de mortalidad, obtuvieron los TL50 más bajos (Fig. 2, Tabla 3). Por otra parte, los aislamientos IBUN-B-76 e IBUN-M-48 que tuvieron una mortalidad similar (63% y 62%, respectivamente) presentaron un día de diferencia en su TL50 (2.5 y 3.6 días, respectivamente), aspecto que posiblemente está asociado con que el aislamiento IBUN-M-48 haya tardado 24 horas más que el resto de los aislamientos, para obtener un porcentaje de germinación superior al 70% (100%, Tabla 2). Diferencias de este tipo en términos del TL50, son de gran relevancia como criterio de selección de cepas con buenos niveles de actividad contra un insecto plaga determinado, ya que en hongos entomopatógenos esto puede significar eficiencia y estabilidad de la acción entomopatogénica debido al mecanismo de acción de estos microorganismos. Dentro de este contexto cabe resaltar el hecho de que a pesar que el aislamiento IBUN-B-10 obtuvo un porcentaje de mortalidad superior al 50%, presentó un TL50 de 6.3 días que duplica y triplica el tiempo que los demás aislamientos requirieron para matar el mismo porcentaje de insectos, razón por la cual no fue seleccionado para la fase de CL50.
Con lo anterior se evidencia entonces, que no sólo pueden presentarse diferencias en el nivel de actividad (porcentaje de mortalidad, Fig. 1), sino en el tiempo requerido para alcanzarlo (Fig. 2, Tabla 3), señalando con esto, la importancia que tiene calcular este parámetro de virulencia para la selección de hongos entomopatógenos como agentes de control microbiológico. Lo anterior es especialmente cierto para insectos del orden Lepidoptera o de ciclo corto, por su condición holometábola, ya que cepas con un TL50 alto pueden permitir que el insecto a controlar pierda su inóculo al momento de cambio de exuvia.
La CL50 se estimó para los seis aislamientos seleccionados en la primera fase por presentar los TL50 más bajos y porcentajes de mortalidad superiores al 60%. Se observa un comportamiento típico dosis-respuesta, puesto que a medida que aumenta la concentración, aumenta el porcentaje de mortalidad (Fig. 3).
En lo referente a la CL50, los aislamientos IBUN-B-59, IBUN-B-76 e IBUN-B-146 presentaron los valores más bajos con un rango entre 9.35 × 106 - 1 × 108 conidias.mL−1 (Tabla 4). Rangos de CL50 similares se han encontrado para N. rileyi sobre larvas de Spodoptera litura de 11 ínstar y otros lepidópteros inoculados con B. bassiana (Vimala 1994; González et al. 1996). Los demás aislamientos evaluados en la fase de CL50 (IBUN-B-16, IBUN-M-48 e IBUN-B 7?) presentaron valores de CL50 en rangos similares, pero con límites de confianza muy amplios (9.8 × 105-8.9 × 1010 conidtas.ml−1, Tabla 4), sugiriendo inestabilidad en estos aislamientos en cuanto a su actividad bioplaguicida.
Conclusiones
Los resultados sugieren que aislamientos de hongos entomopatógenos provenientes de lepidópteros como hospederos, necesariamente no aseguran la obtención de cepas con una buena actividad biopesticida contra algunas especies similares o afines, como S frugiperda.
Los diferentes aislamientos de hongos entomopatógenos mostraron una variabilidad alta en términos de la actividad entomopatogénica, expresada en el amplio rango de mortalidad de las larvas, los diferentes niveles de micosis, TL50 y CL50, lo cual demuestra que la actividad de los géneros Beauveria y Metarhizium contra S. frugiperda, depende de racterísticas intrínsecas de cada aislamiento y no de los géneros en sí mismos.
El parámetro de TL50 es un criterio deter minante par la selección de aislamientos nativos promisorios con alta actividad entomopatogénica, especialmente para insectos del orden Lepidóptera de ciclo corto, por su condición holornetábola. ya que cepas con un TL50 alto pueden permitir que el insecto a controlar pierda su inóculo al momento de cambio de exuvia.
Los aislamientos IBUN-B-59, IBUN-B-76 e IBUN-B-146 presentaron la mayor actividad entomopatogénica, reflejada en porcentajes altos de mortalidad, TL50 y CL50 bajos, caracterizándose además por un comportamiento estable a lo largo de los diferentes bioensayos realizados en las mismas condiciones, con un rango estrecho entre los intervalos de confianza, por lo que evaluaciones a nivel de campo serían de interés para comprobar su actividad entomopatogénica en condiciones naturales, y de esta forma contemplar la posibilidad de introducir alguno de estos aislamientos dentro de un programa de Manejo Integrado para S. frugiperda.