
Abstract
The present work evaluated the aedeagus of males of the species D. martensis, D. starmeri, D. uniseta and D. aldrichi collected in the Colombian localities of Camarones near Riohacha, Santa Marta (North Coast) and Rosalía in the Tatacoa Desert (Huila Department). This character is one of those that has shown the most rapid evolution, favored by natural selection in the processes of new species formation, particularly in insects, and therefore it is considered an adequate indicator to determine the advancement of the aforementioned processes. Morphometric analysis with Fourier elliptical and MANOVA-CVA descriptors showed that among populations of D. martensis (Pxwilk=0.1816), as well as D. uniseta (Px wilk = 0.0507), there were no statistical differences in the form of the aedeagus, as opposed to that obtained for D. starmeri (Px wilk = 0.0004). This same test determined that in the three species of the martensis cluster, the genitalia of the males demonstrated significant differences (Px wilk = 4.23E-163). These results are evidence that each species in the martensis cluster maintains its taxonomic identity in the analysis. Likewise, when comparing each species of the cluster with D. aldrichi, another species of the repleta group collected in Tatacoa and belonging to the mulleri cluster, it was found that the data of D. martensis appeared to be superimposed over those of D. aldrichi, which would show the existence of a closer phylogenetic relation among these two species, than between D. martensis and others of the same cluster.
Introducción
El grupo repleta de amplia distribución en el Neotrópico es uno de los más grandes y más estudiados del género Drosophila (Etges et al. 2001; Mafla 2005). Los procesos evolutivos del grupo repleta y el conglomerado martensis en Colombia, han sido evaluados desde diferentes ángulos, que permiten visualizar la posible formación de razas geográficas y eventos de divergencia entre las especies, es así como se ha planteado que las poblaciones colombianas de la Costa Norte y el de-sierto de la Tatacoa de Drosophila martensis Wasserman & Wilson, 1957, D. starmeri Wasserman & Koepfer, 1973 y D. uniseta Wasserman & Koepfer, 1973, manifiestan divergencias morfológicas y reproductivas (Álvarez et al. 1989; Rojas 2001; Pérez 2002; Prieto 2008) y a nivel cromosómico y molecular la existencia de polimorfismos (De Polanco 1998; Prada et al. 2003; Ordóñez et al. 2004).
Frecuentemente la prueba de muchas hipótesis evolutivas requiere análisis de patrones morfológicos (Sota y Kubota 1998; Adams et al. 2002), pues es claro que el resultado final de todo el proceso evolutivo vivido por las especies puede reflejarse en la diversidad fenotípica, particularmente de caracteres de rápida evolución como el aedeago. De tal manera, la variación morfológica es uno de los criterios básicos para evaluar el avance de procesos como la formación de razas geográficas y la divergencia entre las especies, aún más si se presentan poblaciones aisladas en ecosistemas diferentes. Existe entonces una gran relación entre la variación de las características morfológicas y los procesos evolutivos particulares en las especies; por tal razón se consideró pertinente el análisis morfométrico del aedeago con el fin de evaluar estos aspectos en las poblaciones colombianas del conglomerado martensis. Para esto, la morfometría provee las técnicas que permiten cuantificar dicha variación y separar los componentes genético y ambiental del caracter examinado (Rohlf y Marcus 1993; Castañeda 2000; Jaramillo 2000; Jaramillo y Dujardin 2003). En este contexto en el presente trabajo se involucró el análisis de la forma del aedeago a través del análisis elíptico de Fourier de las especies D. martensis, D. uniseta y D. starmeri recolectadas en Riohacha y Santa Marta en la Costa Norte y el desierto de la Tatacoa (Huila) y D. aldrichi Patterson & Crown, 1940, recolectada solo en Tatacoa.
Materiales y Métodos
Se utilizaron machos recolectados en Camarones, Riohacha (12°28′N y 10°23′N, 71°06 y 63°W), y Santa Marta en la costa norte (11°15'N y 74°13′W) y en Rosalía en el desierto de la Tatacoa (3°N y 75°13′W) (Fig. 1), de las especies D. martensis, D. starmeri, D. uniseta y D. aldrichi (25 individuos por cada población y especie). Las recolecciones se realizaron en noviembre y diciembre del 2004 utilizando trampas con frutas fermentadas. Los adultos recolectados fueron sexados inmediatamente y se colocaron en grupos de máximo 15 individuos en frascos con medio de cultivo, modificado del de Sans de la Rosa et al. (1987). El establecimiento de un cepario se hizo necesario debido a la dificultad que presenta la determinación de los organismos en campo, en consecuencia de su carácter de especies sinmórficas, así mismo, la plasticidad fenotípica propia de los organismos, tendrá un menor efecto sobre las diferencias morfológicas, como resultado de un ambiente de laboratorio uniforme. Como base para la determinación de las especies, se tomaron las descripciones morfológicas de Wasserman y Wilson (1957), Strickberger (1962), Vilela (1983) y De Polanco (1998).
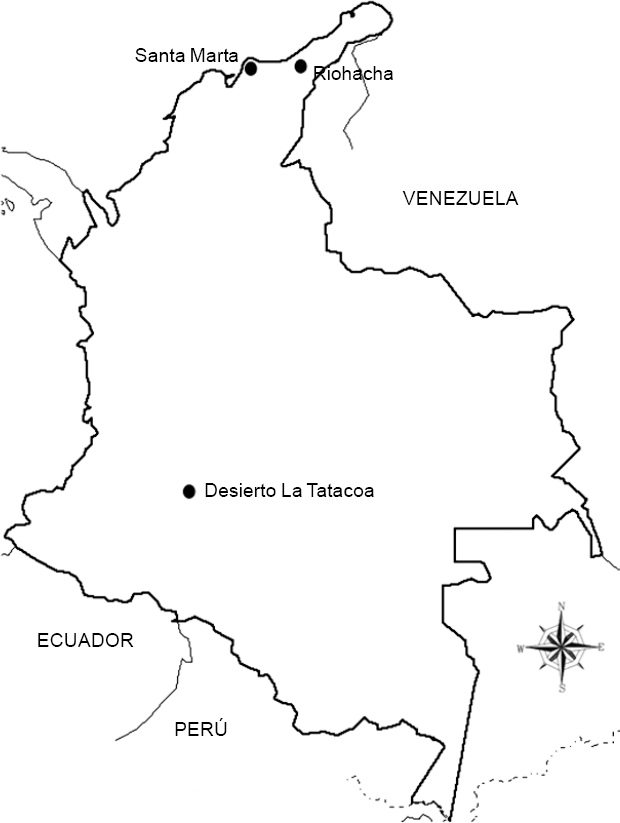
Mapa de Colombia y la ubicación de las tres zonas de muestreo.
Se dejaron machos adultos inmersos en KOH IN durante seis días y posteriormente, se guardaron en frascos con alcohol (Brower 1996a, 1996b). El abdomen se diseccionó directamente en una gota de KOH y las estructuras quitinosas (aedeagus e hypopygium) se limpiaron en un estereoscopio e inmediatamente se llevaron al microscopio y se fotografiaron con cámara digital con objetivo de 40X.
El método geométrico utilizado en este trabajo, Series de Fourier elípticas, ha sido probado en otras investigaciones que involucran la genitalia de los machos, para tratar de evidenciar tipos de selección sexual postcopulatoria y herencia aditiva en caracteres cuantitativos (Liu et al. 1996; Laurie et al. 1997) se basa en la toma de coordenadas del contorno de la estructura a analizar “Outlines” y en el ajuste de los puntos con la función matemática de la ecuación:
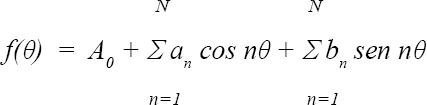
Esta ecuación reproduce gráficamente la forma de la imagen de la estructura originalmente digitalizada, luego de varias repeticiones. Las curvas se comparan, usando los coeficientes que representan la forma en una matriz comprendida por las variables producto de las repeticiones en las armónicas, en un análisis multivariado (Adams et al. 2002; Galindo 2004). El método utiliza la forma del aedeago conservando su relación geométrica a lo largo del análisis como una configuración de datos en un espacio invariante con respecto a los efectos de la rotación, la escala y el desplazamiento de la estructura, tomando en cuenta la información proveniente de las relaciones espaciales entre las variables, en este caso las coordenadas que definen la forma (Lestrel 1997; Rohlf 1999; Adams et al. 2002).
Para este análisis se utilizaron los programas ACDSee 5.0 (copyright 2003 ACD, Systems Ltd.) para dibujar sobre la fotografía, TPSdig para convertir los puntos del dibujo a coordenadas TPS - EFA para hacer el paso de los datos a EFAWIN, EFAWIN para las series de Fourier elípticas (Laurie et al. 1997), Efa-estadística para convertir los datos de EFAWIN a matrices de extensión .TXT que pueden ser trabajadas desde Excel, Excel para construir las matrices, PAST 1.22 (copyright Hammer et al. 2001), para los análisis multivariados y Notepad para transcribir datos. Los programas de la serie TPS y EFA WIN se encuentran disponibles en <http://life.bio.sunysb.edu/morph/>.
Resultados y Discusión
Los individuos analizados presentaron una amplia variación inter-especifica en la forma del aedeago, mostrando una forma particular en cada especie (Fig. 2). Las tres especies del conglomerado martensis mantienen su identidad taxonómica, manifestando diferencias estadísticas entre si en el carácter utilizado. Esto se refleja en el análisis canónico discriminante (CVA), entre las cuatro especies presentes en la Tatacoa: D. martensis, D. uniseta, D. aldrichi y D. starmeri (Fig. 3), el cual mostró diferencias entre las especies (Pλ wilk = 7,516E-83, PA wilk = 1,357E-41), al igual que el CVA entre Drosophila aldrichi y las tres especies provenientes de Camarones (Pλ wilk = 3,286E-63, Pλ wilk = 2,189E-30) (Fig. 4).
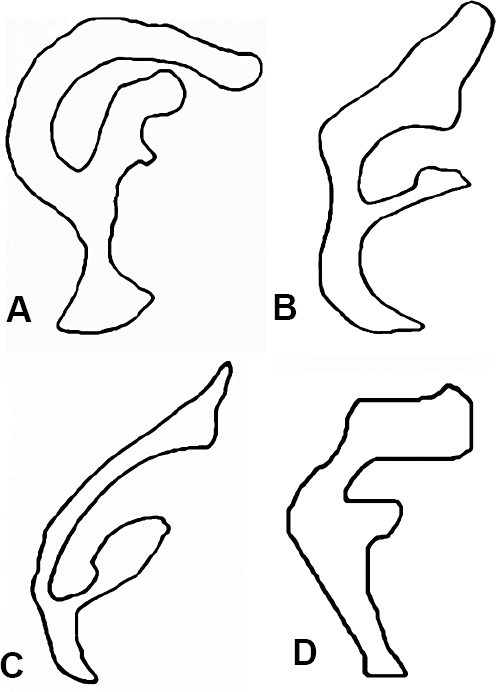
Contorno digitalizado del aedeago a partir de imágenes digitales de 400X:
Ordenamiento de los ejes canónicos luego del análisis discriminante, para la forma del aedeago a través del método “outlines” en las cuatro especies de Tatacoa.
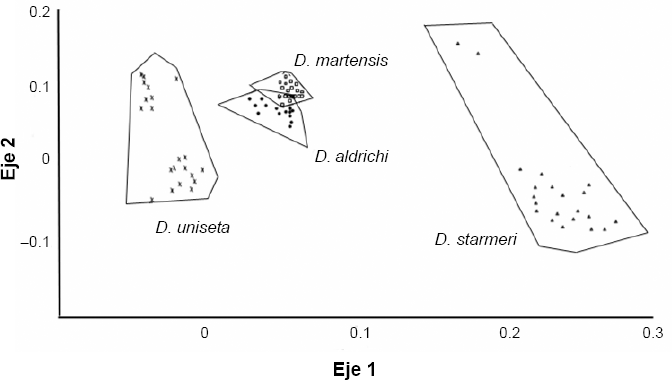
Ordenamiento de los ejes canónicos luego del análisis discriminante, para la forma del aedeago a través del método “outlines” entre Drosophila aldrichi y las tres especies de Camarones: D. uniseta, D. starmeri y D. martensis.
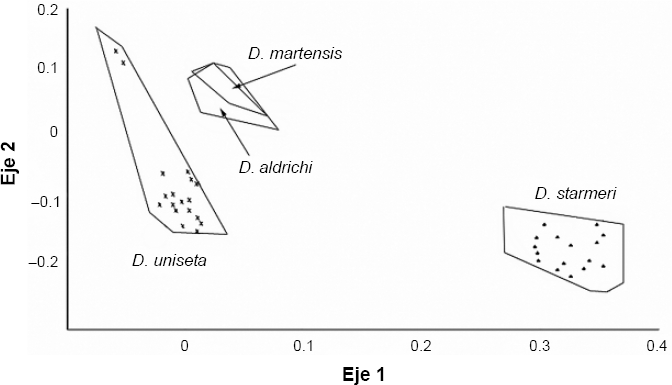
Hubo diferencias estadísticas entre D. aldrichi y las dos especies de Santa Marta (Px wilk= 4,2E-29, Px wilk = 1,052E-9) (Fig. 5), con lo que se puede atribuir la condición de carácter diagnostico de especie al aedeago de los machos. El CVA entre las nueve agrupaciones muestra diferencias significativas entre las poblaciones y entre las especies (P < 0,05) en Wilk 4.23e−163. Las gráficas señalan la gran similitud entre D. martensis y D. aldrichi, al ser comparadas en todas las poblaciones y confirman las diferencias entre estas dos especies y D. uniseta y D. starmeri (Fig. 6).
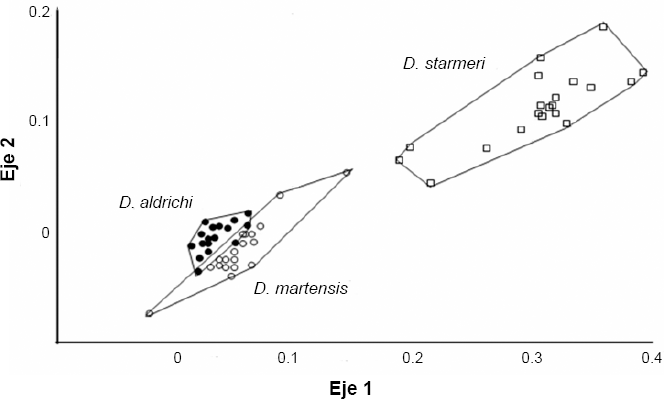
Ordenamiento de los ejes canónicos luego del análisis discriminante, para la forma del aedeago a través del método “outlines” entre Drosophila aldrichi y las dos especies de Santa Marta. (D. starmeri, D. martensis).
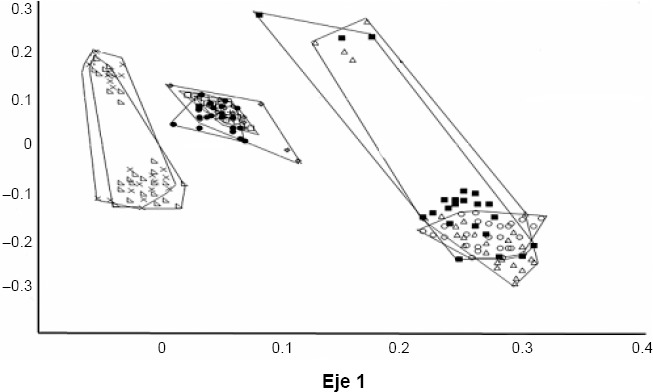
Ordenamiento de los ejes canónicos luego del análisis discriminante, para la forma del aedeago a través del método “outlines” entre las nueve agrupaciones: D. aldrichi (•), D. starmeri Sta. Marta (■), D. martensis Sta. Marta (+), D. uniseta Camarones (x), D. starmeri Camarones; D. martensis Camarones; D. uniseta Tatacoa (△), D. starmeri Tatacoa; MT= D. martensis Tatacoa. Drosophila aldrichi (•) y las tres especies de Camarones: D. uniseta (x), D. starmeri (Δ) y D. martensis (0).
El CVA entre las tres poblaciones de Drosophila martensis: Tatacoa, Camarones y Santa Marta no mostró diferencias significativas entre las poblaciones (Pλ wilk = 0,1816, Pa wilk = 0,1652), lo que indica que para esta especie no hay evidencia de formación de razas, aunque si se empieza a notar una clara distribución de polimorfismos, propios de cada una de las tres poblaciones. La misma prueba en D. starmeri para Tatacoa, Camarones y Santa Marta, permitió establecer diferencias significativas (P < 0,05), 0,0004799 en Wilk, lo cual apoya la hipótesis de formación de razas geográficas. Aunque no hay una total separación de grupos, pues aún se observan traslapes, la diferenciación es más clara que en D. martensis. Las dos poblaciones de D. uniseta: Tatacoa y Camarones, parecen estar en el límite del valor significativo (Px wilk = 0,05079, PA wilk = 0,04878) aunque empieza a notarse una clara diferenciación de polimorfismos propios de cada una de las poblaciones, en las zonas no traslapadas.
Del análisis del contorno de los aedeagos se evidencia la formación de razas geográficas en D. starmeri tendiendo en cuenta que la variación morfológica es un reflejo de la diferente arquitectura genética y de los procesos microevolutivos asociados con las condiciones ecológicas que pueden ser particulares. Precisamente en este caso se compararon poblaciones en ecosistemas totalmente diferentes como lo son poblaciones costeras en el caso de Riohacha y Santa Marta con una diversidad más baja de especies cactáceas y la población del desierto de la Tatacoa en donde las condiciones edáficas y la vegetación cactácea es más diversa. La variación encontrada en el contorno del aedeago de D. starmeri puede ser el resultado de procesos estocásticos o de la acción de la selección natural, sin embargo, antes de determinar cual de estos mecanismos es el responsable de la variación inter-poblacional en esta especie, es patente que la existencia de variación propia en cada población sustentada en cualquiera de las dos hipótesis y en la imposibilidad de ser compartida por el aislamiento geográfico se ajusta a las primeras etapas del modelo de especiación alopátrica el cual incluye la formación de razas geográficas.
En D. uniseta se empiezan a definir polimorfismos propios de cada población al observarse la formación de al menos dos grupos de individuos dentro de cada población. D. martensis y D. aldrichi se sobreponen totalmente como si fueran la misma especie, pero se diferencian perfectamente de D. starmeri y D. uniseta. La alta similitud en la forma del aedeago entre D. aldrichi y D. martensis es muy interesante bajo el contexto planteado por Wasserman y Ruiz (1993), quienes definen a D. aldrichi no como una única especie sino como una categoría “mixed bag” o ensalada de formas homogéneas, pudiendo ser un grupo de especies crípticas al determinarlas por simple inspección, solo distinguibles por pruebas de apareamiento, lo que plantea que se debe hacer un estudio sistemático de su morfología y reproducción, no solo para determinar la naturaleza de la categoría “D. aldrichi” si no también para entender la evolución de las especies de su conglomerado y la relación con las del conglomerado martensis, particularmente con D. martensis.
La evolución requiere la sucesión de una serie de hechos que generalmente se hallan distribuidos en un periodo considerable por lo que resulta difícil determinar todos y cada uno de estos. A pesar de esto, es claro con estos resultados que la forma del eadeago puede constituirse en un carácter clave en el esclarecimiento del proceso evolutivo del grupo repleta de modo que con el modelo biológico utilizado se han encontrado evidencias de este fenómeno que en teoría resulta abstracto. Así mismo, es una herramienta fundamental en la resolución de el problema taxonómico de las especies sinmorficas o crípticas de dicho grupo.
Footnotes
Agradecimientos
Los autores agradecen al Comité Central de Investigaciones de la Universidad del Tolima por la financiación del trabajo.