
Abstract
Summary
The first detection of the spittlebug Prosapia simulans (Walker) (Homoptera; Cercopidae) for Colombia in 1999 was a warning for ranchers and sugar cane growers in the Cauca Valley, where it already has achieved economic levels in Brachiaria. Despite its importance in pastures and cane in Central America, little is known about its biology. In order to obtain the foundation necessary to advance in its management, morphological measurements were used to recognize the life stages. To quantify the life cycle, observations were made of eggs in petri dishes under incubation, and the nymphs and adults on potted host plants. To establish oviposition sites, eggs were recovered from groups of females maintained in pots with different substrates. As with other species of spittlebugs, the eggs of P. simulans passed through four morphologically distinguishable developmental stages, the nymphs passed through five instars, and the adults showed sexual dimorphism. The duration of the egg stage was 18 ± 1,3 days, the nymphs 45,6 ± 5,4 days, and the mean longevity of the adults was 16,5 days, for a total life cycle of 71,9 days. An additional study confirmed a high incidence of diapause in eggs from the field (69,8% in the period of January-May 2000) where egg eclosion was extended up to 128 days. Females exhibited a preference for the stem as an oviposition site with 82,7% of the eggs recovered. In contrast with other species of the spittlebug complex in Colombia, P. simulans has a long life cycle and prefers the stem over soil as oviposition substrate. So far, the presence of P. simulans is limited to four localities of the Cauca Valle.
Keywords
Introducción
En junio de 1999 se detectó por primera vez para Suramérica y Colombia el género Prosapia y la especie P. simulans (Walker) (Rodríguez et al. 2000; Peck et al. 2001). En este estudio P. simulans fue detectado en cuatro municipios en el valle geográfico del río Cauca: Santander de Quilichao (Cauca), Cerrito, Calima el Darién y Yotoco (Valle del Cauca). Estas poblaciones se describieron como persistentes y abundantes, alcanzando niveles económicamente importantes en Brachiaria decumbens (Peck et al. 2001). P. simulans probablemente provino de Centroamérica donde es una plaga importante de forrajes y caña de azúcar.
El mión o salivazo de los pastos es un insecto chupador que se alimenta exclusivamente de la savia de las plantas, particularmente del xilema (Fewkes 1969). Las ninfas en la familia Cercopidae se caracterizan principalmente por la masa de espuma que la ninfa construye en su sitio de alimentación sobre el hospedero, sirviendo de defensa de enemigos naturales y protección en condiciones climáticas adversas (Whittaker 1970). A diferencia de las ninfas, los adultos no producen espuma, empleando como mecanismo de defensa su habilidad para saltar, apoyado adicionalmente por el aposematismo y un mecanismo secundario de defensa denominado autohemorragia (Peck 2000a).
De las aproximadamente 360 especies de cercópidos registrados en el Neotrópico, 20-30 son plagas de gramíneas, principalmente de los géneros Aeneolamia, Deois, Mahanarva, Prosapia y Zulia (Peck 1999). Están ampliamente distribuidas geográficamente y altitudinalmente en gramíneas del neotrópico, desde el sur de los Estados Unidos hasta el norte de Argentina, y desde el nivel del mar hasta los 3.000 msnm. La diversidad taxonómica del grupo se expresa en Colombia donde se han registrado 15 especies de seis géneros: Aeneolamia bogotensis (Distant), A. lepidior (Fowler), A. reducta (Lallemand), A. varia (Fabricius), Mahanarva sp. nov., M. andigena (Jacobi), M. phantastica (Breddin), Notozulia entreriana (Berg), Prosapia simulans (Walker), Sphenorhina rubra (L.), Zulia birubromaculata (Lallemand), Z. carbonaria (Lallemand), Z. pubescens (F.) y Zulia sp. nov. (Peck 2001).
El salivazo es considerado la plaga más limitante en la producción ganadera de América tropical (Lapointe y Miles 1992; Valerio et al. 2001). Un complejo diverso de 11 géneros nativos ataca gramíneas cultivadas, que incluye casi todos los géneros importantes de forrajes (Bothriochloa, Brachiaria, Cynodon, Panicum, Pennisetum) (Peck 2001), al igual que la caña de azúcar (Fewkes 1969; Guagliumi 1972) y bajo ciertas circunstancias arroz y césped (Nilakhe 1985; Tashiro 1987). Por varias razones aún no se cuenta con una efectiva estrategia de MIP (manejo integrado de plagas) para esta plaga en forrajes tropicales (Peck 2000b). La diversidad taxonómica del grupo implica que la biología y las tácticas de manejo apropiadas varían de una región a otra, entonces el manejo de la plaga es afectado por una simplificación del problema dado el rango de variación en las asociaciones salivazo/ forraje/hábitat.
Se estima que 52,4% de la superficie aprovechable en Colombia está dedicado a pastos (DANE 1998). Dentro de éstos, el género Brachiaria, particularmente B. decumbens Stapf., es la más difundida en América tropical con un área aproximada de 40 millones de ha (Pizarro et al. 1998). La difusión de este cultivar se debe a sus características agronómicas que le confieren adaptabilidad a suelos ácidos e infértiles, altos rendimientos, tolerancia al pastoreo intensivo y el pisoteo de los animales (Keller-Grein et al. 1996). Sin embargo, sufre una desventaja comparativa con relación al salivazo de los pastos por su susceptibilidad alta al ataque del insecto. En la actualidad, Colombia cuenta con un área aproximada de 4,720,000 ha. susceptibles al ataque del salivazo en el trópico seco y 1,140,000 ha. en el trópico húmedo, con una disminución en los ingresos del productor de US $8.5-34.0 millones anuales atribuidos al daño causado por el salivazo (Holmann y Peck, manuscrito sometido).
A pesar de su importancia económica, existe poca información biológica sobre especies del salivazo. En Colombia, el Centro Internacional de Agricultura Tropical (CIAT) ha venido estudiando la biología, comportamiento y distribución de varias especies para llenar este vacío y proveer datos fundamentales para lograr avances en el manejo integrado de la plaga. La implementación exitosa de tácticas de manejo depende de información específica del complejo de especies y del hábitat (estacionalidad, hospederos, sistema de manejo de pradera y producción), en la región donde se quiere ejercer un control. Además, requieren un conocimiento detallado de aspectos como reconocimiento de los estados de vida, duración del ciclo de vida, fenología, enemigos naturales, y preferencias de hospedero y hábitat.
Prosapia simulans, además de ser una nueva plaga en B. decumbens para Colombia, se presenta como plaga potencial de la caña de azúcar. Esta especie es una plaga muy dañina en cultivares de caña de azúcar en Centroamérica (Peck et al. 2001). Aunque hasta ahora P. simulans no ha sido detectada atacando este cultivo en Colombia, podría amenazar la producción coincidente con la prohibición de las quemas. La prohibición de las quemas en una región de Brasil causó la emergencia de una nueva plaga de salivazo en caña que anteriormente no sufría por esta clase de plagas (Botelho et al. 2000).
Con el propósito de solucionar la carencia de información específica de P. simulans en el Valle del Cauca, se realizaron una serie de estudios para contribuir a la fase diagnóstica. Esto incluye una descripción de los estados de desarrollo de los huevos, ninfas y adultos, duración de los mismos y preferencias en los sitios de oviposición. Además, se estudió el patrón de eclosión de los huevos obtenidos del campo para confirmar la diapausa, y se determinó la distribución geográfica actual. Esta información ofrece los fundamentos biológicos necesarios para avanzar en el MIP de P. simulans en gramíneas cultivadas de Colombia.
Materiales y Métodos
Origen y manejo del insecto
Los adultos de P. simulans se recolectaron en pasturas de B. decumbens localizados en la vereda La Primavera, municipio Calima el Darién, departamento del Valle del Cauca a 1.621 msnm (Hacienda La María, 3.940°N, 76.438°W). Los estudios biológicos se realizaron en las instalaciones del CIAT, Palmira, Valle del Cauca, localizado a 3.517°N y 76.367°W, altitud 965 msnm, precipitación anual 1.000 mm, y temperatura promedia de 24°C.
Con el propósito de contar con todos los estados de desarrollo del insecto y superar su estacionalidad en el campo, se implementó una colonia en casa de malla según la metodología de Lapointe et al. (1989) y Rodríguez et al. (2002). Los adultos se mantuvieron en una cámara de oviposición constituida por una jaula de madera (40 x 40 x 80 cm), piso removible con sustrato de oviposición (barro de 1 mm de espesor) y aberturas laterales para el alimento. En la colonia y los demás estudios se usó B. ruziziensis Germain y Everard (CIAT 654). Los huevos se separaron del sustrato por flotación en agua saturada con sal y se almacenaron en cajas de petri sobre papel filtro húmedo hasta la eclosión. Las nuevas ninfas se trasladaron en grupos de 15 a materas plantadas con B. ruziziensis preparadas especialmente para alta disponibilidad de raíces superficiales (sitios de alimentación), y cubiertas con una tapa de aluminio pintada de blanco provista de un orificio para darle paso al material vegetal. Estas condiciones aseguraron un microclima favorable para el desarrollo de las ninfas.
De acuerdo con protocolos establecidos anteriormente por Rodríguez et al. (2002), se realizaron mediciones de ciertas características externas para caracterizar los diferentes estados de desarrollo. Para los huevos se midió el largo y ancho máximo de las cuatro fases de desarrollo. Para las ninfas se midió el ancho de la cápsula cefálica, largo del estilete (punto distal del labro al punto distal del labio), largo del cuerpo y largo del muñón alar anterior de los cinco ínstares y para machos y hembras del ínstar Vb próximo a pasar a la fase adulta. Para los adultos se midió el ancho del cuerpo, largo del cuerpo con y sin ala, largo del ala anterior y largo del estilete de ambos sexos. Para la caracterización morfológica de cada uno de los estados, se emplearon 100 huevos de hembras provenientes de la colonia, 40 ninfas de la colonia, y 40 adultos recolectados directamente del campo. Las mediciones se realizaron mediante un microscopio estereoscopio acondicionado con una reglilla ocular.
Ciclo de vida
Se estableció la duración promedia de cada una de las fases de desarrollo según metodologías establecidas previamente (Rodríguez et al. 2002). Para los huevos, se emplearon 100 huevos de <12 horas de edad, provenientes de grupos de hembras diferentes, y evaluados diariamente mientras se mantenían sobre papel filtro húmedo en cajas de petri bajo condiciones controladas (27°C, 100% HR, oscuridad total).
Para las ninfas se emplearon 40 ninfas de <12 horas de edad para infestar en materas individuales establecidas como en la descripción de la colonia. El paso de un ínstar a otro se determinó mediante observación directa de la ninfa o de la exuvia. Para los adultos se emplearon 40 adultos tenerales <12 horas de edad provenientes de la colonia, colocando cuatro individuos no sexados por matera preparada con la planta huésped, bajo un cilindro de acetato transparente. Este cilindro se cubrió en la parte superior con un trozo de muselina para evitar el escape de los individuos. Para facilitar la evaluación de los adultos, se adicionó una capa de arena blanca en la superficie del sustrato la cual facilitó la ubicación de los adultos en el piso. Al retirar los adultos muertos se procedió a determinar su sexo. Se calculó la longevidad promedio para cada una de las cuatro fases de desarrollo de los huevos, los cinco ínstares y los sexos de los adultos.
Sitios de oviposición
Para determinar la preferencia por sitio de oviposición con relación al tipo de sustrato, se simularon condiciones de campo en casa de malla de acuerdo con metodologías ya establecidas (Rodríguez et al. 2002). A materas plantadas con B. ruziziensis, se adicionó una capa de suelo mullido de 2,0 cm de espesor sostenida por una tapa de aluminio invertida. Se cubrió la mitad de la superficie con 2.0 g de hojarasca seca (porciones de 1-2 cm de largo de hojas y tallos de B. ruziziensis). Se colocaron seis adultos tenerales (cuatro hembras) <12 horas de edad en cada una de las 10 materas bajo el cilindro de acetato. A los 10 días se recuperaron los huevos de los cuatro sustratos: suelo bajo hojarasca, suelo descubierto, hojarasca y sobre la planta. Los huevos depositados en el suelo se recuperaron por flotación, y los demás mediante inspección directa del material con la ayuda del estereoscopio.
Incidencia de diapausa
Se realizó un estudio para confirmar la diapausa en los huevos de P. simulans provenientes del campo. Paralelo a un estudio sobre la fluctuación poblacional de P. simulans en el Valle del Cauca (Hda. Piedechinche, vereda Santa Helena, municipio El Cerrito) (CIAT 2001), cada semana se colectó un grupo de 2 hembras de cada mitad de cinco parcelas de muestreo de B. decumbens. Se les colocaron a ovipositar en cajas petri con papel filtro húmedo mantenidos bajo condiciones controladas. Se cuantificaron los días hasta eclosión para determinar la incidencia y duración de la diapausa durante el período enero-mayo de 2001.
Distribución geográfica
Se reunió la información sobre la distribución geográfica conocida de P. simulans desde su primer registro en el país en 1999. Para esto, se registró su presencia o ausencia en muestreos de pasturas en diferentes zonas del país con varios grupos de colaboradores. Se incluyeron los registros de ausencia solamente si otras especies del complejo se encontraron en el muestreo específico (indicando hábitat apropiado).
Análisis
Los datos registrados durante los ensayos se agruparon en valores mínimos, máximos y promedios con el error estándar como medida de variación. Todos los análisis estadísticos se realizaron utilizando el programa SAS y la prueba de comparación múltiple Tukey-Kramer HSD al P<0.05. Para el caso específico de la longevidad de los adultos, se empleó la distribución de Weibull (Pinder et al. 1978) para establecer la longevidad media de la población y de los sexos.
Resultados
Huevo
Los huevos de P. simulans pasaron por cuatro fases de desarrollo (S1, S2, S3, S4, sensu Fewkes 1969 y Wiedijk 1982), plenamente distinguibles por su apariencia externa y tamaño. En la fase S1, los huevos fueron de color amarillo y sin evidencia de desarrollo. En la fase S2, había una mancha roja localizada debajo de la línea de eclosión. En la fase S3, la línea de eclosión estaba abierta y la superficie negra del opérculo estaba expuesta. En la fase S4, finalmente había un par de puntos rojos a los costados del corión, y otra pareja al lado de la línea de eclosión que representaban los ojos. El tamaño de los huevos, tanto ancho como largo, se incrementó significativamente al pasar de una fase a otra (Tabla 1).
Tamaño (mm) de los huevos del salivazo P. simulans según fase de desarrollo (promedios ± E.E., rango)
Para cada medida, promedios seguidos por letras distintas son estadísticamente diferentes al P<0.05.
La duración promedio del estado de huevo fue de 18 días dividido en 6,9, 2,1, 4,0 y 5,2 días para cada una de las fases de desarrollo, que correspondieron a 38,3, 11,7, 22,2 y 28,9% de la duración total del huevo, respectivamente (Tabla 2).
Duración (días) de los huevos de P. simulans según fase de desarrollo
Promedios seguidos por letras distintas son estadísticamente diferentes al P<0.05.
Ninfa
En cada uno de los parámetros morfológicos evaluados, hubo un incremento significativo en el tamaño de un ínstar a otro (Tabla 3). En contraste, a lo largo del cuerpo y estilete no hubo traslape para el ancho de la cápsula cefálica entre los ínstares. El ínstar Vb superó al ínstar Va en todos los parámetros. A nivel de sexo, las hembras del ínstar Vb superaron a los machos del ínstar Vb en cada uno de los parámetros. La duración promedio para la fase ninfal fue 45,6 días dividido en 6,7, 7,5, 9,3, 10,0 y 13,1 días para cada uno de los ínstares, respectivamente, que correspondieron a 14,7, 16,4, 20,4, 21,9 y 28,7% de la duración total de la ninfa, respectivamente. La duración del ínstar V sobrepasó los demás ínstares, y fue más corto para el ínstar I y II (Tabla 4).
Tamaño (mm) de los estados de vida ninfal y adulto de P. simulans (promedio ± E.E., rango; n=40)
Para cada columna, promedios seguidos por letras distintas son estadísticamente diferentes al P< 0.05.
Duración (días) de la fase ninfal de P. simulans según ínstar (promedio ± E.E., rango; n=40)
Promedios seguidos por letras distintas son estadísticamente diferentes al P< 0.05.
Adulto
Con excepción del largo del ala anterior, las hembras superaron a los machos en cada uno de los parámetros morfológicos evaluados (Tabla 3). Con respecto a los sexos del ínstar Vb, los adultos presentaron un incremento en el ancho de la cápsula cefálica y largo del ala anterior, pero una disminución en el largo del cuerpo y estilete. La longevidad media para adultos de P. simulans fue 16,5 días, con una longevidad media de 15,3 días para los machos y 17,9 días para las hembras, superando a los machos en 2,6 días. Bajo las condiciones del ensayo, la duración total del ciclo de vida para P. simulans fue 71,9 días (18,0 + 45,6 + 8,3, huevo + ninfa + adulto).
Las hembras de P. simulans presentaron una preferencia marcada por la superficie de la planta como sustrato de oviposición; 82,6% (8,7 20,5%) de los huevos se recuperaron de este sustrato. Solamente 17,4% (2,0 4,6%) fue recuperado del suelo con 3,6% (0,5 3,1%) en el suelo descubierto y 13,8% (2,0 4,6%) bajo la hojarasca.
Incidencia de diapausa
De los 1.249 huevos de P. simulans obtenidos en las ocho fechas de evaluación, hubo una mortalidad de 5,5% y una incidencia de diapausa de 69,8% (Tabla 5). Con excepción de dos fechas de recolección (22 marzo, 31 mayo), la proporción de diapausa en los huevos superó 70%. Los huevos eclosionaron durante un período de 18-128 días (Fig. 1). El tiempo promedio de eclosión fue 24 días para los huevos no diapáusicos (definido como los que eclosionaron dentro de los 30 días) y 79,5 para los huevos diapáusicos. Se pudo establecer que fue la fase S2 la que se extendió en el caso de diapausa.
Incidencia y tiempo de eclosión de huevos no diapáusicos y diapáusicos de P. simulans en Piedechinche, Departamento del Valle del Cauca (2001)
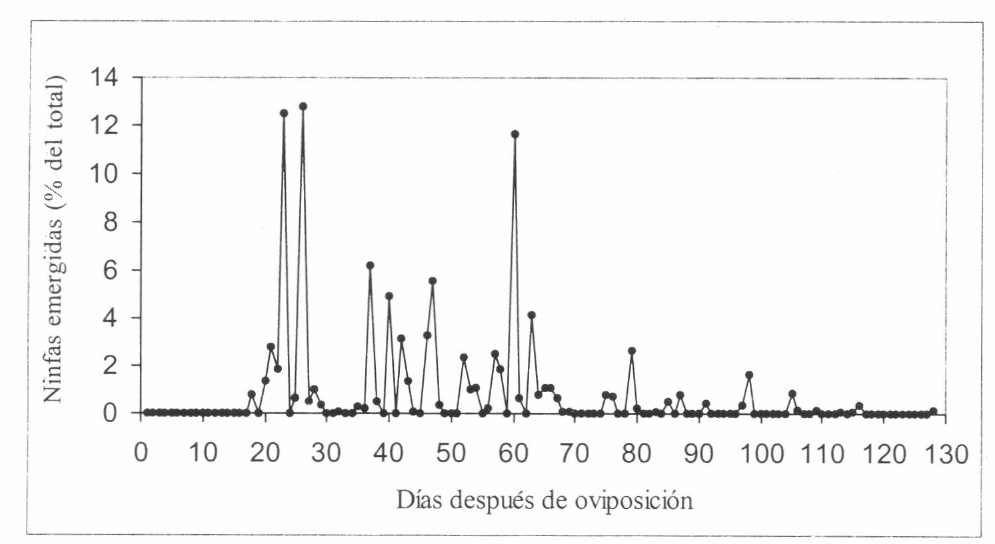
Patrón de eclosión de huevos de P. simulans recolectados en ocho fechas durante el período enero a mayo 2001 en el municipio de Cerrito, departamento del Valle del Cauca.
Distribución geográfica
Hasta la fecha P. simulans no ha sido detectado fuera de los departamentos de Cauca y Valle del Cauca (Tabla 6). Las únicas poblaciones persistentes conocidas están situadas en cuatro localidades del Valle del Cauca. Estos son los municipios de Cerrito (vereda Sta Helena, 1.115 y 1.100 msnm), Yotoco (vereda Cordobitas, 1.535 msnm), Calima el Darién (vereda La Primavera, 1.621 msnm y vereda Diamante la Gaviota, 1.575 msnm). En estos sitios P. simulans estaba asociado con seis hospederos diferentes: Axonopus micay, B. decumbens, B. dictyoneura, Cynodon plectostachys, Dichromena ciliata, Hyparrhenia rufa y Sacharum officinarum. El único otro registro fue el municipio de Santander de Quilichao (Hda. Las Palmas, 1.060 msnm) consistiendo en una sola hembra. Esta especie no ha sido detectada hasta ahora en muestreos del salivazo en la costa caribe (Sucre), el piedemonte Amazónico (Caquetá), el piedemonte de la Orinoquia (Meta), la costa pacífica sur (Nariño), el valle sur del río Magdalena (Tolima) u otros sitios en el valle del río Cauca y zona cafetera.
Distribución conocida de P. simulans en Colombia
En los sitios negativos se encontraron especies del complejo salivazo más no P. simulans.
Discusión
Los resultados obtenidos para P. simulans fortalecen ciertos patrones morfológicos comunes a otras especies del salivazo estudiadas. Primero, los huevos pasan por las cuatro fases de desarrollo morfológicamente distinguibles, con un incremento en tamaño a medida que se desarrollan, principalmente entre las fases S2 y S3 donde el huevo abre el corion facilitando la absorción de agua. Segundo, las ninfas pasan por cinco ínstares distinguibles por el incremento significativo en tamaño de parámetros físicos. Además, se confirmó que el ancho de la cápsula cefálica es la característica morfológica más diagnóstica para separar los ínstares, debido al no traslape entre los rangos obtenidos para cada ínstar, a diferencia de los otros parámetros morfológicos evaluados. Además, se confirmó para P. simulans la diferenciación entre el ínstar Va y el Vb (próximo a transformar en adulto). En el ínstar Vb, además de la espina lateral y la corona de espinas en la metatibia de la ninfa, asociadas a éstas se evidencian las espinas y coronas oscuras del adulto próximo a salir. Tercero, se confirmó un polimorfismo sexual marcado en los adultos expresado por el tamaño mayor de las hembras para cada uno de los parámetros, excepto para el largo del ala anterior. Cuarto, se evidenció una disminución en el tamaño del estilete del ínstar V al pasar al estado adulto. Estas observaciones representan patrones para el complejo salivazo de los pastos establecidos en otros estudios sobre el género Zulia (Rodríguez et al. 2002), otras especies en Colombia (Ballesteros y Gallego 1999; CIAT 2001; Peck et al. manuscrito sometido) y especies en otros países (Peck 1998).
Los estudios biológicos realizados a P. simulans amplían la variación conocida para el complejo salivazo de los pastos en Colombia. La duración del estado de huevo (18,0 días) supera lo referenciado para Aeneolamia (14,1 17,2 días), Mahanarva (16,417,0 días), y Zulia (14,6 - 17,4 días). En el caso de las ninfas, la duración (45,6 días) supera a Aeneolamia (26,1 35,4 días) y Zulia (38,042,7), pero es parecida a la duración de Mahanarva andigena (46,5 días). Para los adultos la duración (16,5 días) supera a Aeneolamia (6,2-7,2) y Mahanarva sp. nov. (6, 8 días), es parecido a Zulia (14,2 19,6) e inferior a M. andigena (24 días). Con relación a la duración total del ciclo de vida, P. simulans (71,9 días) supera a Aeneolamia (45,3 52,6), Mahanarva sp. nov. (64,6), Zulia (61,569,6) y es inferior a M. andigena (74,9 días) (CIAT 1998, 2001; Ballesteros y Gallego 1999; Rodríguez et al. 2002; Peck et al. manuscrito sometido).
Además, P. simulans amplía la variación exhibida por otras especies de salivazo estudiadas anteriormente en Colombia con respecto al tipo de sustrato de oviposición. P. simulans mostró preferencia por la superficie de la planta (82,6%) como sustrato de oviposición y solamente M. andigena (32,4) y Z. pubescens (59,2%) alcanzan una proporción significativa en el mismo sustrato (CIAT 2001; Rodríguez 2001). Las demás especies estudiadas en Colombia colocan 74 100% de los huevos en el suelo (CIAT 1998; Ballesteros y Gallego 1999; Rodríguez et al. 2002; Peck et al. manuscrito sometido).
La información biológica sobre P. simulans en América Central es limitada porque la mayoría fue obtenida bajo condiciones de campo donde P. simulans ocurrió junto con otras especies del salivazo. Oomen (1975) señala una duración para los estados de huevo y ninfa de 58,0 días sobre B. decumbens en México, dividido en 33,5 y 25,5 días para los huevos y ninfas, respectivamente. Velasco y Sifuentes (1970) de igual forma indican, para los mismos estados de desarrollo, una duración de 58,4 días, dividido en 18,7 y 2248 días, para huevos y ninfas, respectivamente.
La nueva información obtenida en estos estudios ofrece los fundamentos iniciales para caracterizar P. simulans. Avances en su manejo como plaga dependerán de otros estudios para fortalecer el conocimiento de su asociación con gramíneas forrajeras en Colombia. Como ejemplo, estudios adicionales sobre la dinámica poblacional y fenología podría dar luz sobre la alta incidencia de diapausa durante el año aún durante épocas de alta humedad. Además, todavía hace falta (1) información sobre aspectos de la biología reproductiva como período de preoviposición y tasa de oviposición, (2) realizar estudios para establecer la susceptibilidad de la caña de azúcar al ataque del insecto, y (3) monitoreos en campo más detallados para establecer la distribución actual y monitorear su difusión futura hacía otras regiones y otros cultivos como la caña de azúcar.
Conclusiones
Igual que otras especies, es posible distinguir con precisión los estados de desarrollo de P. simulans (cuatro fases de desarrollo en los huevos, cinco ínstares ninfales y dos sexos en los adultos) basados en características morfológicas. Además, en el caso de las ninfas el ancho de la cápsula cefálica es el carácter más diagnóstico para diferenciar los ínstares y los adultos exhiben un dimorfismo sexual expresado por el mayor tamaño de las hembras.
Para P. simulans la duración total de la fase de huevo, ninfa y adulto es 18,0, 45,6 y 16,5 días, respectivamente, para un ciclo de vida 71,9 días. La diapausa se exhibe en la fase del huevo extendiendo el desarrollo hasta 128 días. Para el período enero-mayo, la incidencia de diapausa fue 69,8% en el municipio de Santa Helena (Valle del Cauca).
Las hembras de P. simulans exhiben poca flexibilidad por el tipo de sustrato al colocar el 82,6% de los huevos en la superficie de la planta.
Hasta la fecha no se documenta la presencia de P. simulans fuera de los departamentos del Cauca y Valle del Cauca.
Agradecimientos
Se agradece a la Hacienda Piedechinche (Santa Helena) y Finca La María (Calima Darién) por facilitar sus predios, y a Oscar Yela por su colaboración y mantenimiento de la colonia de P. simulans. Este trabajo se realizó gracias a la ayuda financiera de la Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República y aporte parcial del Programa Nacional de Transferencia de Tecnología Agropecuaria (PRONATTA).