
Abstract
The purpose of this study was to determine resistance levels to temephos and deltametrine in two natural populations of Aedes aegypti from Peru. Bioassays in larvae and adults were carried out following the methodology of the World Health Organization. The visualization of B-esterase bands was made by poliacrylamide gel electrophoresis (PAGE) in fourth instar larvae. Natural populations from Sullana (Piura) and El Porvenir (La Libertad) presented resistance ratios to temephos of 1.67 X and 1.45 X, respectively, indicating susceptibility in both populations; for deltametrine the Sullana population presented resistance with a KDT50 of 106 min and 68% mortality at 24 h; in the El Porvenir population susceptibility was observed, with a KDT50 of 36.9 min and 99% mortality. Esterase B2 was identified with a Rf of 0.23 only in El Porvenir population. The insecticide temephos can continue being used in control programs of the Aedes aegypti vector because the species is still susceptible to this organophosphorate; on the contrary with deltametrine, where its effects should be evaluated in natural populations since some of them should be presenting resistance, as in the case of the Sullana population.
Keywords
Introducción
Aedes aegypti (Linnaeus) es el vector del virus del dengue en las Américas, se distribuye usualmente entre las latitudes de 35° LS y 35° LN; pero ha sido encontrado hasta una latitud de 45° LN (Bisset 2002). En el Perú, fue detectado por primera vez en 1852, se cree que éste ingresó por el norte, desde Guayaquil-Ecuador, para luego establecerse progresivamente a lo largo de la costa norte y central peruana, extendiendo su distribución hasta Tacna (Sevilla et al. 2001).
En el Perú, frente a la presencia de Ae. aegypti, el Programa de Control Vectorial del Ministerio de Salud ha hecho uso del larvicida organofosforado temefos y nebulización espacial de piretroides como el adulticida deltametrina (CICE 2000; Vargas et al. 2003).
Estudios sobre el uso indiscriminado de insecticidas indican que en 1947 se registró el primer caso de resistencia a DDT en Ae. taeniorhynchus (Wiedemann) y Ae. sollicitans (Walker) (Brown 1986); en poblaciones de A. aegypti de Venezuela y Cuba se ha indicado la resistencia a temefos pirimifos metil, clorpirifos y cipermetrina (Bisset et al. 2001). Asimismo, la resistencia a malatión, diazinón y DDT ha sido registrada en Brasil y en Puerto Rico (Campos y Andrade 2001).
Se ha demostrado que en Ae. aegypti, la capacidad de resistir al malatión está asociada a la destoxificación mediada por enzimas de actividad específica: carboxilesterasas y el incremento de los niveles de resistencia a temefos está presuntamente asociado con este mecanismo, sólo que las enzimas destoxificadoras son fosfatasas (Bisset et al. 2001). Existen otros mecanismos de resistencia en Ae. aegypti, como son el gen kdr, basado en la insensibilidad en el sitio de acción responsable de la resistencia cruzada a DDT y piretroides, la enzima glutatión-s-transferasa, responsable de la resistencia a organofosforados y DDT, finalmente la familia de oxidasas del citocromo P-450 que interviene en la resistencia a organofosforados, piretroides y DDT (Bisset et al. 2001; Bisset 2002).
Teniendo en cuenta que Ae. aegypti es una especie que se ha establecido paulatinamente en el Perú, ocasionando numerosos casos de dengue en el año 2002, es que el presente trabajo tuvo como objetivo determinar los niveles de resistencia al larvicida temefos y adulticida deltametrina en dos poblaciones naturales de Aedes aegypti del norte peruano, colectadas durante el año 2002.
Materiales y Métodos
Material Biológico
Para el trabajo se utilizaron 2 poblaciones naturales de Aedes aegypti, una del distrito de Sullana (Piura) y la otra del distrito El Porvenir (La Libertad) y una cepa de referencia Rockefeller, suministrada por el Instituto de Medicina Tropical “Pedro Kouri” (IPK) La Habana-Cuba. Las colonias se establecieron y mantuvieron en el insectario del Instituto de Investigación en Microbiología y Parasitología Tropical (INIMYPAT) con temperatura de 26 ± 2°C y humedad relativa de 65%.
Insecticidas
El insecticida temefos al 93,3% de pureza, fue suministrado por el Instituto de Medicina Tropical “Pedro Kouri” La Habana-Cuba. Los papeles impregnados con deltametrina al 0,025%, fueron suministrados por el Centro de Investigación de Paludismo, Tapachula, Chiapas-México.
Bioensayo de resistencia y/o susceptibilidad a insecticidas en larvas
Los bioensayos se realizaron siguiendo la metodología de la Organización Mundial de la Salud (WHO 1981). Se utilizaron larvas del tercer o cuarto estadío temprano de cada población incluyendo la cepa de referencia Rockefeller. Se emplearon 5 réplicas de cada concentración del insecticida (20 larvas por réplica) produciéndose mortalidades entre 2 y 98%. Todas las soluciones se ajustaron a un volumen final de 1 ml con acetona. Esta concentración de acetona no causó mortalidad en los controles. La lectura de las mortalidades se realizó a las 24 h y los resultados se procesaron mediante el programa probit-log (Raymond 1985) para obtener la CL50 y CL95.
Bioensayos de resistencia y/o susceptibilidad a insecticidas en adultos
Los bioensayos se hicieron siguiendo la metodología de la Organización Mundial de la Salud (WHO 1981). Grupos de 18-25 mosquitos adultos hembras se expusieron a papeles impregnados con deltametrina 0,025% en tubos patrones de prueba de la WHO (12 x 4 cm) a 23 ± 2°C y 70% de humedad relativa. El tiempo de exposición fue de 1 h, el número de mosquitos muertos al contacto con el papel impregnado con insecticida se registró a los 10, 20, 30, 40, 50, 60, 70 y 80 min, después se transfirieron a un tubo vacío limpio, provisto con un algodón humedecido con agua azucarada, y se mantuvieron hasta las 24 h para realizar la lectura de mortalidad, donde 100 - 98% de mortalidad indica susceptibilidad, 97–80% de mortalidad indica que se debe realizar confirmación de resistencia y menos de 80% de mortalidad señala resistencia (WHO 1998). Se realizaron 5 réplicas para cada población. Los controles se expusieron a papeles impregnados con aceite de oliva, los resultados se procesaron mediante el programa probit-log (Raymond 1985) para obtener KDT50 y KDT90.
Determinación del factor de resistencia
Se determinó mediante la fórmula (Bisset et al. 1996):
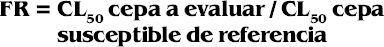
FR < 5 = Susceptible
FR > 5 = Resistente
Electroforesis en gel de poliacrilamida (PAGE)
Se realizó electroforesis en gel de poliacrilamida (9%). Se homogeneizó cada larva con 200 ul de buffer fosfato 0,01M, PH 7,5. En tubos Eppendor (1,5 ml) se adicionaron 15 ul del homogeneizado más 10 ul del indicador azul de bromofenol. Se aplicaron 20 ul de esta mezcla en el gel y se corrió a 150V, durante 45 min. Para la tinción de las bandas de esterasas, se sumergió el gel en 50 ml de buffer fosfato (0,1M, PH 7,5), que contenía 4 ml cada uno del sustrato inespecífico ß-naftilacetato. Después se añadieron 2 ml de colorante Fast Garnet, disuelto previamente en buffer fosfato 0,1M, PH 7,5. Para observar el nivel de migración de las bandas de esterasas en el gel, se determinó la movilidad relativa de las mismas, mediante la fórmula: M. R. = Distancia recorrida de la enzima / Distancia total del gel (Murray et al. 1997).
Resultados y Discusión
Los resultados obtenidos en los bioensayos en larvas (Tabla 1) indican valores de factores de resistencia de 1,67 X y 1,45 X para las poblaciones de Sullana y El Porvenir frente a temefos respectivamente, el resultado encontrado en la población El Porvenir es similar al hallado para esta zona 1,93 X (Vargas et al. 2003). Se registraron altos niveles de resistencia a temefos en poblaciones de Venezuela y Cuba con un FR50 de 11,1 X y FR50 de 5,9 X, respectivamente (Bisset et al. 2001), una población de Brasil presentó un FR50 de 6,3 X (Campos y Andrade 2001); poblaciones de Venezuela manifestaron susceptibilidad con FR50 de 3,9 X y 2,58 X (Bisset et al. 2001), en Brasil una población manifestó un FR50 de 2,7 X (Campos y Andrade 2001). La explicación de estos valores altos, 11,1 X, 6,3 X y 5,9 X, es que a partir de 1977-1980 apareció la epidemia del dengue 1 en el Caribe produciéndose luego la fiebre del dengue hemorrágico con su primera gran epidemia en Cuba en 1981 (Schneider y Droll 2001), por lo que se realizaron campañas de erradicación del vector donde se emplearon insecticidas, ocasionando como consecuencia los primeros casos de resistencia. En el Perú, dengue 1 aparece recién en Loreto en 1990 (Schneider y Droll 2001); haciéndose uso de temefos para el control de Ae. aegypti en los años 1999 y 2000 cuando se tiene un gran número de casos de dengue en el norte peruano (CICE 2000; Vargas et al. 2003); es probable que esta diferencia se pueda asociar a la baja presión de selección ejercida en el norte peruano, comparada con el extensivo uso de temefos en países como Cuba.
Concentraciones letales 50 y 95%, valor de la pendiente de regresión y factor de resistencia a temefos para larvas de Aedes aegypti de las poblaciones colectadas en el año 2002 en Sullana (Piura) y El Porvenir (La Libertad) comparado con la cepa de referencia Rockefeller
Concentración en ppm. Intervalo de confianza entre paréntesis.
Pendiente. Desviación estándar (± DE) entre paréntesis.
Factor de resistencia.
Cepa de referencia.
El KDT50 encontrado para Ae. aegypti de la población Sullana es 106,31 min, lo cual indicaría la evolución de su resistencia a deltametrina (Tabla 2), que comparado con otras especies fue superior, como en Anopheles gambiae (Giles) de África de la localidad de Kao Koffikro que presentó un KDT50 de 78,7 min para este insecticida (Chandre et al. 1999) y Aedes albopictus (Skuse) que presentó un KDT50 aproximado de 53,66 min frente al piretroide cipermetrina (Sulaiman et al. 2002). Asimismo, en trabajos realizados en Malasia con los piretroides cipermetrina y ciflutrina en Ae. albopictus, la especie presentó susceptibilidad con porcentajes de mortalidad de 97,1 y 97,2%, respectivamente (Sulaiman et al. 2002), semejantes a los resultados encontrados en la población El Porvenir con un porcentaje de mortalidad de 99% frente a deltametrina. La resistencia a deltametrina presentada en la población Sullana, es el resultado de la presión de selección por el empleo de aspersiones de deltametrina en viviendas y, otra causa probable en forma indirecta de la presencia de resistencia de Ae. aegypti es debido a los insecticidas empleados para el control de Anopheles albimanus (Wiedemann), principal vector de la malaria en el norte del Perú, ya que las viviendas se encuentran muy cercanas a los campos de cultivos, presentando esta especie, en la actualidad, resistencia a alfacipermetrina, lambdacialotrina, ciflutrina y deltametrina (CICE 2000).
Porcentaje de mortalidad, tiempo “knockdown” (KDT50 y KDT95), valor de la pendiente de regresión y estado de resistencia en poblaciones coleccionadas en el 2002 de Aedes aegypti (adultos) usando tubos de prueba de la OMS
Tiempo en minutos
Pendiente. Desviación estándar (± DE) entre paréntesis.
Número de mosquitos hembras utilizados para los bioensayos.
Cepa de referencia.
En las tablas 1 y 2 se aprecia un valor mayor de la pendiente en las poblaciones El Porvenir y la cepa Rockefeller, siendo estas poblaciones más susceptibles a temefos y deltametrina que la población Sullana, por lo que cuanto más bajo sea el valor de la pendiente de la regresión, más heterogénea será la población y presentará una tendencia mayor a la resistencia (Bisset et al. 2001).
La banda que se distingue en la figura 1 en el gel de poliacrilamida (PAGE) pertenece sólo a la población El Porvenir, identificándose a la esterasa B2 con una movilidad relativa aproximada de 0,23, lo cual concuerda con lo señalado en trabajos previos para la población de El Porvenir, indicándose además la presencia de una esterasa A, (Vargas et al. 2003). El resultado obtenido difiere con el tipo de esterasas detectadas en poblaciones de Venezuela y Cuba en donde el 98% de la poblaciones de Ae. aegypti estudiadas predomina la esterasa A14 con una movilidad relativa de 0,779 (Bisset et al. 2001), la cual estaría produciendo resistencia a organofosforados en estas regiones. Otros estudios en poblaciones de Venezuela detectaron la presencia de otra esterasa denominada A2 con una movilidad relativa de 0,61 (Bisset et al. 2001).
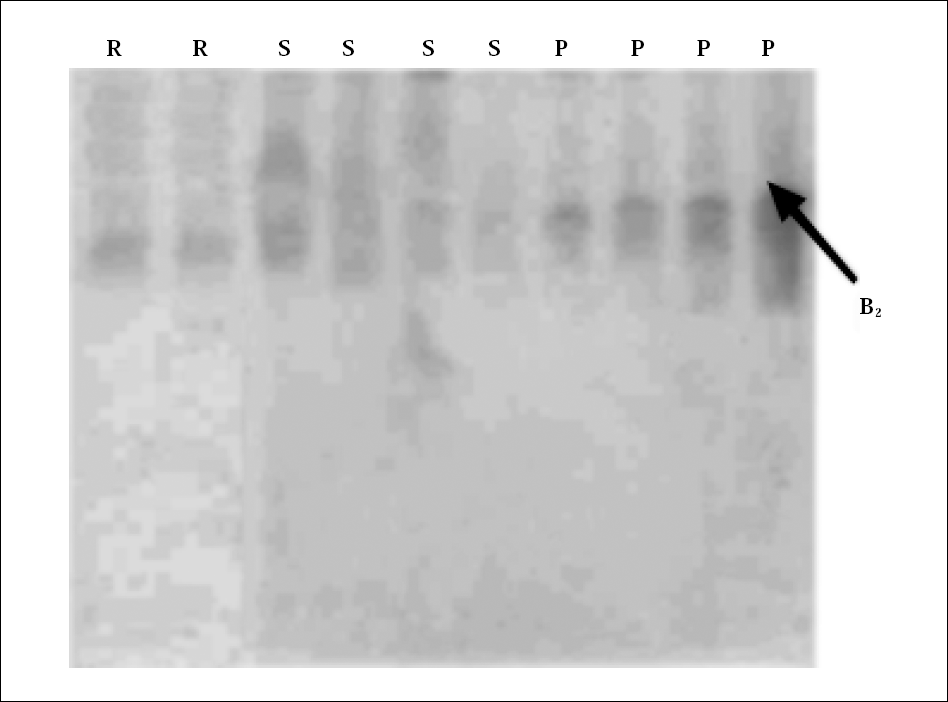
Electroforesis en gel de poliacrilamida (PAGE) mostrando patrones de bandas de esterasa B2 relacionada con la resistencia a organofosforados. R = Rockeffeller, S = Sullana y P = El Porvenir.
Ha sido demostrado la correlación existente entre la actividad de esterasas y la resistencia a insecticidas organofosforados (Bisset et al. 2001). En insectos, de los géneros Culex, Anopheles, Blatella y Pediculus el mecanismo más común de resistencia a insecticidas organofosforados involucra la presencia de una o más esterasas (Amevigbe et al. 2000; Chandre et al. 1999).
La probable intervención de las esterasas en la resistencia cruzada a piretroides ha sido señalada en Culex quinquefasciatus (Say) a través de estudios con sinergistas, detectándose juntos a las esterasas B4 a B6, ya habiéndose registrado previamente también en áfidos (Bisset et al. 1996). Otro mecanismo de resistencia a piretroides es la aparición del gen kdr, y la presencia de oxidasas del citocromo P-450. El primero se debe al producirse un simple cambio de algún aminoácido en el sitio de anclaje del insecticida en el canal de sodio (CICE 2000), y el segundo es un mecanismo de detoxificación mediante oxidaciones e hidroxilaciones (Bisset 2002). Se ha indicado que, en poblaciones de Brasil y Vietnam, la causa de resistencia a piretroides y DDT es la presencia del gen kdr (Brengues et al. 2003), de lo cual se infiere que éste sería el mecanismo probable que estuviera interviniendo en la resistencia a deltametrina en la población Sullana de Ae. aegypti al no detectarse bandas de esterasas B1 y B6.
Los resultados obtenidos en el presente trabajo indican la necesidad de realizar monitoreos para determinar los niveles de resistencia en poblaciones naturales, pues Ae. aegypti podría convertirse en una seria amenaza para las operaciones de control químico.
Conclusiones
El insecticida temefos puede seguir siendo utilizado en los programas de control del vector Aedes aegypti al mostrar está especie aún susceptibilidad a este organofosforado, caso contrario sucede para deltametrina donde se debe evaluar su efecto en poblaciones naturales ya que algunas deben estar presentando resistencia como es el caso de la población Sullana.
Por otro lado, la aparición de la esterasa B2 en la población El Porvenir representa un problema a futuro, ya que este tipo de enzima es la responsable de generar resistencia a los insecticidas organofosforados, debiéndose tomar las medidas necesarias para el caso.