
Abstract
Given that ants respond to different practices, generated by agriculture, this paper analyses and compares information from two studies conducted in sugar cane fields of the Cauca Valley. In both studies, two lineal transects were situated in which ten equidistant (10 m) stations were placed for a total of 20 stations; each station had one tuna bait and one pitfall trap, that were operated for 83,3 and 441,6 continuous days, respectively. To predict and compare richness in sugar cane fields and their neighbouring sites, different richness estimators were used that work with species abundance; multiple comparisons analyses and cluster analyses were conducted. We captured 97 species distributed in 22 genera and 6 subfamilies. The richest sugar cane fields were those whose management had less impact on the soil and better care of adjacent sites. The sugar cane fields of El Hatico and Roma were more similar probably due to the regional effect that the habitat mosaic of the Reserve had on its surroundings, while El Medio and San Julian, which represent degraded conditions, were placed in another group. This analyses highlights the importance of sugar cane management as the dominant matrix in the Cauca Valley and the diversification of the landscape in order to preserve the biodiversity pf ant communities.
Introducción
Al inicio de la década de los cincuenta, la industria azucarera se ubicaba en la parte central y norte del Valle del Cauca, zona que contaba con buenas condiciones climáticas, del suelo y los recursos hídricos ofreciendo así condiciones favorables para el surgimiento y estructuración de este sector (Guardiola 1995).
Posteriormente, la quema de la caña implementada en la década de los setenta, generó un gran impacto ambiental (Molina et al. 2000) e inconformidad entre los habitantes de las poblaciones vecinas a los cultivos (CENICAÑA 1997). La expansión de este cultivo creó así grandes cambios en las comunidades naturales, tanto florísticas como faunísticas del valle geográfico del río Cauca, dejando solo algunos remanentes de hábitats naturales como parches aislados de bosque y algunos humedales siendo el paisaje dominante extensos monocultivos de caña de azúcar (Naranjo 1992).
Resultado de esta preocupación ambiental y presión social, la industria azucarera ha movido rápidamente sus esfuerzos para implementar la cosecha en verde (Cock y Torres 1998) y en menor grado el manejo orgánico, que ha mostrado ventajas a nivel ambiental (Molina et al. 2000). Se ha encontrado por ejemplo, que las poblaciones de bacterias y hongos son tres veces mayores en la caña con manejo orgánico que en la caña con manejo convencional (Delgadillo et al. 1994) y en el contenido de materia orgánica casi le dobla (Arias 1994). Entre tanto las emisiones de partículas tóxicas como monóxido de carbono, hidrocarburos y dióxido de azufre en caña con manejo convencional le exceden en promedio en un 60% a la caña con manejo orgánico (Universidad de la Salle y CENICAÑA 1991).
Cabe aclarar que en la caña con cosecha en verde se elimina la quema pre y post cosecha, práctica que debe estar implantada en todo el Valle del Cauca en el 2005 como un convenio entre el sector y el Ministerio del Medio Ambiente. En la caña con manejo orgánico se elimina además la aplicación de insumos químicos que se sustituye por el uso de abonos orgánicos u orgánico-minerales, se implementa la labor manual, el uso de carneros para limpiar los callejones y el acomodo de hojarasca entre surcos (E. Murgueitio, com. pers.).
Puesto que en la actualidad se está generando un proceso de reconversión de caña con manejo convencional a caña con cosecha en verde y manejo orgánico, surge la necesidad de valorar estos sistemas de producción desde el punto de vista biológico y ecológico de los macroorganismos presentes en ellos.
Entre los artrópodos, las hormigas son componentes importantes del ecosistema por que cumplen una variedad de funciones ecológicas; además se han considerado útiles en la evaluación de respuestas bióticas frente a diversas practicas generadas por la agricultura como la fertilización, el arado convencional, las fumigaciones y las quemas (Folgarait 1998) y han sido usadas como el mejor grupo indicador de riqueza de organismos en agroecosistemas (Roth et al. 1994). Teniendo en cuenta lo anterior, este trabajo pretende comparar la riqueza de hormigas en cultivos de caña con manejo orgánico y convencional dependiendo del contexto local (Hacienda o Reserva Natural) y regional.
Materiales y Métodos
Este artículo analiza la información de dos estudios, usando la misma metodología y realizado en una zona de vida común (Armbrecht y Ulloa-Chacón 1999; Ramírez y Enríquez 2002). Los predios comprendidos en estos estudios se encuentran en el valle geográfico del río Cauca que cuenta con una extensión de 420.000 ha en la zona definida por Holdridge como bosque seco tropical (bs-T), se da en alturas fluctuantes de hasta 1.000 msnm, temperatura media superior a los 24°C y precipitación media anual entre 1.000-2.000 mm (Espinal 1968). Tales estudios se realizaron en haciendas ubicadas en diferentes localidades. En el municipio de Santander de Quilichao se ubica la hacienda San Julián, en el municipio El Cerrito la hacienda Roma y La Reserva Natural El Hatico y en el municipio de Zarzal la hacienda El Medio (Fig. 1). Cada hacienda involucra un parche de bosque y su caña vecina a excepción de la hacienda Roma que se encuentra totalmente rodeada de otros lotes de caña (Tabla 1).
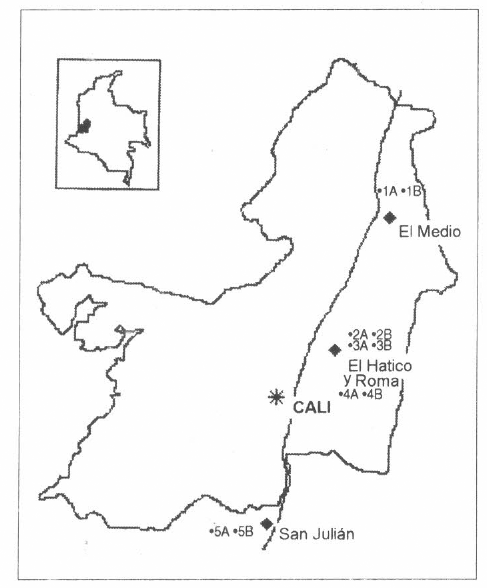
Ubicación de las haciendas donde se encuentran los lotes de caña y sus sitios vecinos. Distancia entre hacienda y hacienda (de sur a norte): San Julián-104 Km – El Hatico – Roma (colinda con El Hatico) 104 Km El Medio. IA bosque El Medio, IB caña vecina El Medio, 2A bosque El Hatico, 2B cañaduzal del bosque El Hatico, 3A guadual El Hatico, 3B cañaduzal del guadual El Hatico, 4A caña Roma 1, 4B caña Roma 2, 5A bosque San Julián, 5B caña vecina San Julián.
Descripción de unidades de cobertura vegetal muestreados en el Valle del Cauca, Colombia. Armbrecht (1995)*, Ramírez y Enríquez (2002)**, común para ambos estudios***
Muestreo de hormigas
En cada sitio se delimitaron dos transectos lineales de 100 m, en cada uno de ellos se ubicaron 10 estaciones equidistantes (10 m) teniendo un total de 20 estaciones por sitio. En cada una se colocó una trampa de caída (pitfall) y un cebo de atún. Las trampas de caída consistieron en un vaso de plástico de 6 cm de diámetro por 6 cm de profundidad, semilleno con alcohol etílico y agua, enterrado a ras del suelo y disimulado con material vegetal. Las trampas se dejaron actuar por un período de 26,5 horas en cada sitio para un total de 441,6 días continuos de muestreo. Los cebos contenían tres a cuatro g de atún sobre una hoja de papel bond de 22 cm x 8 cm y fueron colocados a nivel del suelo por un período de 5 h cada uno, invirtiendo 83,3 días continuos de muestreo total. Las hormigas capturadas se introdujeron en frascos con alcohol al 70%, que fueron rotulados con fecha y lugar de colección.
Se identificaron las morfoespecies colectadas hasta el nivel de género con la ayuda de las claves taxonómicas de Jaffé et al. (1993), Hölldobler y Wilson (1990) y Bolton (1994) y con comparaciones hechas con las colecciones de referencia de la Universidad del Valle. Simultáneamente se elaboraron colecciones de referencia que sirvieron de base de comparación de todo el estudio. Posteriormente, con el fin de establecer las morfoespecies en común para ambos estudios, se hizo una comparación con las colecciones de referencia.
Análisis de los datos
Para realizar los análisis estadísticos de las hormigas, se tomó como unidad de muestreo la trampa de captura. Se calcularon varios estimadores de riqueza que trabajan con la abundancia de especies (ACE, Chao 1 y Michaelis Menten Mean), los cuales permitieron predecir y comparar la riqueza de hormigas en la caña y su respectivos sitios vecinos. Este análisis se realizó mediante el programa Estimates 5 (Colwell 1997).
La densidad de especies de hormigas por unidad muestral fueron comparadas mediante una prueba de Kruskall – Wallis, además la densidad promedio de hormigas (número de especies por estación) entre los lotes de caña fue confrontada mediante una comparación múltiple por rangos de medias. Posteriormente, entre la densidad y la riqueza de hormigas de los lotes de caña, se hicieron análisis de correlación y regresión con el programa Statix para Windows (Statistix 1998). Para visualizar diferencias en la composición de las comunidades de hormigas se utilizó análisis de aglomeración (cluster) con el programa STATISTICA (Statsoft 1995). Para validar el presente trabajo, se usaron los datos de ambos estudios en el bosque El Hatico. Mediante una tabla de contingencia de 2 x 3 (Zar 1996) se examinó la independencia de los dos muestreos (es decir, el de Armbrecht 1995 y el de Ramírez y Enríquez 2002) y la proporción de especies exclusivas y compartidas en los dos períodos de muestreo implicados en ambos estudios. Si la prueba de Chi cuadrado no fuera significativa, se asumiría que los estudios abarcaron especies de hormigas comparables.
Resultados
Como primer paso y para determinar si los períodos de trabajo de campo de los dos estudios reflejaron de una manera confiable la mirmecofauna presente en los sitios incluidos en este artículo, se realizó una comparación de la composición de especies (teniendo en cuenta las compartidas y exclusivas) en los dos períodos de muestreo del bosque El Hatico. Se encontró que no hubo una diferencia significativa (Xβ = 4,56; g.l = 2; 0,1 < p < 0,25) lo que indica que las comunidades de hormigas del bosque para estos dos momentos de muestreo pertenecen a la misma población, sugiriendo que se pueden utilizar, con confianza, los datos de ambos estudios.
Se capturaron 97 especies distribuidas en 22 géneros de 6 subfamilias, de las cuales 60 estuvieron presentes en la caña de azúcar y 64 en los sitios vecinos. Se encontró que los cañaduzales con mayor número de especies de hormigas fueron los de El Hatico, seguidos de la Hacienda La Roma y en último lugar El Medio y San Julián (Tabla 2). De acuerdo con los estimadores de riqueza que permiten estimar el porcentaje de especies que hay en los cañaduzales (Tabla 3), el índice de ACE es un estimador de cobertura basado en la abundancia, indica que se muestreó entre un 41 y 88% de las especies (faltaron por conocer entre 3 y 17 especies), mientras que el índice de Jacknife 1, que se basa en la incidencia y las especies únicas (Colwell 1997), indica que se muestreó entre un 63 y 98% (faltaron por conocer 25 especies).
Especies y número de capturas registradas en cada uno de los cañaduzales y sitios vecinos en el Valle del Cauca, Colombia. (*) Especies registradas por Armbrecht (1996)
Valores observados y estimados de la riqueza de hormigas ordenados de manera decreciente en diferentes caña de azúcar y sus sitios vecinos (S. obs = riqueza observada)
Estos dos índices se ven más afectados por la diferencia entre los llamados “singletons” y “doubletons” con base en los cuales trabajan los estimadores no paramétricos (Mc Kamey 1999). Los “singletons” son aquellas especies que tienen un solo individuo capturado dentro de un hábitat y se convierte en “doubletons” cuando se capturan dos individuos. Cuando el valor de los “singletons” se acerca al de los “duobletons” significa que se está llegando a la asíntota en la curva de saturación de especies (Colwell y Coddington 1994). Por otra parte, el índice de MMMean, que trabaja con la media de especies en la curva de acumulación, es más estable frente a la variación de estos dos valores y sugiere que se ha muestreado entre un 65 y 88% (faltaron por conocer en promedio tres especies).
Al observar los valores de la caña se ve que las de menor riqueza presentan mayor fluctuación en el porcentaje de especies conocidas, esto puede ser por la diferencia que hay entre los “singletons” y los “doubletons” como en el caso de la caña El Medio, o por la ausencia de “doubletons” como en la caña San Julián. En la curva de saturación de todos los sitios los cañaduzales de la Reserva El Hatico son las que menos se acercan a la asíntota de saturación, en un nivel intermedio se encuentran la caña Roma y El Medio, donde hay un mejor conocimiento de las especies de estos cañaduzales y muy paralela al eje x se encuentra la de San Julián, lo que indica que se conocen casi todas las especies de éste (Fig. 2A).
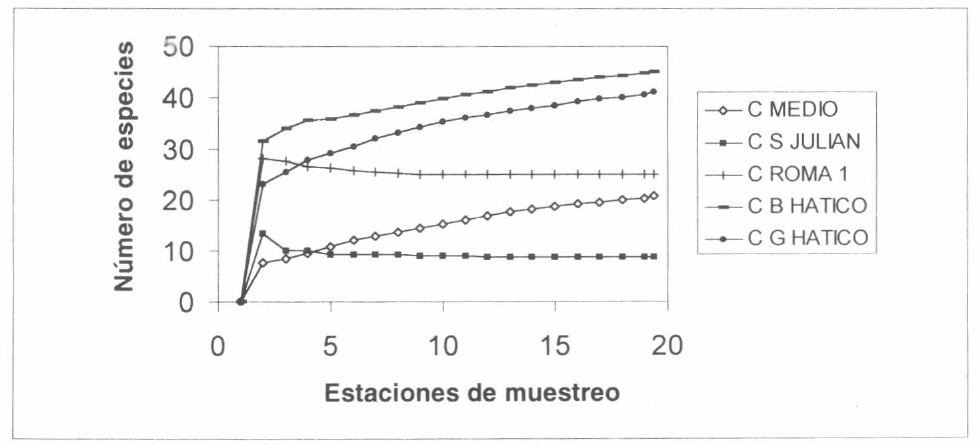
Curva de saturación realizada con el estimador MMMean para los cañaduzales muestreados en el valle geográfico del río Cauca. C = cañaduzal, C B Hatico = Cañadulzal Bosque Hatico, C G Hatico = Cañaduzal Quadual Hatico.
Una tendencia similar muestran los sitios vecinos, ya que la curva del bosque El Hatico indica que aún faltan especies por conocer; más estables se encuentran el guadual El Hatico, la caña Roma 2 y el bosque El Medio. El bosque San Julián al igual que el cañaduzal correspondiente muestra una rápida tendencia a la saturación de especies (Fig. 2B).
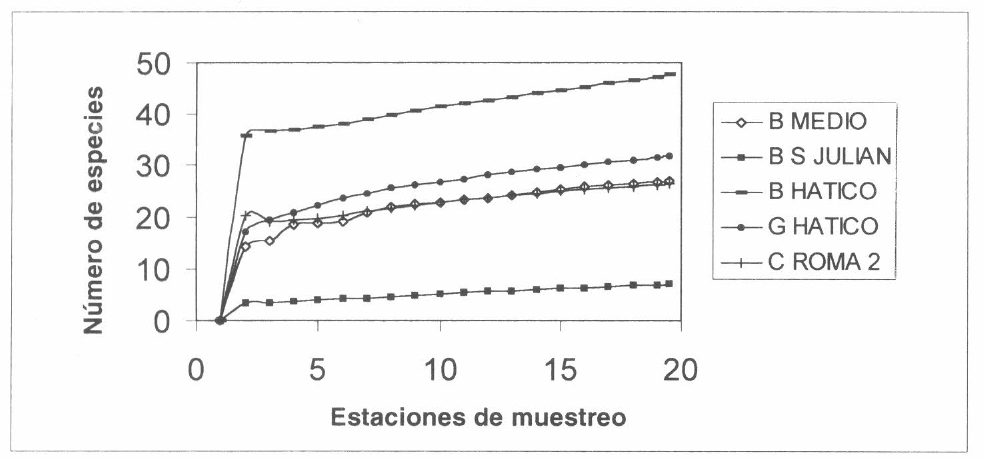
Curva de saturación con el estimador MMMean de los sitios vecinos a cada cañaduzal de la figura 2A, valle geográfico del río Cauca. B= bosque, G= guadual. C= cañaduzal.
Una prueba de Kruskal Wallis encontró diferencia significativa entre los cañaduzales y sus sitios vecinos (H= 9; N= 200; p < 0,0001). Estas diferencias se evidencian al confrontar la abundancia relativa de todos los cañaduzales en un análisis de aglomeración (Fig. 3). Se observa que la caña Roma 1, que se encuentra inmersa en una matriz de caña, está en un grupo aparte pero cercanamente relacionada con la caña San Julián y El Medio que también presentan un manejo convencional. Los cañaduzales que se encuentran más separados de todo el grupo son los de la Reserva Natural El Hatico; es decir, los que presentan un contexto y manejo diferente (orgánico).
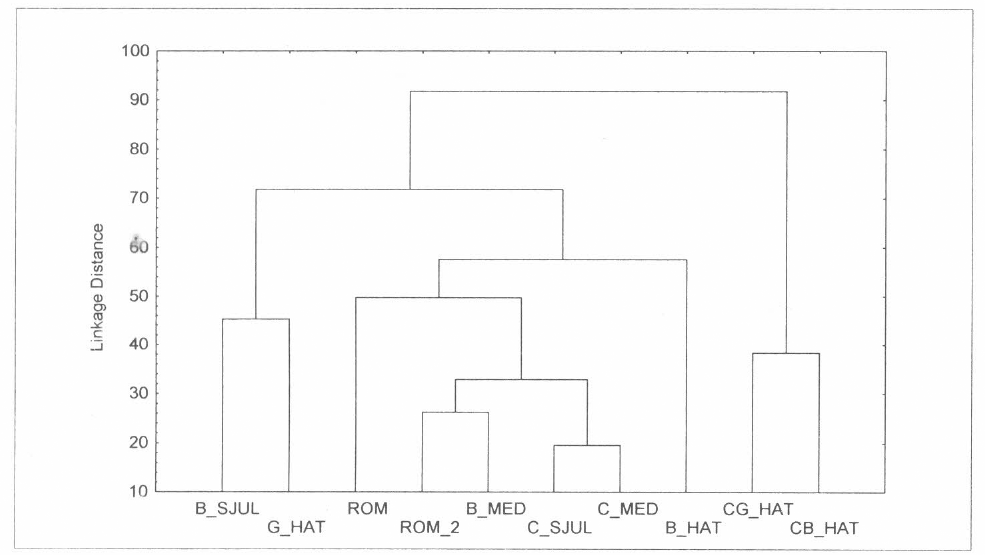
Agrupamiento de los cañaduzales y sitios vecinos con base en la abundancia relativa de la mirmecofauna. Distancias euclidianas (UPGMA).
De acuerdo con la composición de la mirmecofauna, es decir utilizando para los análisis una matriz de datos que solo da cuenta de las identidades en los diferentes cañaduzales y sitios vecinos, muestran un agrupamiento diferente, pues todos los cañaduzales inmersos y vecinos del área El Hatico se encuentran en un grupo y los cañaduzales El Medio y San Julián en otro grupo (Fig. 4). La tendencia se conserva al hacer la comparación múltiple por rangos de la densidad promedio de hormigas entre los cañaduzales (Tabla 4).
Comparación múltiple por rangos de medias en cinco cañaduzales, Valle del Cauca, Colombia
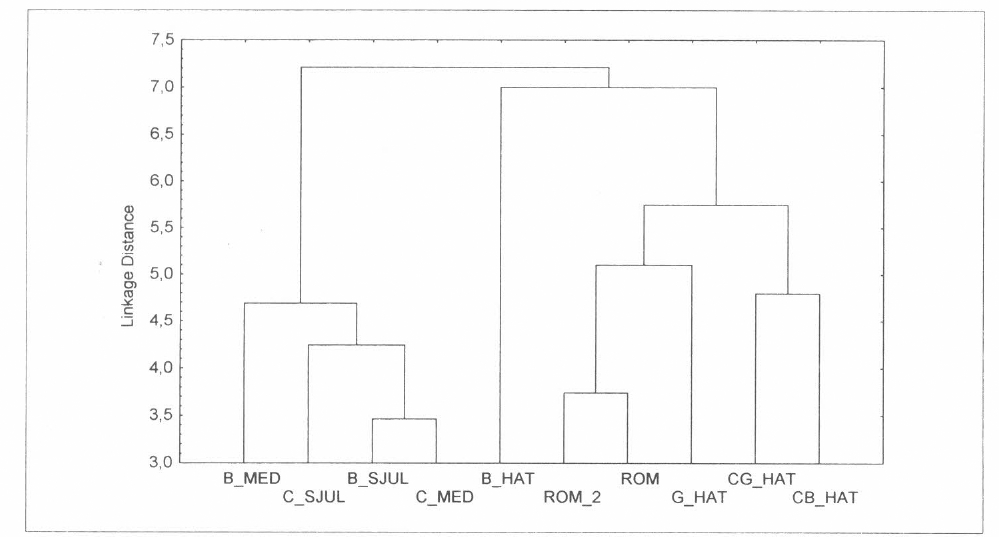
Agrupamiento de los cañaduzales y sitios vecinos con base en la composición de la mirmecofauna. Distancias euclidianas (UPGMA).
Se observa que hay cuatro grupos en que las medias no son significativamente diferentes; éstos coinciden con el tipo de manejo o influencia que están recibiendo. Por un parte está el manejo orgánico en El Hatico, le sigue Roma que aunque es convencional es cercana a una zona de transición de caña convencional a orgánica y a la reserva El Hatico, por último el manejo convencional en El Medio y San Julián. La comparación de la densidad promedio de los cañaduzales es demostrativa si se tiene en cuenta que la densidad y la riqueza de especies se encuentran estrechamente relacionadas (p = 0,95) y además el número de especies por estación explica la riqueza total de cada lote de caña (R2 = 0,9169; t = 6,66; p = 0,0026).
Discusión
Las especies que fueron capturadas en los cañaduzales presentan una tendencia en su composición. Algunos géneros como Wasmannia, Solenopsis, Pheidole, Paratrechina y Ectatomma, tienden a predominar y han sido encontrados con frecuencia en cañaduzales del Valle del Cauca en otros estudios (Gutiérrez et al. 1996; Lastra y Gómez 1996). Es importante anotar que en ninguno de éstos se contó con la presencia de la hormiga loca Paratrechina fulva (Mayr) (Hymenoptera: Formicinae), especie que ha causado impacto ambiental, social y económico en diferentes hábitats (Aldana et al. 1995) y principalmente en cultivos de caña de azúcar en el Valle del Cauca (Gómez y Lastra 1997).
Teniendo en cuenta los valores observados y estimados de la riqueza de Formicidae que se encuentra en los cañaduzales, los de la región El Hatico son los más ricos, en un nivel intermedio se hallan los cañaduzales El Medio y Roma y con una pobreza notable se ve San Julián, esto puede deberse a la combinación de varios factores.
En primer lugar el manejo que recibe cada uno de los cañaduzales puede estar influenciando su riqueza, prácticas como la quema, la aplicación de insumos químicos y la remoción de la hojarasca del suelo pueden tener un impacto negativo sobre las comunidades de hormigas, ya que eliminan todo posible refugio y fuente disponible de alimento y se ha demostrado que a mayor complejidad del hábitat y mayor diversidad de las comunidades de hormigas es menor la incidencia de la hormiga loca (Vargas 2002).
También puede salir afectado el bosque como en el caso de la hacienda El Medio cuando se quema la caña, pues en algunas ocasiones el fuego alcanza los bordes de éste (obs. pers. Inge Armbrecht 1995). Este tipo de manejo crea un gran impacto sobre el suelo pues primero se hace una quema para dejar el tallo de la caña limpio para cortar y posteriormente se hace una requema con el fin de eliminar la hojarasca para facilitar las labores de preparación del terreno (Molina et al. 2000).
Una situación contraria es la que se da en El Hatico en donde se han establecido una serie de prácticas que son más amigables con el medio ambiente, como la eliminación de quemas e insumos químicos y la disposición entre surcos de la hojarasca que queda como residuo de la cosecha. Dichas labores, pueden crear condiciones para que se formen microhábitats donde las hormigas puedan anidar y buscar presas disponibles para su alimentación. Por ejemplo, el gremio de las generalistas en la comunidad de hormigas forrajeras muestra cambios frente a alteraciones de las condiciones físicas de la sombra, teniendo la hojarasca un efecto de considerable importancia (Perfecto y Vandermeer 1996).
Por otro lado se ha notado que en cañaduzales que tenían una mayor complejidad espacial (debido a la presencia de malezas) y aumento de sustratos aprovechables para anidar y forrajear, había una mayor diversidad de hormigas y otros organismos depredadores (Hernández et al. 2002) y por el contrario se ha encontrado una relación inversa entre la complejidad vegetal y diversidad con la presencia de P. fulva (Vargas 2002).
En este sentido esta misma diversidad de Formicidae podría ser utilizada en beneficio propio de los cultivos de caña, ya que existen hormigas de alta eficiencia depredadora. Por ejemplo, la acción depredadora sobre huevos y larvas de barrenadores en el caso de Solenopsis geminata (Fabricius) (Hymenoptera: Myrmicinae) ha sido observada por Velez (1985) y Long et al. (1987); además, esta especie ha sido importante consumidora de semillas en agroecosistemas (Saks y Carroll 1980; Torres 1984) y ha sido propuesta para el manejo de malezas en este cultivo en particular (Reagan 1986). La acción de esta especie podría ser un punto de especial interés y en el que valdría la pena profundizar en estudios futuros ya que se ha encontrado una relación significativa de asociación negativa entre P. fulva y S. geminata (Vargas 2002).
El agrupamiento por abundancia relativa muestra que los cañaduzales tienen una tendencia a agruparse de acuerdo con el manejo que se les da. En el centro del cluster están todos los cañaduzales que presentan un manejo convencional y en transición y en otro grupo están las que presentan un manejo orgánico; esto podría significar que la mayor presencia de ciertas especies o dominancia como la de Wasmannia auropunctata, S. geminata y Solenopsis sp. 1 podría estar influenciando el parentesco entre los cañaduzales.
Comparando los sitios con la composición de especies se encontró que la forma de agruparse es diferente ya que los cañaduzales de la reserva y de la hacienda Roma están en un grupo y los cañaduzales de las haciendas El Medio y San Julián se encuentran en otro grupo; esto puede deberse al efecto local que ejerce cada uno de los sitios. En este caso la riqueza del bosque y el guadual Hatico es mayor a la de los bosque San Julián y El Medio, lo que muestra la dinámica; es decir, el intercambio biótico que existe entre la matriz y los parches de bosque (Armbrecht 1996; Ramírez y Enríquez 2002). Pero por otro lado la caña podría también estar jugando el papel de fuente para unas especies y sumidero para otras, haciendo posible el aprovechamiento de los recursos de uno u otro lado por parte de las especies de hormigas habitantes. Y una matriz con hábitats muy degradados puede estar poniendo en peligro de extinción las especies del bosque (Perfecto y Vandermeer 2001), por que a pesar de que dos biotopos adyacentes sean muy diferentes entre sí, éstos no se encuentran completamente aislados (Doak y Mills 1994).
Al observar la comparación por rangos de medias entre los cañaduzales se puede pensar que hay un efecto regional (Perfecto y Vandermeer 2001), ya que en los cañaduzales de la hacienda Roma se presenta una riqueza superior a las del Medio y San Julián. Esto posiblemente puede deberse a que estos cañaduzales están rodeados de lotes que están en una transición del manejo convencional al manejo orgánico y segundo al mosaico de El Hatico que cuenta una variedad de biotopos además del bosque y la caña como son potreros arbolados, zonas de frutales, áreas en regeneración y corredores biológicos. Esto puede estar creando refugios temporales para anidar, reproducirse y colonizar áreas adyacentes. Contrario a lo que pasa en el área donde se encuentra el bosque y la caña San Julián que en el momento de estudio era una zona con una matriz de potreros muy degradada.
Conclusiones
La riqueza de hormigas de los cañaduzales varía de acuerdo con el contexto local y regional en que se encuentra cada una de ellas ya que el flujo de especies entre los cañaduzales y sus sitios vecinos así como entre localidades pueden estar jugando un papel fundamental en el mantenimiento de la diversidad de hormigas en éstos.
A nivel local, el manejo que recibe cada cañaduzal es importante ya que prácticas como la acumulación de hojarasca, la eliminación de quemas e insumos químicos pueden favorecer la riqueza de las comunidades de hormigas por la disposición de alimento y sitios para anidar.
A nivel regional, la diversificación de un área en particular como el caso de El Hatico es de gran importancia ya que puede crear refugios temporales para las hormigas y actuar como un corredor biológico para colonizar áreas adyacentes. Por otro lado, el degrado de un área pude influir sobre la riqueza de un cañaduzal como es el caso del Medio y San Julián.
Una parte de las especies de la comunidad puede cambiar (posiblemente en forma cíclica) a lo largo del tiempo debido posiblemente a la estacionalidad de un recurso o condición dada, para que éstas se queden en un lugar determinado o emigren hacia otros hábitats. Esto da una idea de que puede existir una dinámica a través del tiempo y el espacio. En este análisis se encontró que las comunidades de hormigas del bosque El Hatico se conservan de una manera más o menos estable en el tiempo, permitiendo así hacer comparaciones de los cañaduzales y sus sitios vecinos en dos épocas diferentes.
Sería útil abordar estudios sobre las comunidades de hormigas, principalmente de aquellas especies que sean conspicuas en un cañaduzal determinado y precisar como éstas traslapan sus nichos con otros depredadores con el fin de dilucidar el papel que ellas tiene dentro de un complejo de enemigos naturales, lo cual sería interesante para implementar programas de control biológico en el año 2005 cuando comienza el sistema de producción de cosecha en verde.
Por último, dada la gran importancia paisajística de la caña de azúcar como monocultivo dominante en el valle geográfico del río Cauca y con base en los datos de este estudio se recomienda la diversificación del paisaje y protección de la cuenca del río Cauca, así como la protección de vegetación nativa como guaduales y bosque tanto para proteger la fauna relictual del bosque seco como para prevenir invasiones de la hormiga loca a cañaduzales no infestados aún.
Footnotes
Agradecimientos
A Patricia Chacón, Philip Silverstone, Enrique Murgueitio y Enrique Molina las autoras agradecen su valiosa colaboración. A los dueños de las haciendas en especial a la familia Molina Durán de El Hatico por permitir la realización de las investigaciones. Los estudios fueron financiados por el Programa de Becas para la conservación, WCS/GEA/FES/ FEN, Fundación para la Promoción de la Investigación y la Tecnología Banco de la República y la Universidad del Valle.