
Abstract
On the Valley of Suárez River (Santander), the association between P. fulva with sucker bug has caused losses up to 3.000 millions pesos in the panelera production. This situation conduced some producers to make wrong handling of crops, raising the negative effects of the ant. The purpose of this research was to establish the P. fulva workers populational fluctuation under the effect of nitrogenade fertilization and low and intensive weed control in panelera cane crops, during 18 th month. Besides, the ant population was tracked in low-disturbed vegetation areas. Finally, the sucker bugs associated with the ant were estimated and the sample methodologies of the P. fulva populations were compared. To estimate the P. fulva populations, pitfall traps with a bait (Frankfurter) were used. The infestation percentage of Saccharicoccus sacchari, Sipha flava, Melanaphis sacchari y Pulvinaria sp. was evaluated according to the location of species in the cane plant. It was determined that nitrogenade fertilization didn't influence in the fluctuation form of P. fulva population. At the same time, a higher abundance of the ant was determined under intensive weed control rather than the low weed control. In relation with sucker bugs evaluated, S. sacchari, established a mutualism relationship with P. fulva, and finally, both sample methods reflected a similar populational fluctuation of P. fulva. These results contributed to the knowledge of environmental and ecological process, which influence the P. fulva population, and can be considered in a management proposal of the bug.
Introducción
La hormiga loca, Paratrechina fulva Mayr, originaria del Sur del Brasil, fue introducida a Colombia hace aproximadamente 30 años como control biológico de las hormigas cortadoras de hoja (Atta y Acromyrmex) y de serpientes. El primer registro oficial de su presencia data de finales de 1971 en el municipio de Puerto Boyacá (Boyacá) (Zenner y Ruiz 1985). En la región de la Hoya del río Suárez (Santander), ubicación del presente trabajo, se halló en agosto de 1989 en el municipio de San Benito, a donde se transportó desde Cimitarra en un cargamento de madera (Insuasty y Manrique 1999).
En esta región, la población de P. fulva ha alcanzado niveles muy altos de infestación, favorecidos por las extensiones amplias de áreas destinadas a la agricultura, especialmente a cultivos de caña de azúcar, en donde parece haber encontrado un ambiente favorable para su multiplicación, y el complemento de su dieta alimenticia ofrecido por insectos chupadores. En consecuencia, la producción panelera, principal base socioeconómica de la región, se redujo en un 60%, con daños anuales que superaron, hasta 1999, los 3.000 millones de pesos, estimándose un total de 5.085 hectáreas afectadas, de las cuales 3.065 (60.18%), correspondían a cultivos de caña de azúcar y las restantes (39.72%) a cultivos de frutales, café, yuca y maíz (Insuasty y Manrique 1999).
Frente a esta situación, algunos agricultores de la zona en su afán de contrarrestar los daños producidos por la asociación entre la hormiga y los insectos chupadores implementaron prácticas de manejo del cultivo fuera de los contextos técnicos establecidos para la caña panelera. Contrario a lo esperado, el problema empeoró intensificándose las pérdidas económicas e incrementando la densidad poblacional de la hormiga.
Con base en estos antecedentes, este trabajo se realizó con el propósito de determinar el efecto de dos prácticas agronómicas de amplia difusión en el cultivo de caña, tales como: el control de malezas y la fertilización nitrogenada sobre el comportamiento fluctuacional de la población de obreras de P. fulva, durante el primer ciclo vegetativo de la variedad de caña para panela RD 75-11. A su vez, para establecer si los cambios fluctuacionales de la hormiga se relacionaban de una u otra manera con el ambiente, se realizó el seguimiento poblacional a P. fulva en zonas de vegetación poco disturbada, a las que se les denominó áreas de seguimiento.
Se utilizó como método de muestreo, para estimar la fluctuación poblacional de la hormiga, la trampa de caída (Pitfall trap) y con atrayente, utilizando como cebo una rodaja de salchicha. Por otro lado, se buscó determinar la especie del insecto chupador que establecía una relación mutualista con P. fulva, registrando el porcentaje de infestación de cada una de las especies presentes en el cultivo de caña.
Finalmente, se compararon los dos sistemas de muestreo utilizados para estimar los cambios poblacionales del insecto y como complemento al trabajo se realizó la identificación hasta el nivel de género de otras especies de hormigas encontradas tanto dentro del agroecosistema de caña como en las áreas de seguimiento, en condiciones de poblaciones altas y bajas de la hormiga loca, con el fin de establecer su efecto sobre la mirmecofauna.
Los resultados obtenidos, además de contribuir al conocimiento ecológico de la población de P. fulva ofrecen herramientas para influir sobre los aspectos y procesos ambientales que están determinando su abundancia, con el objetivo de regular las problemáticas poblaciones del insecto, no sin antes ser incluidos y correlacionados dentro de los parámetros contemplados en un Manejo Integrado de Plagas.
Materiales y Métodos
El trabajo de campo se realizó en parcelas semi-comerciales de los municipios de Guepsa y San Benito (Santander) pertenecientes a la región de la Hoya del río Suárez, durante el período de junio/98 a noviembre/99.
Esta región, según la clasificación de Holdridge (1979) se encuentra dentro de la categoría de Bosque Húmedo Premontano (bh-PM), entre un rango altitudinal de los 900 a los 2000 msnm, caracterizada por presentar una temperatura promedio aproximada de 18 a 24°C, y un promedio anual de lluvias de 1000 a 2000 mm.
Experimentos de fertilización nitrogenada y control de malezas
El experimento de fertilización nitrogenada se localizó en el municipio de San Benito, vereda San Lorenzo a 1080 msnm y contó con una extensión de 11.052 m2. Se aplicaron tres dosis de nitrógeno por hectárea: 50 kg/ha dosis mínima permitida por el productor; 100 kg/ha la recomendada por los técnicos de la región y 300 kg/ha, siguiendo los antecedentes de sobre dosificación hechos por algunos productores de la región. Los otros nutrientes como el fósforo y el potasio se mantuvieron constantes en dosis de 100 kg y de 80 kg/ha, respectivamente.
El experimento de control de malezas se ubicó en el municipio de Guepsa, vereda la Teja, a 1480 msnm y contó con una extensión de 5.934 m2. En este experimento se aplicaron dos tratamientos. El primero de ellos, mediante el control intensivo, buscó no favorecer la presencia de malezas dentro del cultivo, aplicando Anikilamina (150 cm3), Karmex (150 g) y Mixel (40 cm3) (por bomba de 20 L). El segundo tratamiento por el contrario, buscó favorecer las malezas utilizando machete y rastrillo para su remoción.
A su vez, se ubicaron áreas de vegetación poco disturbada adyacentes a los experimentos para evaluar el efecto del medio ambiente sobre la población de hormiga y como punto de comparación al cultivo de caña panelera.
El diseño experimental utilizado para los experimentos fue el de Bloques Completos al Azar, con dos y tres repeticiones de dos y tres tratamientos para los experimentos de control de malezas y fertilización nitrogenada, respectivamente. Por otro lado, específicamente en el experimento de control de malezas, se realizó análisis de varianza como herramienta para determinar la diferencia entre los promedios de captura de la hormiga entre el poco e intensivo control. De la misma forma, se utilizó el análisis gráfico para la interpretación de las respuestas más relevantes obtenidas en este estudio, graficando la fluctuación poblacional de la hormiga representada tanto en la trampa de caída como en la de salchicha por evaluación, para lo cual se tomó el promedio por trampa en cada tratamiento de ambos experimentos y para el caso de las áreas de seguimiento se tomó el promedio en cada evaluación.
Método de muestreo
Para estimar la población de P. fulva se utilizaron dos métodos de muestreo. El primero de ellos, la trampa de caída (Pitfall trap), se basó en el mecanismo de intercepción; estaba constituida por dos recipientes, uno de mayor tamaño que actuaba como contenedor de un segundo recipiente que poseía una solución jabonosa (5 g de jabón en polvo/Litro de agua) en donde quedaban atrapadas las hormigas. Su tiempo de exposición en campo fue de 21 horas.
El segundo, la trampa de salchicha, se basó en la metodología descrita por Gómez y López (1995) en la cual se utilizan cajas para exámenes coprológicos con 8 perforaciones laterales y se utiliza como cebo una rodaja de salchicha, con un tiempo de exposición en campo de 2 horas.
Para estimar la cantidad de individuos capturados, en la trampa de caída se tomó el peso húmedo de cada muestra después de contar el número de individuos presentes en una submuestra y para la trampa de salchicha se realizó un conteo manual de los individuos capturados.
Evaluación de insectos chupadores
Con el fin de establecer las especies de insectos chupadores con las que la hormiga podría mantener una relación mutualista dentro del cultivo de caña, se evaluó el porcentaje de infestación de: Saccharicoccus sacchari (Ckll) (Hom: Pseudococcidae), Sipha flava (Forbes) y Melanaphis sacchari (Z) (Hom: Aphididae), y Pulvinaria sp. (Hom: Diaspididae), siguiendo la metodología descrita por Gómez et al. (1990), que consiste en tomar de cada tallo las cuatro primeras hojas a partir del primer cuello visible, contando el número de hojas infestadas por cada especie. En los pulgones se registró al menos una colonia grande o dos colonias de dos o más individuos cada una, y la escama se determinó por su presencia ó ausencia en el envés de la hoja. La infestación de S. sacchari se evaluó registrando su presencia ó ausencia en la vaina de cada tallo de caña evaluado. Finalmente, se graficaron las tendencias de infestación de cada especie de homóptero y se realizaron análisis de correlación entre estas tendencias y la fluctuación poblacional que presentó P. fulva en cada experimento.
Efecto de P. fulva sobre la mirmecofauna
Como complemento al trabajo se identificó hasta el nivel de género la mirmecofauna presente en el cultivo de caña y en las áreas de seguimiento, colectada con el mismo método de muestreo de P. fulva. De esta forma se determinó el efecto de las poblaciones altas y bajas de la hormiga loca sobre otras especies de hormigas. Para el proceso se contó con la asesoría del Dr. F. Fernández, Taxónomo del Instituto Alexander Von Humboldt (Villa de Leiva, Boyacá) y haciendo uso de las claves taxonómicas de Fernández et al. (1996) y Trochez (1998) para el nivel de subfamilia y de Fernández (1990) y Lattke (1993) para el nivel de género.
Resultados y Discusión
Fluctuación poblacional de P. fulva bajo efecto de la fertilización nitrogenada
Durante el período evaluado la fluctuación poblacional de P. fulva se representó similarmente tanto por la trampa de caída como por la de salchicha, estableciéndose mediante el análisis gráfico que la fertilización nitrogenada no influyó en la forma como fluctúa la población de obreras de la hormiga loca.
Según los registros de los dos tipos de trampa, durante los primeros seis meses de edad del cultivo, la captura de obreras de P. fulva, fue baja e inferior a 126 ind/ trampa y a 383 ind/trampa en las trampas de caída (Fig. 1) y salchicha (Fig. 2) respectivamente. En los siguientes seis meses la captura se incrementó notoriamente a 5.180 ind/trampa en el primer caso y a 335 ind/trampa en el segundo. Finalmente, la población disminuyó a 1.877 ind/trampa de caída y a 181 ind/trampa de salchicha.
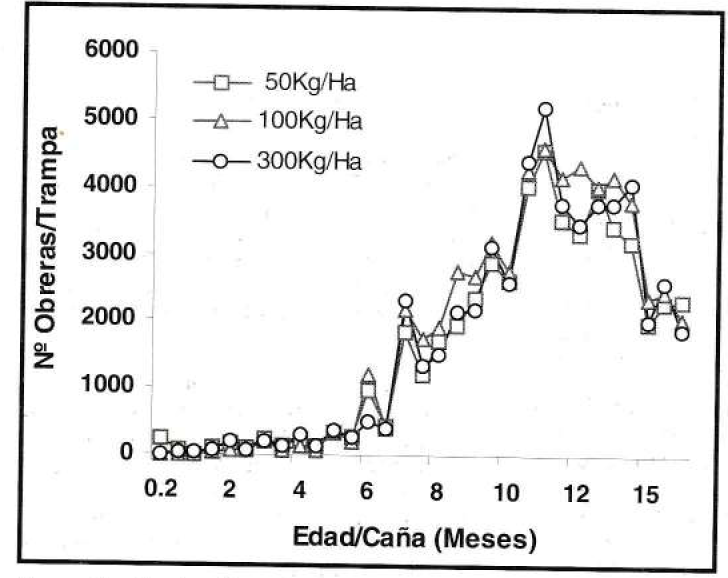
Fluctuación poblacional de P. fulva, con la trampa de caída (prom./trampa/eval) en el experimento de fertilización nitrogenada.
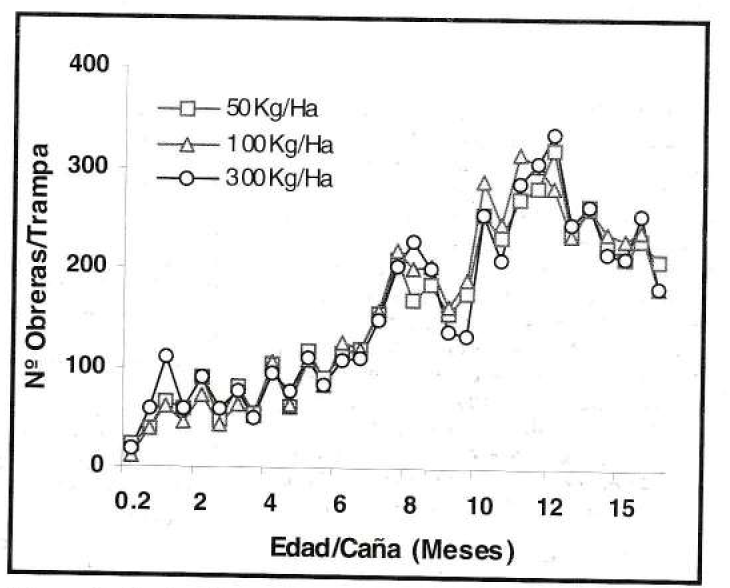
Fluctuación poblacional de P. fulva, con la trampa de salchicha (prom./trampa/eval) en el experimento de fertilización nitrogenada.
Evidentemente, el agroecosistema de caña de azúcar a lo largo de su desarrollo fenológico ofreció diversos nichos ecológicos en los que la hormiga presentó diferentes estados de proliferación. Por tal razón, durante los primeros meses de edad del cultivo, cuando los condiciones aún no eran adecuadas para la proliferación de P. fulva, se presentó el establecimiento del insecto con poblaciones bajas, pero posteriormente se creó un hábitat de condiciones favorables para su proliferación. Sin embargo, el descenso poblacional de los últimos meses, probablemente fue consecuencia de factores dependientes de su densidad, ya que la hormiga al quedar confinada en un ambiente reducido se desplazó a otras áreas de poca proliferación.
En comparación, en el área de vegetación poco disturbada la población de P. fulva fue baja, presentando una fluctuación constante pero irregular con capturas entre 26 y 212 ind/trampa de salchicha y entre 2 y 176 ind/trampa de caída (Fig. 3). Este comportamiento poblacional es atribuible a la diversidad y estabilidad vegetal en el área, con relación a la del cultivo, ya que ciertas condiciones microclimáticas y la posible presencia de enemigos naturales no favorecieron la invasión de la hormiga.
Fluctuación poblacional de P. fulva, en la parcela de seguimiento alterno (prom./trampa/eval).
Fluctuación poblacional de P. fulva bajo efecto del control de malezas
Inicialmente, el área en la que se ubicó este experimento se caracterizó por la abundancia de P. fulva. Sin embargo, durante el período evaluado la densidad poblacional disminuyó notoria e inesperadamente, variando la captura entre nula y hasta 24 ind/trampa de caída (Fig. 4) y 140 ind/trampa de salchicha (Fig. 5).
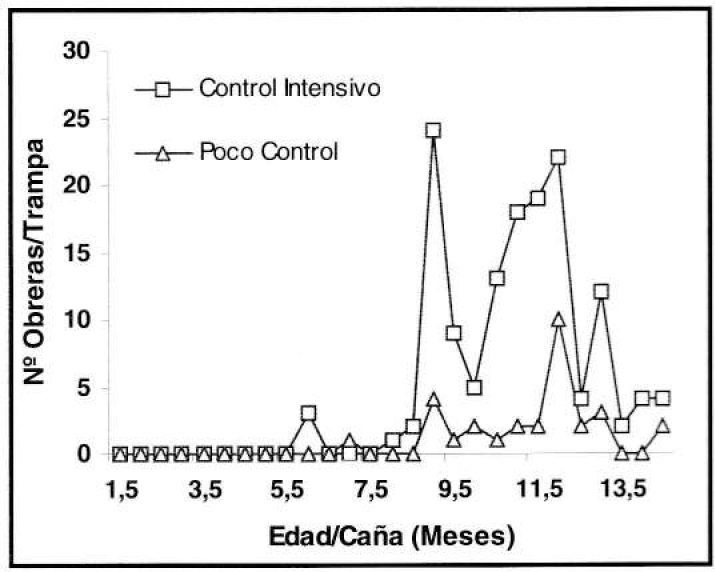
Fluctuación poblacional de P. fulva, con la trampa de caída (prom./trampa/eval) en el experimento de control de malezas.
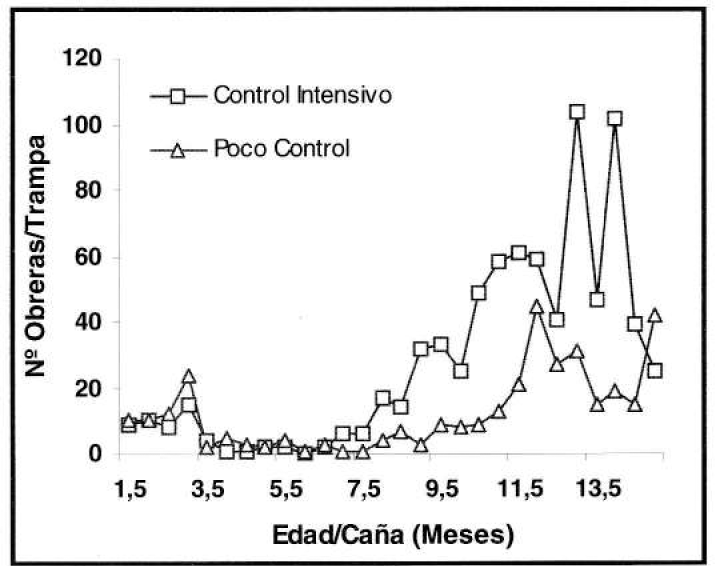
Fluctuación poblacional de P. fulva, con la trampa de salchicha (prom./trampa/eval) en el experimento de control de malezas.
El registro poblacional más bajo se presentó en los primeros meses de edad del cultivo, en comparación con los últimos, pues en éstos se comenzó a detectar con los dos métodos de muestreo la tendencia a capturar un número mayor de hormigas en donde la presencia de malezas fue menor. Según el análisis estadístico se encontraron diferencias significativas a un nivel de confiabilidad del 95% entre el poco y el intensivo control de malezas, tanto para la trampa de caída (F= 27.73) como para la de salchicha (F= 46.07), estableciéndose un mayor promedio de captura en el control intensivo, con relación al poco control posiblemente a causa de la menor competencia entre la caña y las malezas controladas, permitiendo un mayor crecimiento y desarrollo del cultivo, ofreciendo a la hormiga condiciones microclimáticas favorables y número mayor de sitios de anidamiento.
Seguidamente, se registró un descenso poblacional hacia el final del ciclo vegetativo del cultivo, comportamiento fluctuacional muy semejante al del experimento de fertilización, pues la densidad poblacional alcanzó un máximo para luego disminuir.
Nuevamente, en la tendencia fluctuacional de la hormiga influyeron, además del propio desarrollo fenológico de la caña, ciertas condiciones ambientales, creadas por el efecto de la presencia o ausencia de las malezas dentro del cultivo, ya que al presentarse una mayor heterogeneidad vegetal se vio favorecido el establecimiento de otros organismos como depredadores naturales u otras especies de hormigas que incidieron desfavorablemente sobre la abundancia de P. fulva. Confirmándose, que el desarrollo físico y fenológico así como las prácticas asociadas al cultivo de caña de azúcar, favorece el establecimiento y proliferación de la hormiga loca.
Finalmente, en la franja de seguimiento no se capturaron más de 27 ind/trampa de salchicha y de 4 ind/trampa de caída, presentándose una abundancia mayor hacia las últimas evaluaciones. No obstante, el patrón fluctuacional de la población mantuvo una secuencia irregular (Fig. 6).
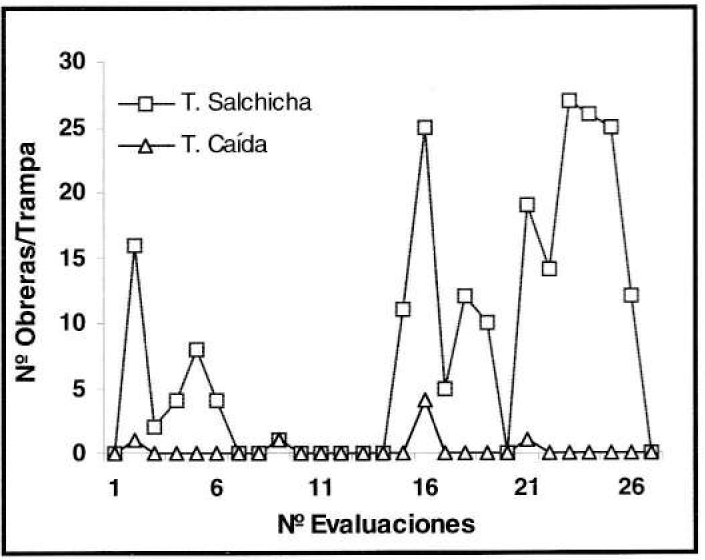
Fluctuación poblacional de P. fulva, en la franja de seguimiento adyacente al experimento de control de malezas (prom./trampa/eval).
Infestación de insectos chupadores
De los homópteros evaluados S. sacchari y S. flava fueron los que presentaron los mayores porcentajes de infestación, en comparación con M. sacchari y Pulvinaria sp., que por su esporádica presencia no se tuvieron en cuenta para los resultados.
La tendencia de infestación establecida por S. sacchari, en el experimento de fertilización nitrogenada, registró que el insecto presentó durante el primer mes de evaluación, infestación en alrededor del 51% de los tallos de caña, siendo el nivel mas bajo de presencia del homóptero. En los dos meses siguientes su presencia aumentó considerablemente alcanzando en el 8º mes de edad del cultivo el 91% de tallos infestados. Posteriormente, en los últimos nueve meses la infestación osciló entre el 84% y 100% (Fig. 7). Por el contrario, S. flava, mostró en los tres primeros meses de evaluación el porcentaje mayor de hojas infestadas con rangos entre el 10% y 50%, disminuyendo su presencia en los restantes seis meses a niveles que no superaron el 14% de infestación (Fig. 8).
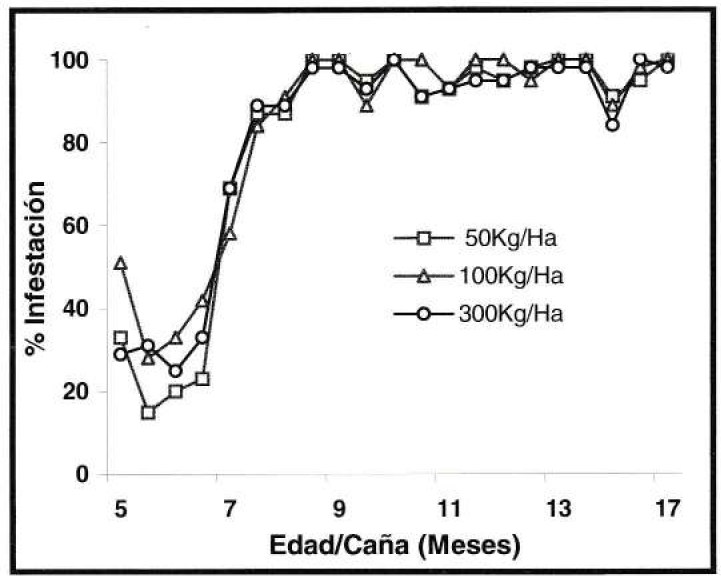
Tendencia de Infestación de S. sacchari, en el experimento de fertilización nitrogenada (prom./eval), durante 12 meses (Valores expresados en porcentajes).
Tendencia de infestación de S. flava, en el experimento de fertilización nitrogenada (prom./eval), durante 9 meses (Valores expresados en porcentajes).
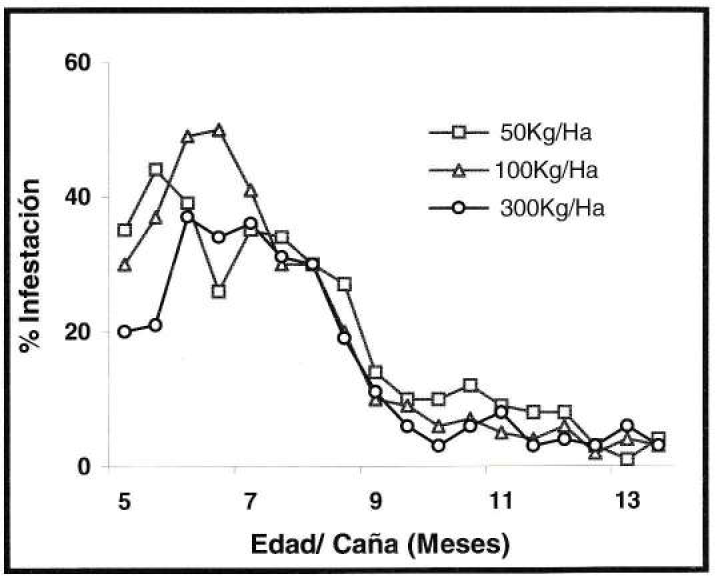
Con relación a la tendencia de infestación de S. sacchari, en el experimento de control de malezas, durante el primer mes de evaluación fluctúo entre nula y el 7%, registrándose un aumento al 89% en los 3.5 meses siguientes, para luego disminuir hasta el 50% en los últimos meses (Fig. 9). Por otra parte, S. flava, durante el primer mes de evaluación presentó como máximo el 14% de hojas infestadas, presentándose en los cinco meses siguientes los porcentajes más bajos que no superaron el 4% de infestación. En los meses restantes la infestación aumentó levemente (Fig. 10).
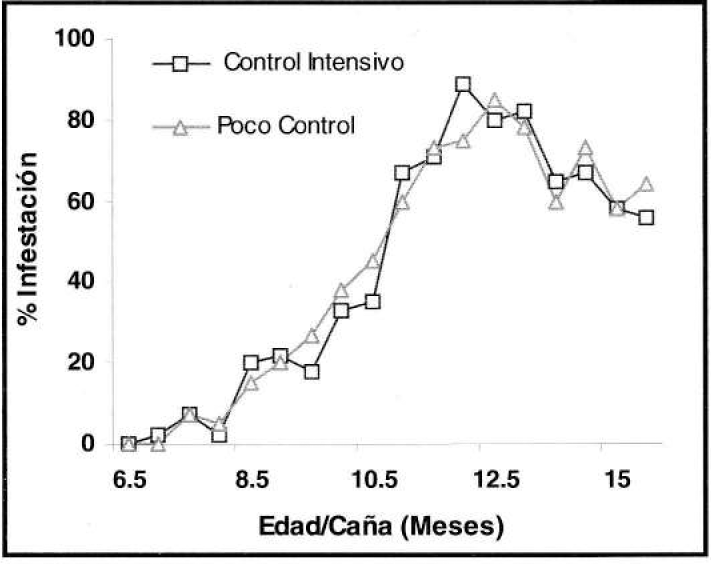
Tendencia de infestación de S. sacchari, en el experimento de control de malezas (prom./eval), durante 10 meses (Valores expresados en porcentajes).
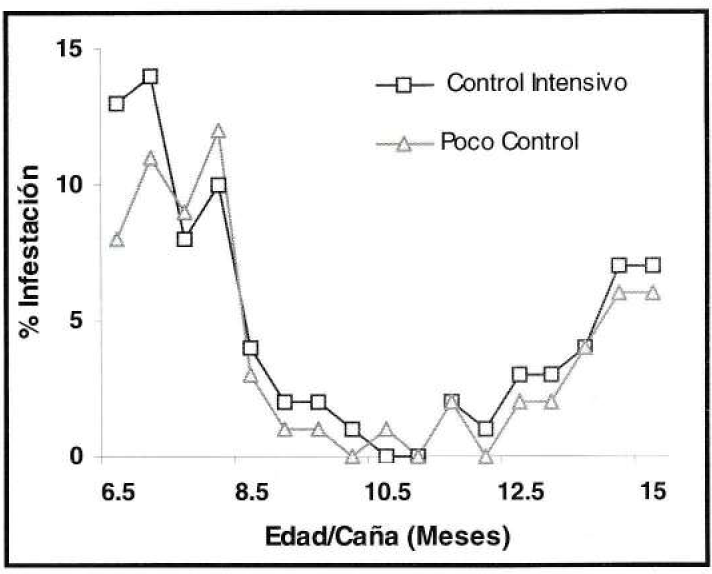
Tendencia de infestación de S. flava, en el experimento de control de malezas (prom./eval), durante 8.5 meses (Valores expresados en porcentajes).
De los anteriores resultados y considerando la tendencia de las gráficas, se pudo determinar que el comportamiento de infestación que presentaron S. sacchari y S. flava en el cultivo de caña de azúcar fue semejante entre las tres dosis de nitrógeno aplicadas, así como en el poco e intensivo control de malezas, por lo tanto no se evidenciaron diferencias entre los tratamientos de cada experimento. Por consiguiente, la necesidad alimenticia de la cochinilla fue independiente de los efectos fisiológicos que pudieron haber causado en las plantas las diferentes dosis de nitrógeno en el experimento de fertilización y el poco e intensivo control de malezas, lo que indica que la tendencia de infestación de S. sacchari estuvo asociada a los cambios poblacionales de P. fulva.
Del mismo modo, la tendencia de infestación de S. flava sugiere, tal como lo describen Londoño y Gómez (1990), que alrededor de un valor máximo de la presencia del insecto, hay factores internos de su población o bien el estado fisiológico en el que se encontraban las plantas de caña, que inducen la subsiguiente migración o desaparición de los áfidos.
Relación de P. fulva con insectos chupadores
Para el caso de la relación de la hormiga con S. sacchari, en el experimento de fertilización nitrogenada, se determinó un coeficiente de correlación del 82% para la trampa de caída (ρ=0.82) (Fig. 11) y del 77% para la de salchicha (ρ=0.77). Por el contrario, entre P. fulva y S. flava se presentó una correlación negativa del 87% para la trampa de caída (ρ= -0.87) (Fig. 11) y del 76% para la de salchicha (ρ= -0.76).
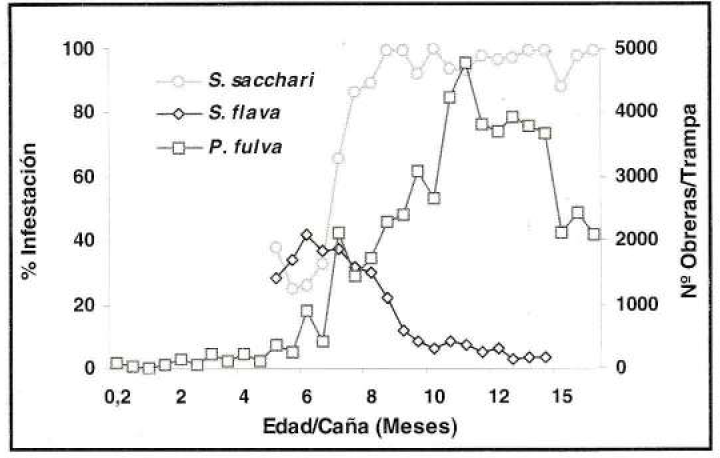
Correlación entre la fluctuación poblacional de P. fulva con la trampa de caída (prom./trampa/eval) y el porcentaje de infestación de S. sacchari y S. flava (prom./eval) en el experimento de fertilización nitrogenada.
Así mismo, en el experimento de control de malezas entre P. fulva y S. sacchari se determinó un coeficiente de correlación del 88% para la trampa de salchicha (ρ=0.88) y del 48% para la de caída (ρ=0.48) (Fig. 12), con referencia a este último el coeficiente bajo probablemente se debió a la captura aleatoria de la trampa y a las bajas poblaciones de la hormiga. A diferencia, entre S. flava y P. fulva, se presentaron coeficientes negativos y menores del 70%, registrándose para la trampa de salchicha el 42% (ρ=0.42) y para la de caída el 69% (ρ= -0.69) (Fig. 12).
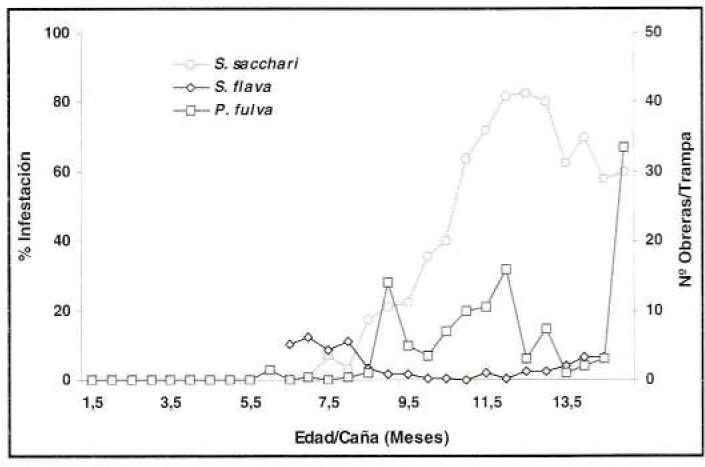
Correlación entre la fluctuación poblacional de P. fulva, con la trampa de caída (prom./trampa/eval) y los porcentajes de infestación de S. sacchari y S. flava (prom./eval), en el experimento de control de malezas.
En conclusión, las poblaciones de S. sacchari y P. fulva presentaron un comportamiento fluctuacional semejante en los dos experimentos, además de correlacionarse positivamente, lo que demostró una relación mutualista entre las dos especies, en la que la hormiga transporta, disemina y protege de depredadores a la cochinilla y ésta le proporciona a la hormiga el suplemento líquido de su alimentación. Además, tal como fue demostrado por Zenner y Ruiz (1985), las poblaciones de los chupadores son considerablemente más altas en presencia de P. fulva.
Con respecto a la relación entre el pulgón y la hormiga, mediante el análisis de correlación, se determinó que la tendencia de infestación del chupador estuvo más en función de la dinámica de su especie que de una relación mutualista con la hormiga, ya que a medida que aumentó la población de P. fulva, la presencia dei pulgón disminuyó, además, el insecto por no producir secreciones azucaradas probablemente no fue de interés para la hormiga.
Comparación de los métodos de muestreo
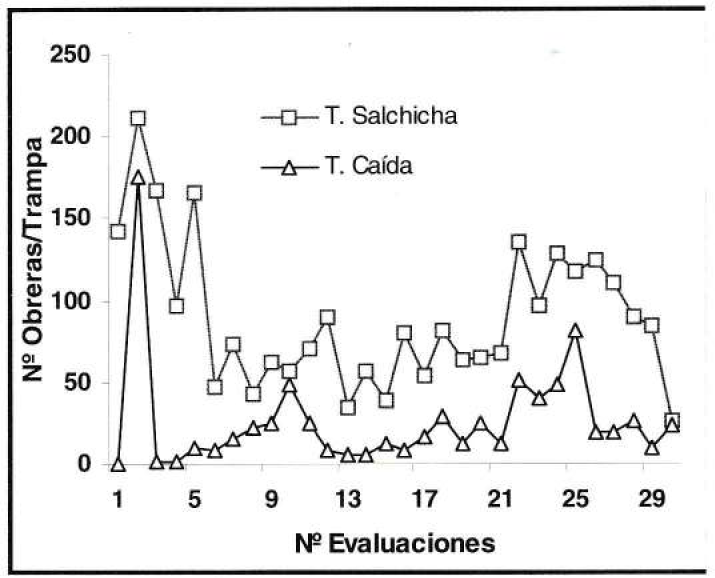
En presencia de poblaciones altas de la hormiga loca, como las estimadas en el experimento de fertilización nitrogenada, la trampa de caída registró el número mayor de obreras capturadas, con un máximo de 5.000 ind/trampa, mientras que la captura máxima en la trampa de salchicha fue de 212 ind/trampa (Fig. 13). Contrario a lo anterior, en condiciones de poblaciones bajas de la hormiga, presentes en el experimento de control de malezas, en las trampas de salchicha se capturó el número mayor de individuos por unidad con un máximo de 67 obreras, mientras que en la trampa de caída no se capturaron más de 14 ind/trampa (Fig. 14).
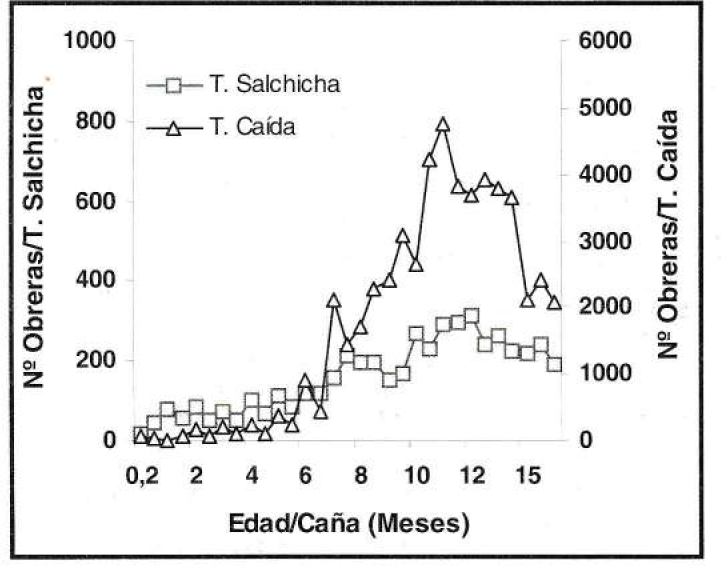
Fluctuación de poblaciones altas de P. fulva.
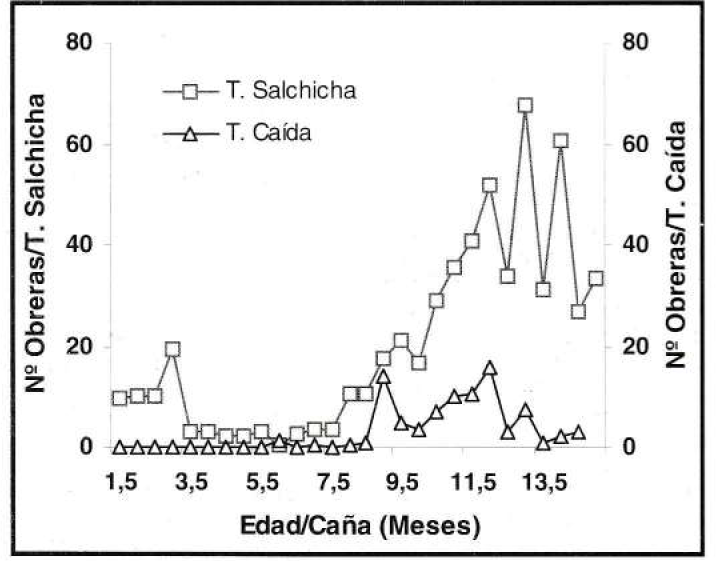
Fluctuación de poblaciones bajas de P. fulva.
Lo anterior, determinó que ambos sistemas de muestreo reflejaron similarmente los cambios poblacionales del insecto, independientemente de la cantidad de individuos capturados. En condiciones de población alta de la hormiga la captura mayor se dio cuando se empleó la trampa de caída que, según Gómez (1998), se caracteriza por colectar individuos que caen aleatoriamente dentro del recipiente, pero por el contrario, en poblaciones bajas la captura mayor fue con la trampa de salchicha, lo que se explica por la concentración mayor de obreras resultante del efecto atrayente que tiene la salchicha, lo que concuerda con lo señalado por Gómez y López (1995).
En relación con la cantidad de esfuerzo requerido para obtener la información fue claro que la trampa de caída demanda un mayor tiempo de exposición, al igual que un mayor tiempo para contar los individuos capturados. Por otro lado, al usar la trampa de caída se detectaron incrementos bruscos y puntuales en la captura, los cuales pueden deberse más a cambios momentáneos del comportamiento de las obreras (aún sin explicación) que a tendencias generales de la población. Por las razones anteriores se considera más conveniente emplear la trampa de salchicha como método de muestreo de las poblaciones de la hormiga loca.
Efecto de P. fulva sobre la mirmecofauna
En el experimento de fertilización nitrogenada las poblaciones altas de la hormiga loca causaron un efecto negativo sobre la presencia de otro tipo de hormigas, puesto que el único género colectado esporádicamente durante el total de las evaluaciones fue Solenopsis sp. (Subgénero Fugaz) en proporciones de 1:10.000 respecto a P. fulva (Tabla 1). En relación con lo anterior, Zenner (1992) sugiere algo similar en el municipio de Cimitarra (Santander), en donde por el efecto de las poblaciones de P. fulva, la fauna local de formicidae disminuyó en un 98% su riqueza original de especies.
Porcentaje de las subfamilias y géneros de mirmecofauna identificados dentro de los cultivos de caña panelera y en áreas de vegetación poco disturbada
A = Experimento de fertilización nitrogenada.
B = Parcela de seguimiento adyacente al experimento de fertilización nitrogenada.
C = Control intensivo de malezas.
D = Poco control de malezas.
E = Franja de seguimiento adyacente al experimento de control de malezas.
% = Porcentaje de las subfamilias.
G = Géneros de mirmecofauna presentes en cada sitio de muestreo.
Por el contrario, en condiciones de poblaciones bajas de la hormiga loca, como las determinadas en el experimento de control de malezas, se identificaron 20 géneros de otras especies de hormigas en el control intensivo y 19 en el poco control, agrupados en 6 subfamilias en donde Myrmicinae ocupa el primer lugar, seguidas de Ponerinae, Formicinae, Ecitoninae, Dolichoderinae y Pseudomyrmex (Tabla 1). De los géneros caracterizados, Zenner y Ruiz (1985) señalan a seis (Solenopsis, Atta, Cephalotes, Camponotus, Pseudomyrmex y Azteca) como desplazados por P. fulva, por lo tanto su presencia dentro del cultivo sugiere la poca agresividad que ejercen las poblaciones bajas de la hormiga loca.
Por otro lado, en las áreas de seguimiento se identificaron 14 y 17 géneros para la parcela y la franja de seguimiento adyacente al experimento de fertilización nitrogenada y control de malezas respectivamente. Para el primer caso, los géneros se agruparon en cuatro subfamilias correspondientes a Myrmicinae, Formicinae, Ecitoninae y Ponerinae, y para el segundo caso los géneros se agruparon en las mismas seis subfamilias presentes dentro del experimento de control de malezas.
En relación con la mirmecofauna presente en estas áreas de seguimiento, la diversidad de hormigas es resultado además de las poblaciones bajas de P. fulva, de la complejidad espacial dentro de las áreas (dada por su estructura vegetal) y del aumento de substratos aprovechables por las hormigas para anidar o forrajear, ya que de acuerdo con Bustos y Chacón (1997) la complejidad espacial permite a ciertas especies ocupar nichos muy específicos y eludir la competencia.
Conclusiones
La población de obreras de P. fulva, fluctuó de acuerdo con su propia dinámica y habilidad de adaptación a las condiciones y fenología propia del agroecosistema y no en función de las dosis de nitrógeno aplicadas al cultivo.
El control intensivo de malezas, a diferencia del poco control, mostró la tendencia de presentar una abundancia mayor de P. fulva.
La población de P. fulva en áreas de vegetación poco disturbada mostró un comportamiento fluctuacional variable pero con tendencia a ser constante, con relación al presentado dentro del cultivo de caña de azúcar.
Las labores del cultivo y el desarrollo fenológico de la caña de azúcar favorecen el establecimiento y proliferación de la hormiga loca.
Se determinó que P. fulva estableció un relación mutualista con S. sacchari, en comparación con otros insectos chupadores evaluados.
Los dos métodos de muestreo reflejaron en forma semejante los cambios poblacionales de P. fulva, independientemente de la cantidad de individuos capturados.
Se estableció que las poblaciones altas de P. fulva ejercieron un efecto negativo sobre la diversidad de hormigas en el experimento de fertilización nitrogenada, en donde únicamente se colectó una especie del género Solenopsis.
Footnotes
Agradecimientos
Esta investigación hace parte del programa Nacional sobre Hormiga loca, financiado por COLCIENCIAS, CENICAÑA, CORPOICA CIMPA y la Universidad del Valle. Agradecemos el apoyo logístico y de asesoría ofrecidos por el Departamento de Biología de la Universidad Pedagógica y Tecnológica de Colombia; al Dr. Fernando Fernández, Taxónomo del Instituto Alexander Von Humboldt por la ayuda y orientación en la identificación de las especies colectadas.