
Abstract
The chaetotaxy of fourth instar larva of Lutzomyia (Nyssomyia) hernandezi (Ortiz, 1965) is described, illustrated and measured their setas from specimens reared in the laboratory. By means of cluster analysis, it was shown the morphometric differentiation among larva of L. (N.) hernandezi and L (N.) antunesi, L. (N.) flaviscutellata, and L. (N.) anduzei. On the basis of these results, the morphological differentiation and possible relationships among the fourth instar larva of L. (N.) hernandezi and others Nyssomyia species, is discussed.
Introducción
Como bien lo señalan Young y Duncan (1994), las especies flebotominas neotropicales pertenecientes al subgénero Nyssomyia Barreto 1962 (= grupo intermedia Theodor 1965) del género Lutzomyia França 1924, abarcan alrededor de 22 especies y subespecies, cuya distribución geográfica va desde México, la América central, pasando por la isla caribeña de Trinidad, hasta llegar al Sur de América. Las mismas incluyen, L. (N.) flaviscutellata, L. (N.) trapidoi, L. (N.) hernandezi, L.(N.) waltoni, L. (N.) reducta, L. (N.) olmeca olmeca, L. (N.) o. bicolor, L. (N.) o. nociva, L. (N.) inornata, L. (N.) antunesi, L. (N.) whitmani, L. (N.) sylvicola, L. (N.) ylephiletor, L. (N.) edentula, L. (N.) richardwardi, L. (N.) intermedia, L.(N.) anduzei, L. (N.) umbratilis, L. (N.) shawi, L. (N.) bibinae, L. (N.) yuilli yuilli, y L. (N.) y. pajoti. Del diagnóstico morfológico del grupo resaltan la posesión de ascoides simples, palpómero 5 generalmente más corto que el 3; coxito con o sin setas persistentes; estilo con 4 espinas y sin seta subterminal; parámero sin divisiones y con setas simples; espermatecas de las hembras con cuerpos más largos que anchos, de forma anillada y con segmento terminal largo.
El hecho de que la mayoría de las especies que integran al subgénero Nyssomyia posean hábitos antropofílicos, hace que las mismas tengan una gran importancia epidemiológica, siendo consideradas varias especies como comprobadas, o se les ha incriminado, vectores de Leishmania al hombre (Grimaldi et al. 1989; W.H.O 1990). En este sentido, se tiene a L. (N.) flaviscutellata, L. (N.) reducta y L. (N.) ο. nociva como vectores de Le. (Leishmania) amazonensis (Shaw y Lainson 1968; Arias et al. 1987); a L. (N.) whitmani, L. (N.) anduzei y L. (N.) umbratilis de Le. (Viannia) guyanensis (Arias y Freitas 1978; Ryan et al. 1987); a L. (N.) intermedia y L. (N.) whitmani de Le. (V.) braziliensis (Gómez 1986; Vexenat et al. 1986); a L. (N.) o. olmeca, L. (N.) ylephiletor y L. (N.) flaviscutellata de Le. (Le.) mexicana (Biagi et al. 1967; Williams 1970; Porter et al. 1987); a L. (N.) trapidoi y L. (N.) ylephiletor de Le. (V.) panamensis (Zeledón et al. 1981; Zeledón 1985); a Lu. (N.) o. bicolor de Le. (Le.) venezuelensis (Bonfante- Garrido et al. 1987); y a L. (N.) antunesi de Le. (Le.) chagasi (Ryan et al. 1984).
De las 400 especies que integran al género Lutzomyia, apenas se conocen alrededor de un 15% de sus larvas (Young y Duncan, 1994); desafortunadamente, esto ha hecho que las relaciones de afinidades taxonómicas y sistemáticas entre las especies se haya realizado totalmente sobre caracteres morfológicos imaginales. Mientras que existen grupos de especies o subgéneros donde sólo se han descrito las larvas de una sola especie (e.g., en Micropygomyia Barreto 1962), llama la atención que de las 22 especies y subespecies que comprenden al subgénero Nyssomyia, se han ilustrado las larvas de casi el 36% de las mismas (Ward 1976; Young y Duncan 1994), incluyendo L. (N.) intermedia, L. (N.) olmeca bicolor, L. (N.) trapidoi, L. (N.) whitmani, L. (N.) ylephiletor (Barreto 1941; Hanson 1968), L. (N.) anduzei, L. (N.) antunesi y L. (N.) flaviscutellata (Ward 1976).
L. (N.) hernandezi posee una distribución restringida a Colombia en el Departamento de Norte de Santander (Young y Morales 1987) y Venezuela (Feliciangeli 1980). En este último país, se le ha capturado en los 3 estados andinos y en el Zulia (Feliciangeli 1988). Mediante la implementación de la técnica de inmunodifusión en geles de agarosa, González et al. (1991) y Añez et al. (1994) encontraron que L. (N.) hernandezi muestra apetencias alimentarias hacia aves, caprinos y ovinos; siendo capturada, además, por los primeros autores picando intradomiciliarmente sobre cebo humano. Estos hallazgos hablan a favor de considerar a L. (N.) hernandezi como una especie flebotomina con hábitos eurifágicos y oportunistas, y de que no debería, por lo tanto, ignorarse su papel en el mantenimiento de los ciclos enzoóticos, y su potencialidad como vector de Leishmania al hombre.
De acuerdo con la literatura revisada, aún no se han descrito los estadios preimaginales de L. (N.) hernandezi, desconociéndose la importancia de los mismos en la separación taxonómica con otras especies afines o morfológicamente indistinguibles, del subgénero Nyssomyia.
En el presente trabajo se describe e ilustra la quetotaxia de larvas de estadio IV de L. (N.) hernandezi. Así mismo, se dan las medidas de las setas, lo que permite realizar, de una manera preliminar y tentativa, la comparación morfométrica con otras tres especies del subgénero Nyssomyia, mediante la técnica multivariante de cluster análisis (CA).
Materiales y Métodos
El estudio se realizó con 3 larvas del estadio IV de primera generación, obtenidas a partir de distintas hembras de L. (N.) hernandezi capturadas por aspiración directa en huecos de árbol, en la población de "El Albarico" (entre los 9°20′N/ 70°50′W y 9°10′N/71°00′W), Municipio Monte Carmelo, del estado Trujillo, en la región andina de Venezuela.
Las metodologías para la obtención de las larvas de estadio IV de Añez y Oviedo (1985) y la clarificación y montaje se presentaron en un trabajo previo (Cazorla y Oviedo 1998). Se adoptó el sistema de Forattini (1973) para la numeración y nomenclatura de la quetotaxia.
Las estimaciones morfométricas se hicieron con un micrómetro ocular previamente calibrado, y las estructuras morfológicas se dibujaron con una cámara lúcida. Las medidas, todas expresadas en micras (µm), se presentan incluyendo la media aritmética y el rango de las mismas en paréntesis.
Cluster análisis
Para la realización de este análisis estadístico multivariante, se escogieron la media aritmética, que representa un individuo ubicado en el promedio poblacional, y el rango mínimo y máximo, representando dos individuos en los extremos, de las medidas de larvas IV de L. (N.) hernandezi (dadas en la Tabla), L. (N.) anduzei, L. (N.) antunesi y L. (N.) flaviscutellata. Las medidas de las tres últimas especies se tomaron del trabajo de Ward (1976), el cual fue realizado con material de Brasil.
Los caracteres morfológicos incluyen las setas
Las similitudes entre cada par de larvas se determinó por el coeficiente de distancia de Manhattan (city -block), construyéndose una matriz de distancia (12 x 12). A partir de esta última, se realizó un cluster análisis (CA), construyéndose un fenograma (dendrograma) por la técnica del ligamiento promedio de la media aritmética no ponderada (UPGMA), mediante el cual la admisión de un individuo (larva IV) dentro de un cluster se basa en el promedio de la distancia de ese individuo con los miembros existentes del cluster. Esto permitió estimar las relaciones fenéticas entre las cuatro especies mencionadas.
La descripción más detallada del CA está dada por Sneath y Sokal (1973) y Crisci y López (1983). Los datos se procesaron utilizando el sistema de programas computacionales STATISTICA (Stat Soft, Inc. 1993).
Resultados
Descripción de la quetotaxia larval de L. (N.) hernandezi
Longitud promedio del cuerpo, desde el margen anterior de la cápsula cefálica hasta el margen posterior del peine del noveno segmento abdominal, 3122 µm (3012 – 3232).
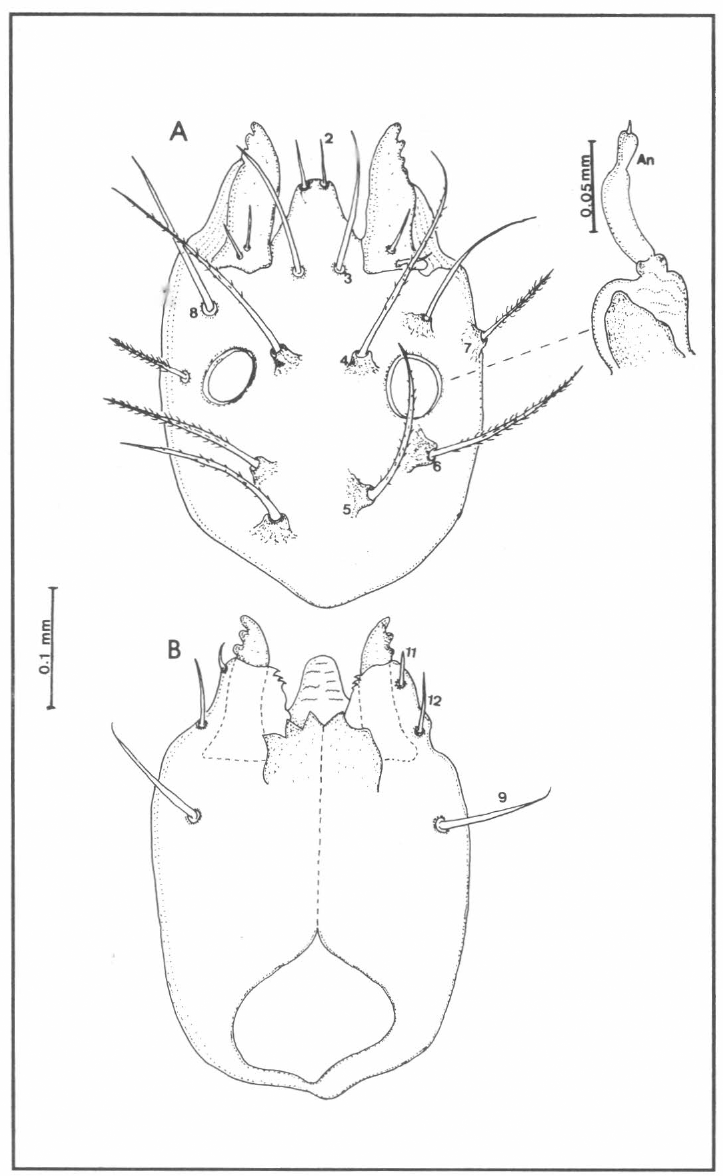
Quetotaxia de la región cefálica dorsal (A) y ventral (B) de larvas del estadio IV de Lutzomyia (N.) hernandezi; An= antena.
De forma ovoide, 305 μm (300 – 310) de largo; 240.3 µm (235 – 245.5) de ancho.
Setas 2 – 8 dorsales: Setas
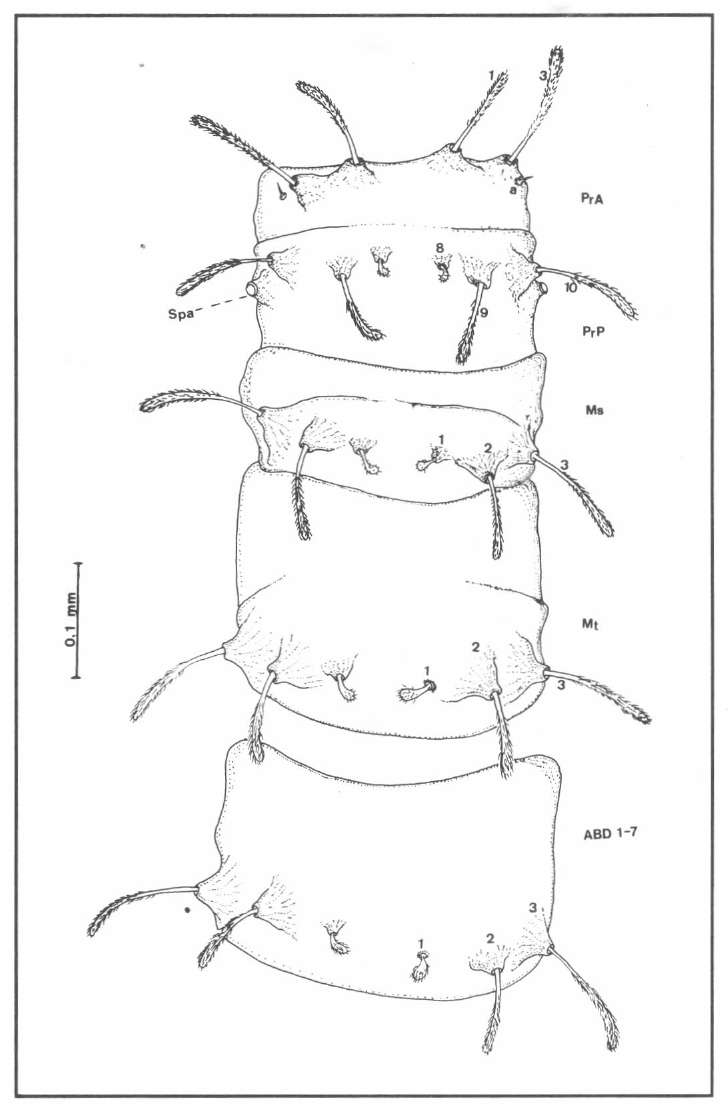
Quetotaxia de los segmentos torácicos y abdominales dorsales de larvas del estadio IV de Lutzomyia (N.) hernandezi; para= protórax anterior; PrP= protórax posterior; Ms= mesotórax; Mt= metatórax; ABD 17= segmentos abdominales 1 - 7; Spa= espiráculos anteriores.
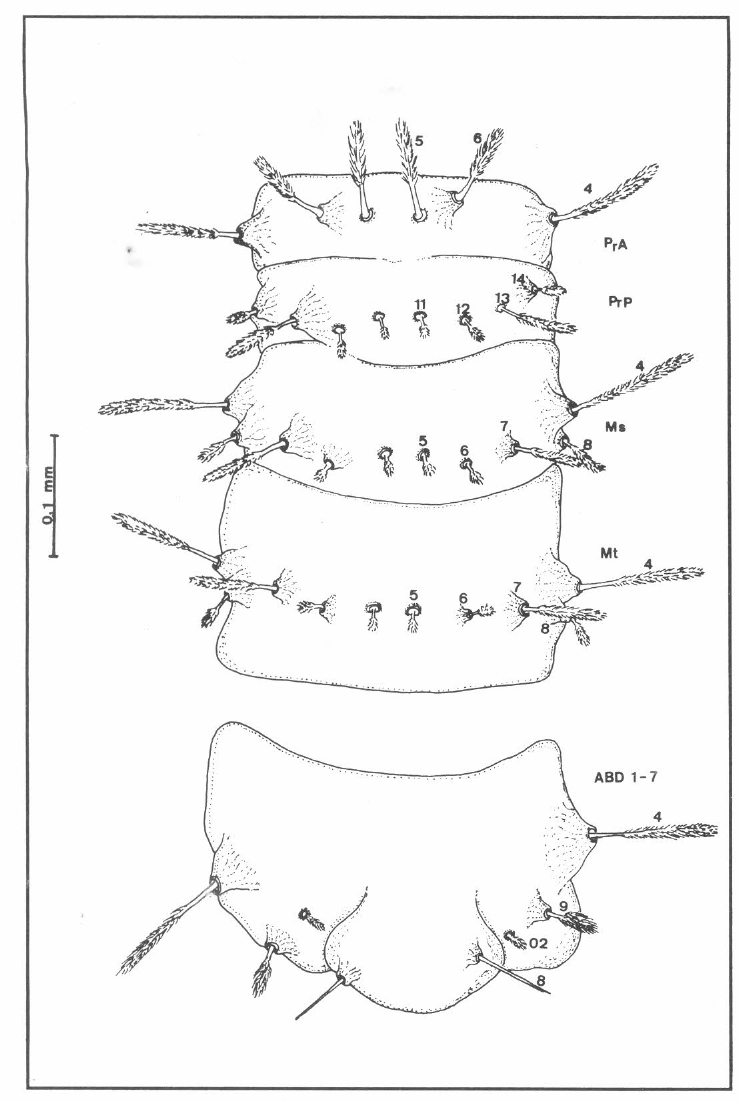
Quetotaxia de los segmentos torácicos y abdominales ventrales de larvas del estadio IV de Lutzomyia (N.) hernandezi. Abreviaturas como en figura 2.
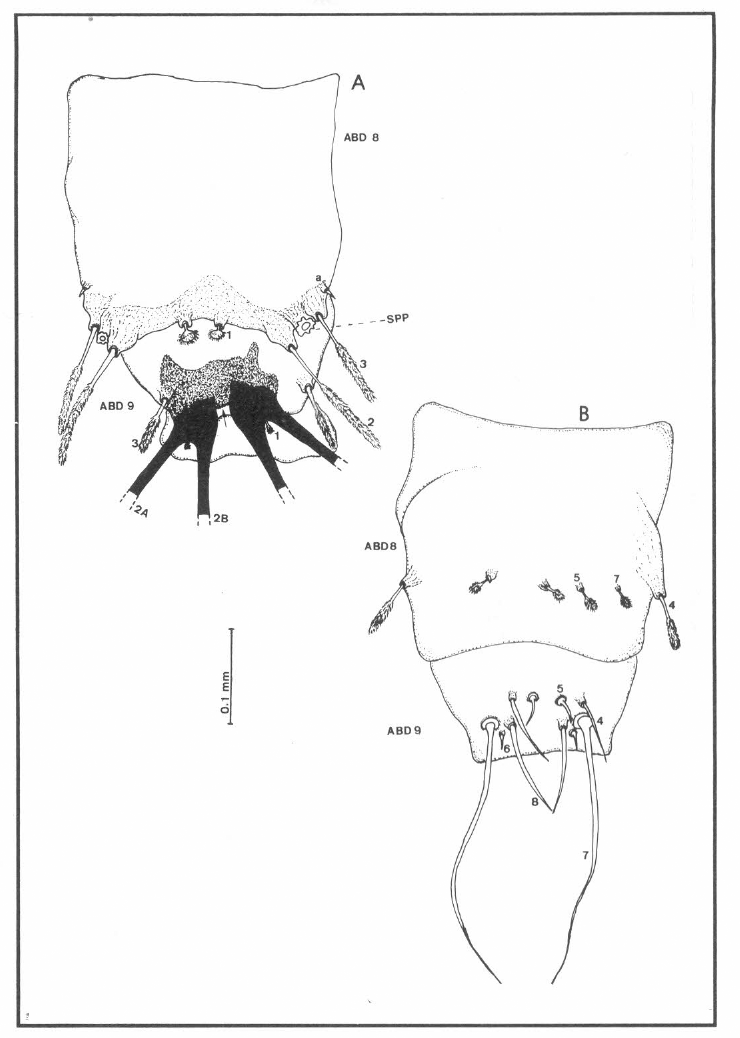
Quetotaxia de los segmentos abdominales 8 y 9 dorsales (A) y ventrales (B) de larvas del estadio IV de Lutzomyia (N.) hernandezi; ABD 8= segmento abdominal 8; ABD 9= segmento abdominal 9; 2A y 2B= setas caudales. SPP= espiráculos posteriores. La flecha en A señala los dientes en el segmento abdominal 9.
En la Tabla 1 se presentan las medidas, con media aritmética y rangos, de las setas descritas.
Medias y rangos* (µm) de las longitudes de setas del IV estadio larval de Lutzomyia (N.) hernandezi (n=3)
(-): seta ausente.
las setas sin rango exhibieron las tres mediciones iguales.
= sólo se midieron las setas de los segmentos 1 y 7.
Cluster análisis (CA)
La figura 5 muestra los resultados del CA; la abscisa se utiliza para separar las larvas IV de los flebotominos, mientras que la ordenada muestra la escala de distancia desde 0 hasta 800. Así por ejemplo, los individuos 7 y 8 de L. (N.) hernandezi se encuentran distantes a 10, y su distancia promedio con el individuo 9 de su misma especie es alrededor de 20. Cuando la distancia es cerca de 250, hay tres clusters representados por los individuos de L. (N.) antunesi, L. (N.) anduzei y L. (Ν.) hernandezi. En definitiva, en el fenograma se detectan cuatro grupos fenéticos (cluesters) bien definidos: L. (N.) anduzei (1 – 3); L. (N.) antunesi (4 – 6); L. (N.) hernandezi (7 – 9) y L. (N.) flaviscutellata (10 - 12), aunque las larvas de esta última especie mostraron la diferenciación morfométrica más distante con respecto a las restantes especies, formando un cluster relativamente aislado.
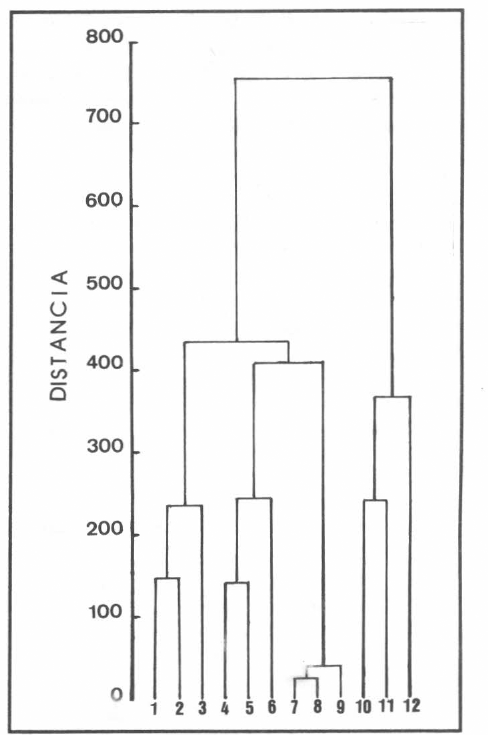
Fenograma de larvas IV de Lutzomyia (N.) anduzei (1-3), L. (N.) antunesi (4-6), L. (N.) hernandezi (7-9) y L. (N.) flaviscutellata (10-12), obtenido por la técnica de UPMGA y basado en la matriz de distancia del coeficiente de Manhattan. La escala se da en valores de distancia.
Discusión
Como bien lo señalan Young y Duncan (1994), los machos de L. (N.) hernandezi y L.(N.) sylvicola, especie restringida a la Guyana Francesa y Brasil, son morfológicamente indistinguibles, arrojando la posibilidad de que ambas sean conespecíficas. Desafortunadamente, aún no se han descrito los estadios pre-imaginales de L. (N.) sylvicola, por lo que no es posible realizar comparaciones morfológicas ni morfométricas con los de L. (N.) hernandezi.
Por otra parte, los imagos de L. (N.) hernandezi están muy relacionados morfológicamente con los de L. (Ν.) whitmani, especie flebotomina distribuida en la Guyana Francesa, Brasil, Perú, Paraguay y Argentina. En las claves dicotómicas elaboradas para Nyssomyia por Young y Duncan (1994), la separación específica entre las mismas es realizada basándose en las diferencias morfométricas de caracteres internos del aparato reproductivo de ambos sexos. Los estadios inmaduros de L. (N.) whitmani fueron descritos por Barreto (1941), quien considera, al igual que Forattini (1973), sus larvas y las de L. (N.) intermedia (distribución: Brasil, Paraguay y Argentina) son morfológicamente indistinguibles, teniendo ambas el último segmento antenal en forma de banana, lo que las relaciona con L. (N.) hernandezi. A pesar de que no hay datos de tipo morfométrico, una de las diferencias resaltantes entre las larvas IV de L. (N.) hernandezi y estas dos especies señaladas, es la ausencia en las larvas de ésta de la seta
Con respecto a L. (N.) trapidoi (distribución: Centroamérica, Colombia y Ecuador) y L. (N.) ylephiletor (distribución: México, Centroamérica y Colombia), cuyas larvas son muy parecidas morfológicamente, se tiene en primer término que aunque el segmento antenal distal de las mismas es en forma de banana, lo que las relaciona con L. (N.) hernandezi, sus segmentos antenales proximales son tangiblemente más largos que los de ésta, a juzgar por las ilustraciones de Hanson (1968). Además, las larvas de L. (N.) hernandezi no presentan la seta
Tomando como referencia a Hanson (1968), las larvas de L. (N.) olmeca bicolor (distribución: Costa Rica, Panamá, Colombia, Perú, Ecuador y Venezuela) exhiben la cabeza de forma oval-redondeada, con proyecciones angulares y tubérculos antenales de mayor longitud que sobresalen del margen cefálico, a diferencia de la de L. (N.) hernandezi que es básicamente ovalada, y con tubérculos antenales que virtualmente no sobresalen del margen cefálico. Además, su segmento antenal distal no presenta curvatura y es digitiforme, y el proximal es subigual a este último, mientras que los de L. (Ν.) hernandezi son de forma de banana y muy cortos, respectivamente. La seta
La diferenciación entre las larvas de L. (N.) flaviscutellata (distribución: Colombia, Guyana Francesa, Surinam, Brasil, Ecuador, Perú, Bolivia, Trinidad y Venezuela), tal como fueron descritas por Ward (1976), y L. (N.) hernandezi sigue básicamente los lineamientos señalados para L. (N.) olmeca bicolor.
En relación con las larvas IV de L. (N.) antunesi y L. (N.) anduzei, especies cuya distribución abarca entre otros países a Venezuela, las mismas poseen la seta
En la descripción de algunos estadios inmaduros de flebotominos de Brasil, Ward (1976) llamó la atención sobre la dificultad de distinguir entre larvas de especies flebotominas muy afines morfológicamente (L. (N.) intermedia y L. (N.) whitmani; L. (N.) trapidoi y L. (N.) ylephiletor; L. (Psychodopygus) panamensis y L.(Ps.) pessoana), sugiriendo la necesidad de realizar medidas de las setas para poder cuantificar las posibles diferencias entre las mismas.
En el presente trabajo se inician, de una manera preliminar y tentativa, los estudios comparativos en este sentido. En efecto, el análisis multivariante por CA de la variación morfométrica de 32 setas larvales de las especies L. (N.) antunesi, L. (N.) anduzei, L. (N.) flaviscutellata y L. (N.) hernandezi sugiere que las larvas IV de estas especies se diferencian significativamente. Estos estudios cobran mayor relevancia cuando se tiene que aún por alta resolución con la Microscopía Electrónica de Barrido (MEB), Feliciangeli et al. (1993) no detectaron diferencias aparentes entre los patrones exocoriónicos de huevos de L. (N.) flaviscutellata y L. (N.) olmeca bicolor.
De acuerdo con los criterios de Leite y Williams (1996) de reunir las larvas de estadio IV de los flebotominos neotropicales en cuatro grupos, L. (N.) hernandezi se ubicaría en el Grupo 2, con tubérculo antenal en forma de cono truncado, con segmento antenal basal corto y el distal largo y en forma de banana. Con la excepción de L. (N.) flaviscutellata y L. (N.) olmeca bicolor, las restantes especies del subgénero Nyssomyia, a las cuales se les conocen sus estadios inmaduros, concuerdan con esta agrupación.
Las especies del subgénero Psychodopygus exhiben sus segmentos antenales distales digitiformes, conformando de este modo el grupo 3. L. (N.) flaviscutellata y L. (N.) olmeca bicolor caerían igualmente en este grupo 3, a juzgar por la morfología de sus antenas. Por otra parte, el patrón exo-coriónico de los huevos de ambas especies poseen mayor afinidad morfológica con el detectado en los de Psychodopygus (Ward 1976; Feliciangeli et al. 1993).
Estos hallazgos parecieran encontrar apoyo en los resultados obtenidos en el presente estudio, al detectarse mediante el análisis multivariante por CA, que las larvas de L. (N.) flaviscutellata exhiben poca afinidad fenética con las de L. (N.) antunesi, L. (N.) anduzei y L. (N.) hernandezi. Así mismo, se apoyaría la tesis de Ward (1976), según la cual las larvas de L. (N.) flaviscutellata y L. (N.) olmeca bicolor tienen mayor similitud morfológica con aquellas del subgénero Psychodopygus que con las del subgénero Nyssomyia.
Los comentarios anteriores ponen de manifiesto, una vez más, la incongruencia entre los esquemas taxonómicos basados sobre caracteres imaginales y los hallazgos taxonómicos hechos sobre los estadios pre-imaginales. El mismo Ward (1976) llegó a la conclusión de que las larvas del subgénero Psychodopygus no parecieran guardar congruencia con el agrupamiento en series de los adultos propuesto por Theodor (1965).
En este mismo sentido, Leite y Williams (1996; 1997) detectan con MEB nuevas características morfológicas en los estadios inmaduros de L. (Lutzomyia) longipalpis, especialmente en el primer ínstar y sugieren que las mismas pudieran ser distintivas aún al nivel de género.
Finalmente, se insiste, desde un primer artículo (Cazorla y Oviedo 1998), en la necesidad de homogenizar los sistemas actuales de nomenclatura y numeración de la quetotaxia de los flebotominos. Esta necesidad se basa en la dificultad que, muchas veces, tienen que enfrentar los investigadores cuando aplican algunos sistemas existentes para describir la quetotaxia de otros subgéneros distintos a aquellos para los cuales fueron creados dichos sistemas. En este sentido, Ward (1976) ideó un sistema de nomenclatura y numeración adaptado en su momento para estudiar larvas de los subgéneros Nyssomyia y Psychodopygus. El mismo autor hizo la salvedad de que su sistema podría resultar exclusivamente útil para el estudio de ambos subgéneros. Esta preocupación se hizo patente veinte años después, cuando Leite y Williams (1996) encontraron dificultades para describir las larvas IV de una especie del subgénero Lutzomyia (L. Longipalpis) al detectar por MEB sensilas en el tegumento de las mismas, no descritas por Ward (1976).
Conclusiones
Siguiendo la clasificación de Leite y Williams (1996), L. (N.) hernandezi se ubica en el Grupo 2, como la mayoría de las especies del subgénero Nyssomyia.
El análisis multivariante por eA sugiere, de una manera preliminar y tentativa, que las larvas IV de L. (N.) hernandezi se diferencian morfométricamente de las de L. (N.) antunesi, L. (N.) anduzei y L. (N.) f7aviscutellata.
Las larvas IV de L. (N.) hernandezi se diferencian morfológicamente de las de las restantes especies descritas del subgénero Nyssomyia.
Se necesita homogenizar los actuales sistemas de nomenclatura y numeración de la quetotaxia de los flebotominos.
Footnotes
Agradecimientos
Al TSU R. Barazarte por su asistencia técnica en el campo.