
Abstract
Llanera locust, Rhammatocerus schistocercoides (Rehn), represents a serious threat for agriculture and livestock in the Eastern savannas of Colombia. A native isolate of the entomopathogenic fungi Metarhizium anisopliae, grown in rice and formulated, has been successfully used for locust control under field conditions. The effect of the culture medium over fungi virulence has been reported in the scientific literature. The objective of the present work was to evaluate the effect of some potential virulence inductors over biological and enzymatic activity of the M. anisopliae conidia. For this purpose, the isolate Mt004 previously selected by its high virulence against R. schistocercoides and the isolate Mt006 of M. anisopliae which presented the lowest biocontrol activity, were grown in a shaken liquid medium and in solid medium containing different potential virulence inductors (colloidal chitin, locust wings and legs, wheat bran, Rhizoctonia solani dead mycelia and Aspergillus niger dead mycelia). Conidia produced in presence of virulence inductors were evaluated under laboratory conditions, against adult locust. The maximum mortality in locust was caused when the fungi was grown in liquid medium supplemented with an homogenized powder of locust wings and legs, this strain presented a lethal time 50 (LT50) of 11 days and 70% of total mortality, 15 days after treatment. Otherwise, Mt004 isolate presented the highest biocontrol activity when the fungi was grown in liquid medium supplemented with both homogenized powder of locust wings and legs and wheat bran. This strain presented a LT50 of 5,8 days and 7,2 days respectively and a total mortality of 100% and 100% respectively, 13 and 14 days after treatment. This result suggest that these substrates have a virulence induction effect. Enzymatic activity was higher when the culture was made in liquid medium than solid medium and when it was supplemented with homogenized powder of locust wings and legs and wheat bran. The biological activity presented a direct relation with the protease activity of PR1. The found results let conclude that this enzyme is directly related with fungus virulence.
Introducción
En el año 1994 en la Orinoquía colombiana se observó un incremento alarmante de la población de langostas, las cuales fueron identificadas como Rhammatocerus schistocercoides (Rehn 1906). Esta plaga se encuentra distribuida en los ecosistemas tropicales conocidos como sabanas, que están localizados en el interior del continente (León 1996).
Los registros actuales han mostrado daños en praderas nativas y pastos mejorados. Sin embargo, se considera que esta plaga tiene la potencialidad para consumir cultivos como arroz, maíz, sorgo y soya (León 1996). Aunque en estos momentos la plaga se encuentra concentrada en el departamento del Vichada, es una amenaza potencial para la agricultura y la ganadería de los Llanos orientales de Colombia.
Para su control, Corpoica desarrolló una formulación a base de Metarhizium anisopliae, para la cual, la biomasa es producida masivamente utilizando un método semiartesanal de multiplicación en arroz. La formulación consiste en un polvo para reconstituir en una emulsión aceite en agua. Dentro del polvo formulado, se encuentran los conidios del hongo encapsulados con filtros solares que le brindan un 100% de protección frente a la radiación ultravioleta (Gómez y Villamizar 1996). Dicho producto ha demostrado su eficiencia en campo en Carimagua (Meta), alcanzando niveles de hasta el 70% de control de la plaga (Espinel 1998).
Varios componentes cuticulares de insectos hospederos, al ser incorporados al medio de cultivo, producen un aumento considerable de la virulencia de los conidios (Bidocha y Khachatourians 1992). Este incremento en la actividad patogénica posiblemente se debe a la activación por parte del sustrato de sistemas enzimáticos en los hongos, los cuales son determinantes en su patogenicidad, por lo que se hace necesario buscar inductores de virulencia que además permitan conocer mejor el mecanismo de acción de los hongos entomopatógenos.
Materiales y Métodos
Los entomopatógenos que se utilizaron fueron la cepa de Metarhizium anisopliae Mt004 proveniente de Rionegro (Antioquia), aislada de Ancognata scarabaeoides (Ericshon) y la cepa de M. anisopliae Mt006 aislada de Anthonomus sp. en Venezuela. Estas cepas se seleccionaron para este trabajo, dado que en estudios previos mostraron diferencias contrastantes en la virulencia sobre R. schistocercoides; es así como la cepa Mt004 tuvo la actividad patogénica mayor frente a la plaga (Espinel 1998), mientras que la cepa Mt006 presentó un control deficiente de la langosta Ilanera (Zamora 1996). Estos aislamientos se encuentran conservados en el banco de cepas del Laboratorio de Control Biológico del Programa Nacional de Manejo Integrado de Plagas de Corpoica en su sede C.I. Tibaitatá.
Para la producción de conidios sumergidos se utilizó un medio líquido indicado por Villamizar (1998), el cual fue seleccionado por permitir la obtención de conidios bajo estas condiciones de cultivo. Los medios se suplementaron con los sustratos potencialmente inductores de virulencia (quitina coloidal, homogeneizado de alas y patas de langosta, salvado de trigo, micelio muerto de Rhizoctonia solani y micelio muerto de Aspergillus niger), éstos se prepararon de acuerdo con los procedimientos descritos por Villamizar (1998) y fueron adicionados al medio líquido seleccionado, antes de su esterilización en una concentración de 0.40%. En erlenmeyers de 250 ml de capacidad que contenían 100 ml de cada medio de cultivo líquido a evaluar, se inoculó 1 ml de suspensión de conidios de M. anisopliae de las cepas Mt004 o Mt006 ajustada a 1 x 106 conidios/ml. Los medios se incubaron a 25°C y 155 rpm durante 10 días.
Para separar del medio líquido los conidios que iban a ser utilizados para el bioensayo y en las pruebas enzimáticas, se filtró el medio de cultivo a través de una muselina estéril. El filtrado se centrifugó a 4000 rpm durante 10 minutos, se retiró el sobrenadante y el sedimento se resuspendió en una solución de Tween 80 a 0.2% y se centrifugó nuevamente. Este lavado se realizó dos veces sucesivas. Los conidios se secaron en estufa con corriente de aire a 25°C durante 24 horas.
Para el cultivo de M. Anisopliae, en medio sólido, se utilizó arroz humedecido estéril contenido en bolsas de polietileno de densidad alta, siguiendo los procedimientos descritos por Gómez y Villamizar (1996). Este medio fue suplementado con los inductores de virulencia. Cada sustrato fue adicionado en forma de polvo al medio de arroz antes de su esterilización en una concentración de 0.4% en peso. Para preparar el medio de cultivo, se introdujeron 100 g de arroz, 80 ml de agua y 0.4 g de inductor en cada bolsa. Las bolsas estériles se inocularon con 1ml de suspensión de conidios de M. anisopliae ajustada a una concentración de 4 x 107 conidios/ml. Estos medios se incubaron a 25°C durante 15 días en presencia de luz constante.
Con el fin de separar los conidios del medio sólido, una vez terminado el tiempo de incubación, se realizó un lavado del arroz esporulado con una solución de Tween 80 a 0,2%. Posteriormente, esta suspensión se centrifugó a 4000 rpm, durante 10 minutos. Los conidios se secaron en estufa con corriente de aire a 25°C durante 24 horas.
Los conidios obtenidos, tanto de los cultivos líquidos como de los cultivos sólidos, se utilizaron para preparar las suspensiones requeridas en el desarrollo del bioensayo utilizando una solución de Tween 80 a 0.2%. Para todos los tratamientos la suspensión de M. anisopliae se ajustó a una concentración de 6 x 105 conidios/ml, de manera que al aplicar 20 µl de suspensión por insecto se ajustó la dosis letal 90 determinada sobre R. schistocercoides para la cepa Mt004 por Espinel (1998), que correspondió a 7 conidios por insecto.
Como controles positivos para los medios líquidos y sólidos se utilizaron respectivamente los conidios del hongo producidos en medio líquido y en arroz sin inductores de virulencia.
Los insectos utilizados en este ensayo, fueron langostas adultas recolectadas en campo en Carimagua (Meta). Estos fueron transportados y mantenidos en cuarentena durante 6 días antes de ser utilizados para los bioensayos. En el ensayo se tuvo un testigo tratado, que consistió en insectos tratados tópicamente con una solución de Tween 80 a 0.2% y un testigo absoluto en el cual no se realizó ningún tratamiento de los individuos.
La aplicación del hongo a los insectos se hizo en forma tópica, colocando 20 µl de la suspensión del hongo sobre el protórax de cada insecto. Se inocularon 20 individuos por tratamiento y se colocaron en una jaula dotada de alimento y agua. Las jaulas se ubicaron en un cuarto de incubación a 30°C y 60% de humedad relativa. Cada tratamiento contó con tres repeticiones ubicadas bajo un diseño experimental completamente al azar, la unidad experimental fue una jaula con 20 individuos y la unidad de muestreo fue un insecto muerto.
Diariamente se colocó alimento (pasto kikuyo) y agua en cada jaula y se recogieron los individuos muertos, los cuales se situaron en cámaras húmedas que se incubaron a 25°C hasta que se observara esporulación, para confirmar la infección fúngica.
El registro de datos se realizó diariamente hasta que el testigo absoluto llegara a un 13% de mortalidad o hasta que todos los individuos de los tratamientos hubieran muerto.
Los resultados correspondientes a la tasa de mortalidad acumulada se transformaron en porcentajes y posteriormente se calculó el porcentaje de mortalidad corregida, mediante la fórmula de Abbot (CIBAGEIGY 1978):
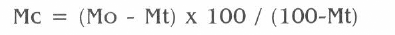
donde Mo es el porcentaje de mortalidad observado en el tratamiento y Mt es el porcentaje de mortalidad observado en el testigo tratado.
A partir de los resultados de los bioensayos, se determinaron los tiempos letales medios (TL50) correspondientes a cada tratamiento.
Los resultados del experimento se sometieron a un análisis de varianza y una prueba de comparación de rangos múltiples de Duncan.
Para los ensayos de cuantificación de la actividad enzimática se utilizaron conidios producidos de la misma forma que los utilizados en los biensayos. A partir de ellos, se hizo un extracto enzimático con solución de Tween 80 a 2% tamponada a pH 7, siguiendo los procedimientos descritos por St Leger et al. (1991).
La cuantificación de la quitinasa Nacetilglucosaminidasa (NAGasa) se realizó según lo recomendado por St Leger et al. (1986). A cada 0.2 ml de extracto enzimático, se le adicionó 1ml de pnitrofenil-N-acetilglucosaminida (1mg/ml amortiguado a pH 5 con tampón de citratos 0,1M). Esta mezcla se incubó a 35°C por 30 minutos. Finalizado el tiempo de incubación, se adicionó 1,5 ml de NH₂OH 1N con EDTA disódico en una concentración equivalente a 2 mM. Se leyó la absorbancia en espectrofotómetro a 400 nm, estos resultados se extrapolaron en una curva de calibración de absorbancia versus concentración de p-nitrofenol y los resultados se expresaron en µmoles de pnitrofenol liberadas por miligramo de conidio.
La cuantificación de la actividad de la quimoelastasa proteasa PR1 se realizó siguiendo los procedimientos recomendados por St Leger et al. (1986). A partir de los extractos de conidios, se tomaron 0,2 ml del sustrato 1mM succinil-ala-ala-propilfenialanina-p-nitroanilida disuelto en dimetilsulfóxido a los que se les adicionaron 0,2 ml de extracto enzimático, 2,5 ml de solución tampón Tris-HCl (2mM, pH = 8), el cual contenía KCl en una concentración equivalente a 0,3 M. Inmediatamente después se leyó la absorbancia de esta mezcla en espectrofotómetro a 410 nm. Los resultados se extrapolaron en una curva de calibración de absorbancia versus concentración de p-nitroanilina y los resultados se expresaron en µmoles de p-nitroanilina liberadas por miligramo de conidio.
La determinación de la actividad esterasa se hizo de acuerdo con las recomendaciones de St Leger et al. (1986a). La reacción enzimática se llevó a cabo tomando 0.2 ml de extracto enzimático y adicionándoles 1 ml de p-nitrofenil-caprilato (1mg/ml en dimetilsulfóxido). Esta mezcla se incubó a 35°C durante 30 minutos. Posteriormente, se adicionaron 1,5 ml de NH₂OH 1N con EDTA disódico en una concentración equivalente a 2 mM. Se leyó la absorbancia en espectrofotómetro a 400 nm. La actividad enzimática se determinó extrapolando la absorbancia obtenida en una curva de calibración de absorbancia versus concentración de p-nitrofenol y los resultados se expresaron en µmoles de p-nitrofenol liberadas por miligramo de conidio.
Para todas las determinaciones de la actividad enzimática correspondiente a NAGasa, esterasa y PR1 se cultivó M. anisopliae en presencia de los sustratos seleccionados por duplicado y a partir de cada uno de los cultivos se tomaron tres muestras de conidios; con cada una de ellas se realizaron los extractos a partir de los cuales se hizo la determinación de actividad enzimática. Los 6 valores obtenidos para cada tratamiento se promediaron y sometieron a un análisis de varianza con un α=0.05 y las diferencias significativas entre tratamientos se determinaron utilizando la prueba de comparación múltiple de Duncan.
Resultados y Discusión
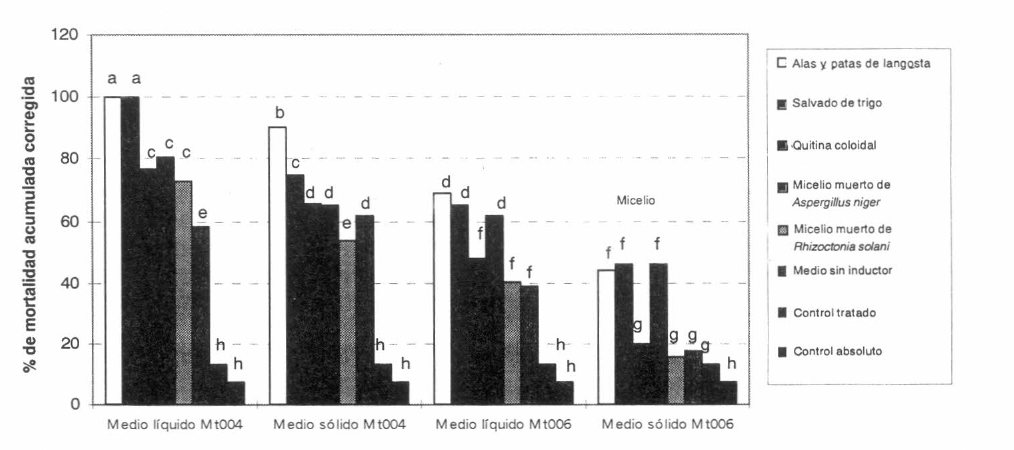
Cuando los conidios correspondientes a las cepas Mt004 y Mt006 de M. anisopliae, provenientes de los medios líquidos que contenían cada uno de los potenciales inductores de virulencia, se utilizaron para su aplicación tópica sobre langostas adultas a una concentración de 6 x 105 conidios/ml, se encontraron para una misma cepa diferentes porcentajes de mortalidad acumulada en los insectos, relacionados con el tipo de inductor adicionado al medio de cultivo. En todos los casos, siendo inferiores los ocasionados por la cepa Mt006 (Tabla 1; Fig. 1).
En este ensayo, en los insectos testigo se encontraron porcentajes de mortalidad bajos, debido a que el testigo tratado presentó una mortalidad acumulada de 13%, 15 días después de iniciado el bioensayo y el testigo absoluto presentó en este mismo tiempo una mortalidad acumulada de 7.2%. Estos valores de mortalidad en los testigos son aceptables para ensayos biológicos y se atribuyen a factores tales como el "stress" causado por el confinamiento de las langostas, el cambio en su alimentación y el efecto de las condiciones ambientales.
Actividad patogénica de dos cepas de M. anisopliae cultivados en medio líquido y sólido en presencia de varios potenciales inductores de virulencia.
Tiempos letales medios (TL50) obtenidos en el control de Rhammatocerus schistocercoides con dos cepas de Metarhizium anisopliae multiplicadas en medios líquidos y sólidos suplementados con diferentes potenciales inductores de virulencia
* Los resultados seguidos por la misma letra no presentan diferencias significativas según la prueba estadística de Duncan
Cuando la cepa Mt004, seleccionada en trabajos previos por su actividad patogénica alta fue cultivada en el medio de cultivo líquido que contenía salvado de trigo y homogeneizado de alas y patas de langosta, produjo el 100% de mortalidad acumulada 15 días después de iniciado el ensayo, no presentando diferencias significativas entre sí, pero sí con respecto a todos los demás tratamientos. El efecto biocontrolador obtenido con los conidios producidos en presencia de micelio muerto de A. niger, de micelio muerto de R. solani y de quitina coloidal fue significativamente menor que el obtenido con la biomasa multiplicada en presencia de salvado de trigo y de homogeneizado de alas y patas de iangosta, pero superior y significativamente diferente al obtenido con el inóculo proveniente del cultivo sin inductores, indicando que todos los sustratos evaluados, aunque en diferente medida, ejercen un efecto inductor de la virulencia. Cuando esta cepa se cultivó en medios suplementados con micelio muerto de Aspergillus niger, produjo una mortalidad de 80.7%, cuando se cultivó en presencia de micelio muerto de Rhizoctonia solani, la mortalidad obtenida fue de 73% y cuando se cultivó en presencia de quitina coloidal se produjo una mortalidad de 77%, tratamientos que no fueron significativamente diferentes entre sí, pero si lo fueron cuando no se adicionó ningún inductor al medio cuya mortalidad fue de 58.1% y el TL50 de 13.3 días.
Estos resultados indican que el salvado de trigo y el homogeneizado de alas y patas de langosta tienen un efecto inductor sobre la virulencia del hongo M. anisopliae además porque con estos tratamientos se obtuvieron los menores tiempos letales medios (TL50), los cuales fueron de 7,2 días y de 5,8 días, respectivamente. El aumento en la actividad patogénica encontrada en el presente trabajo, cuando el hongo se cultivó en presencia de alas y patas de langosta, estuvo relacionado con el aumento en la virulencia registrada en otros trabajos con Beauveria bassiana por Bidochka y Khachatourians (1992) cuando cultivaron el hongo en presencia de componentes cuticulares de su hospedero Melanoplus sanguinipes. La pérdida de virulencia en un hongo que ha sido repicado repetidamente en medios de cultivo, ha sido relacionada por algunos investigadores con la disminución de la actividad enzimática de los conidios, debido a que las enzimas se expresan en menor cantidad a medida que el microorganismo se cultiva en ausencia de sustratos para éstas, los cuales generalmente están presentes en el insecto hospedero. Si se tiene en cuenta que las alas y patas de langosta están compuestas principalmente por quitina, ceras y proteínas, se podría sugerir que este sustrato estimula la producción de enzimas por parte de los conidios, las cuales son capaces de degradar dichos compuestos y que estos conidios cuya actividad enzimática ha sido inducida, tendrían más contra las langostas.
De otra parte, el salvado de trigo, ha sido señalado como un inductor de enzimas tales como glucanasas, quitinasas y celulasas (Thonart et al. 1991). Teniendo en cuenta que la producción de estas enzimas puede ser concomitante con la producción de otras que se encuentran en la membrana celular, se podría pensar que en presencia de salvado, además de las mencionadas enzimas se podrían haber producido otras que tienen actividad contra el insecto.
Cuando se evaluó la actividad patogénica de la cepa Mt006, cultivada en medios líquidos que contenían los potenciales inductores de virulencia, se encontró en todos los casos, significativamente menor mortalidad en las langostas que la encontrada con la cepa Mt004 (Tabla 1, Fig. 1). Este resultado está relacionado con lo registrado por Espinel (1998) y por Zamora (1996) quienes demostraron que las cepas Mt004 y Mt006 tenían, respectivamente alta y baja actividad patogénica contra R. schistocercoides.
Al inocular langostas con la cepa MT006 no se alcanzó el 100% de mortalidad acumulada con ninguno de los tratamientos después de 15 días de iniciado el ensayo, lo cual confirma que este aislamiento es menos patogénico a este insecto. La mayor actividad patogénica se obtuvo cuando la cepa Mt006 creció en medio líquido en presencia de alas y patas de langosta, obteniéndose un TL50 de 11,8 días y una mortalidad acumulada a los 15 días de iniciado el ensayo de 69,2%. Esta actividad patogénica no fue significativamente diferente de la observada con conidios provenientes del medio líquido suplementado con salvado de trigo y con micelio muerto de A. niger. En estos casos, se obtuvieron porcentajes de mortalidad acumulada de 65.3% y 61.5% y TL50 de 12.4 días y de 11.8 días respectivamente. Estos resultados fueron significativamente diferentes de los obtenidos en presencia de los demás inductores y en ausencia de éstos, lo cual indicaría que el salvado de trigo, el micelio muerto de A. niger y el homogeneizado de alas y patas de langosta tienen un efecto inductor sobre la virulencia de este aislamiento de M. anisopliae.
Los conidios del aislamiento Mt006 producidos en los medios líquidos sin inductor y suplementados con quitina coloidal y con micelio de R. solani presentaron significativamente menor actividad patogénica que los obtenidos en medios suplementados con otros potenciales inductores; éstos produjeron mortalidades acumuladas de 38.8%, 48% у 40.3%, respectivamente, resultados que no fueron significativamente diferentes entre sí, lo cual podría indicar que el micelio muerto de R. solani y la quitina coloidal no tienen efecto en la actividad patogénica de este aislamiento.
Cuando se cultivaron las cepas Mt004 y Mt006 en medio sólido, se encontraron las mismas tendencias que cuando se cultivaron en medios líquidos. Sin embargo, cuando el sustrato de crecimiento del hongo fue arroz, la mortalidad observada en las langostas, fue inferior a la obtenida cuando éste fue cultivado en medios líquidos. Al cultivar la cepa Mt004 en medio sólido, el mayor porcentaje de mortalidad acumulada se alcanzó con los conidios producidos en presencia de homogeneizado de alas y patas de langosta, seguido por el obtenido con los conidios provenientes del cultivo suplementado con salvado de trigo, tratamientos que fueron significativamente diferentes entre ellos y diferentes de los restantes. Cuando el hongo se cultivó en presencia de alas y patas de langosta, se obtuvo un porcentaje de mortalidad de 90.3% y un TL50 de 10.8 días, mientras que cuando se cultivó en presencia de salvado de trigo, la mortalidad obtenida en las langostas fue de 75% y el TL50 fue de 12.9 días.
Cuando el aislamiento Mt004 se cultivó en el medio sin inductor y en los medios suplementados con micelio muerto de R. solani, con quitina coloidal y con micelio muerto de A. niger, las mortalidades acumuladas fueron de 61.5, 53.8, 65.4 y 65.3%. El análisis de varianza no detectó diferencias significativas entre estos tratamientos, resultado que indica que estos suplementos no tuvieron un efecto inductor en la virulencia del microorganismo.
Cuando la cepa Mt006 de M. anisopliae se cultivó en medio sólido se obtuvieron niveles de control muy bajos y en ningún caso la mortalidad obtenida superó el 50% (Tabla 1; Fig. 1), siendo los conidios provenientes de medios suplementados con micelio muerto de A. niger, con salvado de trigo y con homogeneizado de alas y patas de langosta, los que produjeron en las langostas las mortalidades mayores (Tabla 1).
Para una misma cepa se observaron diferencias significativas entre la actividad patogénica de los conidios producidos en medio líquido y los producidos en medio sólido. Siendo los conidios más virulentos cuando el microorganismo se cultivó en medio líquido, esto podría ser debido a la distribución homogénea del sustrato inductor en el medio líquido, lo que permitiría un mejor aprovechamiento del mismo por parte del hongo entomopatógeno; mientras que en el medio sólido el potencial inductor no queda bien distribuido, si se tiene en cuenta que la cantidad usada es baja (0.40%). Esta concentración baja permitiría que sólo algunos conidios puedan utilizarlo. En los cultivos líquidos, los suplementos ejercen un efecto inductor en la virulencia de los conidios debido a qué los constituyentes de éstos sirven de sustrato a diferentes enzimas encargadas de la actividad patogénica, de tal forma que al encontrarse presentes en el medio, inducen la producción de estas enzimas. Por el contrario, cuando el medio de cultivo estaba constituido únicamente por arroz, sustrato cuya composición básica es almidón, se estimularía la producción de enzimas capaces de degradarlo, lo que a su vez podría reprimir la síntesis de las enzimas determinantes en la actividad entomopatogénica, obteniéndose así porcentajes de mortalidad bajos y tiempos letales superiores a los alcanzados con los cultivos líquidos suplementados.
Los resultados de mortalidad obtenidos en las langostas, confirmaron la poca eficiencia patogénica del aislamiento Mt006 de M. anisopliae sobre R. schistocercoides. Dicho aislamiento mostró ser menos virulento que la cepa Mt004, a pesar de que ambas pertenecen a la misma especie de hongo entomopatógeno. Esta diferencia se podría atribuir a diferencias genéticas entre las cepas que estarían directamente relacionadas con la producción de enzimas y de toxinas. Dichas disimilitudes además, podrían ser determinadas por factores climáticos, estructurales, espaciales y genéticos de los insectos hospederos, factores que pueden producir cambios fisiológicos o genéticos de estos microorganismos, que incidirían en la reducción de la patogenicidad de los aislamientos (Lezama 1994).
Independientemente de la cepa utilizada o del tipo de medio de cultivo utilizado (líquido o sólido), los niveles mayores de control se produjeron en todos los casos por los conidios obtenidos en presencia de alas y patas de langosta y de salvado de trigo, lo que sugiere que estos suplementos actúan como inductores de la virulencia de M. anisopliae sobre R. schistocercoides.
Con base en los resultados de mortalidad, se seleccionaron para las pruebas enzimáticas los cultivos suplementados con homogeneizado de alas y patas de langosta y salvado de trigo por presentar los mayores niveles de control, también se seleccionaron el cultivo con quitina coloidal por presentar un comportamiento intermedio y los medios sólido y líquido sin suplementar, como tratamientos de comparación.
La actividad mayor de PR1, en extractos de conidios, se obtuvo cuando la cepa Mt004 de M. anisopliae se cultivó en medio líquido suplementado con alas y patas de langosta, siendo este resultado significativamente diferente de todos los demás. Esta actividad (Fig. 2) correspondió a 0.16 µmol de PNA por mg de conidios y fue seguida por la obtenida cuando el hongo se cultivó en presencia de salvado, siendo ésta de 0.13 µmol de PNA por mg de conidios. Estudios previos han demostrado que la inducción de enzimas sucede 24 horas después del contacto del hongo con el insecto, sugiriendo que la expresión de PR1 está directamente relacionada con la presencia de los componentes cuticulares de los insectos hospederos, los cuales le sirven de sustrato a dicha enzima (Clarkson y Charnley 1996). En los tratamientos control para las dos cepas se detectaron actividades enzimáticas de 0.088 µmol de PNA por mg de conidios y 0.080 µmol de PNA por mg de conidios para las cepas Mt004 y Mt006 respectivamente, valores que no fueron significativamente diferentes entre sí pero si lo fueron de los encontrados en los conidios provenientes de los medios líquidos suplementados con los potenciales inductores de virulencia. Este resultado podría indicar que los suplementos tuvieron un efecto inductor de la quimoelastasa proteasa PR1.
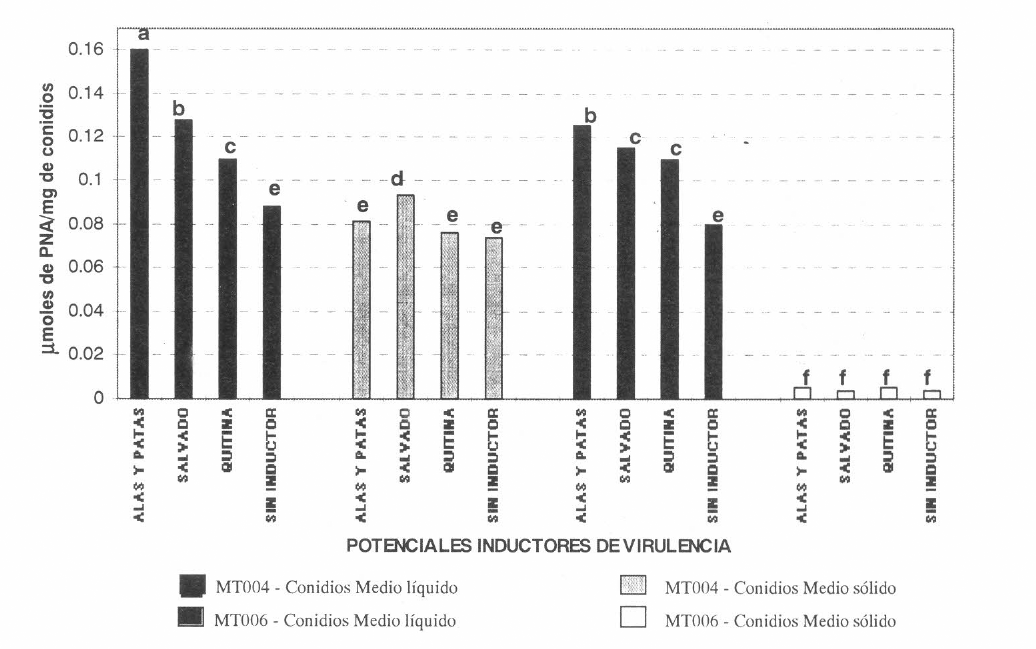
Efecto de los potenciales inductores de virulencia sobre actividad enzimática de la quimoelastasa proteasa PR1 en los extractos de conidios de M. anisopliae
Los conidios de la cepa Mt004 de M. anisopliae cultivada en medio líquido suplementado con alas y patas de langosta, fueron los más virulentos en los ensayos biológicos debido a que con éstos se alcanzó el 100% de mortalidad y se obtuvo el menor TL50. Este resultado fue seguido por el obtenido cuando el hongo se cultivó en presencia de salvado de trigo.
Estos resultados sugieren una relación entre la expresión de PR1 y la virulencia del hongo, la cual fue demostrada por Goettel et al. (1989), quienes transformaron genéticamente un aislamiento de M. anisopliae introduciéndole el gen benA3 de Aspergillus nidulans, para aumentar simultáneamente la virulencia del hongo clonado al inducir la actividad quimoelastasa proteasa PR1.
Independientemente de la cepa utilizada, se observaron diferencias marcadas entre las actividades enzimáticas detectadas para los extractos de los conidios provenientes de los cultivos sólidos y líquidos, siendo superiores las presentadas por estos últimos. Este fenómeno podría atribuirse a un efecto de represión de dicha enzima por efecto de una alta concentración de fuente de carbono en el medio sólido de arroz, sistema de regulación sugerido para PR1 (Clarkson y Charnley 1996).
Los conidios de la cepa Mt006 cultivados en medio sólido con todos los inductores presentaron niveles muy bajos de actividad PR1, los cuales fueron significativamente diferentes de los encontrados en los tratamientos restantes. Los niveles más bajos de actividad patogénica contra la plaga se obtuvieron con dichos conidios, sugiriendo que la enzima PR1 juega un papel determinante en el mecanismo de infección de M. anisopliae.
Al cultivar las dos cepas en medios sólido y líquido, se observaron diferencias significativas entre la actividad esterasa de los extractos de los conidios, siendo superior cuando se cultivó el hongo en medio líquido; esta diferencia podría deberse a un mecanismo de represión catabólica por exceso de fuente de carbono producida por el sustrato sólido utilizado, el cual está constituido por arroz que es rico en carbohidratos como el almidón (Clarkson y Charnley 1996).
La actividad esterasa en los extractos de conidios de ambas cepas de M.anisopliae crecidas en medio de cultivo líquido demostró ser significativamente mayor, para la cepa Mt004 cuando se cultivó en medio suplementado con salvado de trigo y para la cepa Mt006 cuando se cultivó en presencia de salvado de trigo y de alas y patas de langosta, siendo estos tratamientos significativamente diferentes de los demás, pero no significativamente diferentes entre sí (Fig. 3).
Efecto de los potenciales inductores de virulencia sobre actividad Nacetilglucosaminidasa detectada en los extractos de conidios de M. anisopliae
La actividad esterasa para la cepa Mt004 crecida en medio líquido suplementado con salvado de trigo fue de 860.47 ¡Lmol de PNA por mg de conldios, mientras que en presencia de alas y patas de langosta ésta fue de 821.13 ¡Lmol de PNApor mg de conidios y en presencia de quitina coloidal fue de 616.25 ¡Lmol de PNA por mg de conidios. Los tres tratamientos presentaron diferencias significativas entre sí, indicando que cada suplemento tiene diferente efecto inductor de la enzima, siendo el salvado de trigo el inductor más potente de la actividad esterasa.
Para la cepa Mt006 se determinó una actividad esterasa superior y estadísticamente diferente cuando se utilizaron los tres suplementos y cuando se cultivó el hongo en ausencia de los mismos, este resultado indica que para este aislamiento, todos los potenciales inductores de virulencia tienen un efecto inductor de dicha enzima.
Con los conidios producidos en los medios de cultivo líquido suplementados con alas y patas de langosta y con salvado de trigo, se obtuvieron los niveles mayores de control de la langosta lIanera con las dos cepas de H. anisopliae y a mayor actividad esterasa. sugiriendo que las esterasas juegan un papel importante en la virulencia de los conidios, debido a que los porcentajes mayores de mortalidad y los tiempos letales menores se obtuvieron con los conidios que presentaron los niveles mayores de esta enzima. Las esterasas son determinantes en las primeras etapas de la infección fúngica, debido a que son las encargadas de degradar los ésteres grasos que constituyen la capa cerosa que protege la cutícula del insecto hospedero. Cuando las esterasas hidrolizan estos ésteres, las enzimas que degradan la cutícula pueden actuar y permitir la penetración de la misma por el hongo entomopatógeno (Clarkson y Charnley 1996). Sin embargo, no se detectaron diferencias estadísticamente significativas entre la cantidad de enzima determinada para los conidios de la cepa Mt004 cultivada en presencia de alas y patas de langosta, los cuales fueron los conidios más virulentos contra la langosta y los conidios del aislamiento Mt006 cultivados con quitina coloidal, tratamiento que presentó un nivel de actividad controladora deficiente. Este resultado sugiere que a pesar de la relación encontrada para algunos casos entre la virulencia de los conidios y su actividad esterasa, éste no es el único factor determinante, sino que dicha virulencia podría estar influenciada por un conjunto de procesos físicos y químicos (Lezarna 1994).
Independientemente de la cepa utilizada y del tipo de medio de cultivo, se obtuvo la mayor actividad esterasa en los extractos de los conidios provenientes del cultivo suplementado con homogeneizado de alas y patas de langosta y con salvado de trigo, lo cual indica que estos sustratos podrían inducir una mayor síntesis de esta enzima o mejorar las características de permeabilidad de los conidios, permitiendo una mayor disponibilidad de la enzima (Thonart et al. 1991).
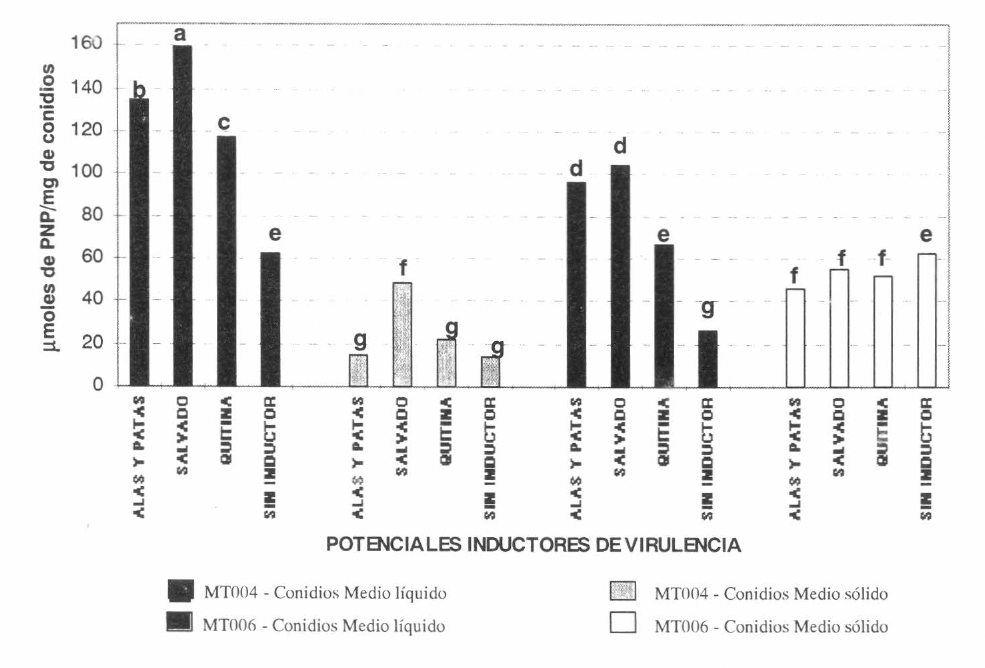
En cuanto a la actividad N-acetilglucosaminidasa en los extractos de los conidios de la cepa Mt004 provenientes del cultivo líquido, se detectaron actividades enzimáticas de 134. ¡Lmol de PNApor mg de conidios, 159.63 ¡Lmol de PNA por mg de conidios y 117.36 ¡Lmol de PNA por mg de conidios, cuando el medio de cultivo se suplementó con alas y patas de langosta, con salvado de trigo y con quitina coloidal respectivamente (Fig. 4). El tratamiento control presentó una actividad enzimática de 62.81 ¡Lmol de PNApor mg de conidios, la cual fue significativamente inferior a la actividad obtenida cuando se utilizaron los medios suplementados con los potenciales inductores de virulencia, dicha diferencia podría indicar que los suplementos tienen un efecto inductor sobre la actividad N-acetilglucosaminidasa. La mayor actividad enzimática fue encontrada en los extractos de los conidios cuando el medio se suplementó con alas y patas de langosta y con salvado de trigo, siendo la actividad enzimática detectada para estos tratamientos signlñcatlvamente diferente de la obtenida en los demás. Este resultado sugiere que estos compuestos inducen la actividad de la N-acetilglucosaminidasa, observándose una tendencia de mayor inducción cuando se utiliza salvado de trigo, el cual, ha demostrado en estudios previos la capacidad de inducir la síntesis y la actividad de enzimas hidrolíticas como celulasas, β-1,3-glucanasas y quitinasas en Trichoderma viridae (Thonart et al. 1991).
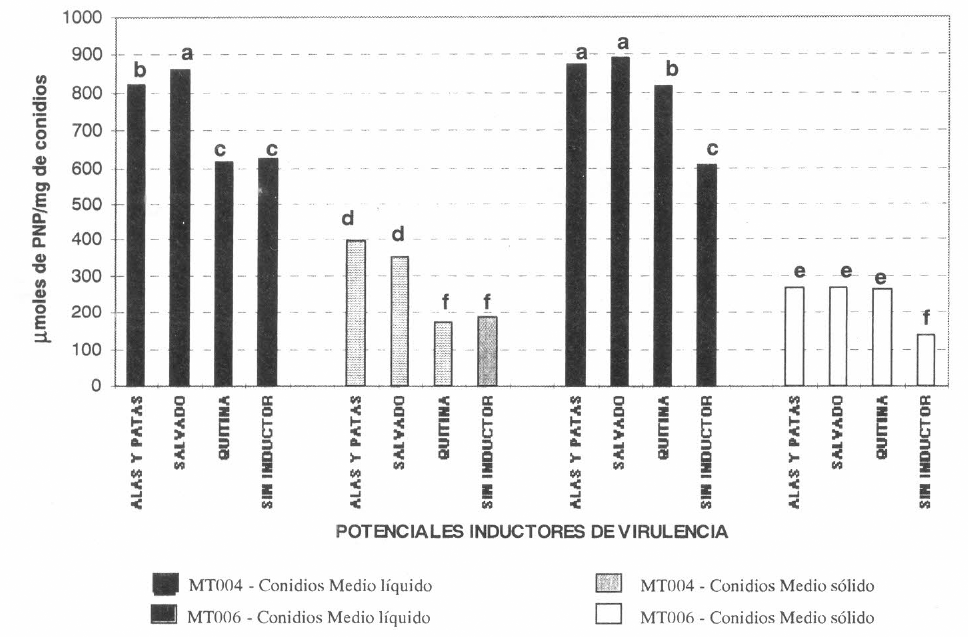
Efecto de los potenciales inductores de virulencia sobre la actividad esterasa en los extractos de conidios de M. anisopliae.
La actividad mayor N-acetilglucosaminidasa se detectó en los extractos de conidios de la cepa Mt004 provenientes de cultivo líquido en presencia de salvado de trigo, siendo este tratamiento significativamente diferente de todos los demás. Este resultado fue seguido por el encontrado para esta cepa cultivada en medio líquido suplementado con alas y patas de langosta, tratamiento que también fue significativamente diferente de los demás. Esta actividad enzimática mayor estuvo relacionada con los mayores niveles de control de la plaga en los bioensayos. La cepa Mt006 presentó la mayor actividad enzimática cuando creció en presencia de alas y patas de langosta y de salvado de trigo, presentando actividades equivalentes a 95.80 ¡Lmol de PNA por mg de conidios y 104 ¡Lmol de PNApor mg de conidios. El tratamiento control presentó una actividad significativamente inferior a la actividad obtenida con los conidios provenientes de los medios suplementados con los potenciales inductores de virulencia, confirmando el efecto de dichos sustratos sobre la actividad de dicha enzima. Al igual que con la cepa Mt004, fueron estos tratamientos los que produjeron los porcentajes de mortalidad mayores de la langosta llanera, Este resultado sugiere que dicha enzima podría estar relacionada con la virulencia de H. anisopliae sobre la langosta lIanera.
Los niveles menores de la enzima, para los extractos de los conidios de ambas cepas, se detectaron cuando el cultivo se hizo en medio sólido (arroz precocido). esto podría ser debido a la inhibición causada por la concentración alta de azúcares presentes en este sustrato. Thonart et al. (1991) señalaron una inhibición similar de las quitinasas para Trichoderma viridae cuando se utiliza glucosa como fuente de carbono.
En presencia de todos los potenciales inductores evaluados, se observó una mayor actividad de N-acetilglucosaminidasa en los extractos de los conidios de la cepa Mt004 cultivada en los medios líquidos con respecto a la actividad obtenida con los extractos de los conidios de la cepa Mt006 bajo las mismas condiciones, detectándose diferencias significativas entre las dos. Este resultado indica una posible relación entre la actividad de dicha enzima y las diferencias entre la actividad patogénica de estos dos aislamientos.
Al correlacionar los resultados de mortalidad acumulada corregida para las dos cepas de M. anisopliae con la cantidad de esterasa detectada en los extractos de los conidíos. se encontró un coeficiente de correlación de 0.55. Este valor es muy bajo, lo cual no permite usar dicha enzima como un marcador de virulencia. Al correlacionar los resultados de mortalidad acumulada corregida para las dos cepas de M. anisopliae con la cantidad de N-acetilglucosaminidasa detectada en los extractos de los conidios, se obtuvo un coeficiente de correlación de 0.44, valor que tampoco fue tenido en cuenta por ser muy bajo, mientras que el coeficiente de correlación logrado al aplicar regresión lineal a los datos de actividad enzimática de PR1 y porcentaje acumulado de mortalidad para las dos cepas fue de 0,82, valor que indica una correlación positiva entre la cantidad de dicha enzima presente en los conidios y la virulencia de las cepas sobre la langosta llanera.
Conclusiones
Se detectó una relación entre la composición del medio de cultivo y la virulencia del inóculo producido.
El efecto de los inductores potenciales de virulencia, sobre la actividad patogénica de M. anisopliae fue mayor cuando se cultivó en medio líquido que cuando se cultivó en medio sólido.
El homogeneizado de alas y patas de langosta y el salvado de trigo, fueron los mejores inductores de virulencia.
Los potenciales inductores de virulencia afectaron la actividad de las enzimas de M. anisopliae importantes en el mecanismo de infección.
Se determinó una correlación positiva entre la virulencia de los conidios de M. anisop/iae y la actividad de la quimoelastasa proteasa PR1.
Footnotes
Agradecimientos
Los autores del presente trabajo agradecen a Corpoica, ICA y Pronatta por su apoyo financiero.