
Abstract
In order to select entomopathogenous isolates against Premnotrypes vorax four isolates of Beauveria bassiana, three of Beauveria brongniartii and one of Metarhizium anisopliae, were evaluated under laboratory conditions. Adults of Premnotrypes vorax were treated with a unique concentration containing 1x108 propagules/ml, of each of them and mortality was evaluated daily. To determine their letal concentrations 50 (CL50) and 90 (CL90) successive concentrations of the selected isolates were used, from 1x105 to 1x109 progagules/ml. Biocontrol activity by combination of selected isolates of Metarhizium anisopliae and Beauveria bassiana was determined; the bioessay was performed by using the CL50 of each of them, and comparing the result with the mortality obtained by using twice CL50 of individual isolates. From the eight isolates evaluated Mt.L1 of Metarhizium anisopliae and Bv.L2 of Beauveria bassiana were selected because caused 100% insects mortality after 24 and 25 days respectively. These isolates showed letal concentrations 50 y 90 of 13x105 and 50x106 propagules/ml, respectively for Mt.L1 and 55x105 and 48x107 propagules/ml, respectively for Bv.L2. The mortality observed in Premnotrypes vorax was significantly superior when the isolates were utilized in combination because produced 100% of mortality ten days after the application, while when the individual activity was evaluated with the isolate Mt.L1 53,33% the mortality was observed and with the isolate Bv.L2 the mortality was of the 43,44%, in the same time.
Introducción
De las plagas que afectan al cultivo de la papa, el "gusano blanco", Premnotrypes vorax (Hustache) es uno de los más limitantes si se tiene en cuenta su amplia distribución en las zonas productoras, el costo que su control implica y el daño que ocasiona al tubérculo (López-Ávila 1996). Para su control se han utilizado métodos culturales, químicos y biológicos (Zenner 1986). El control cultural de la plaga y el manejo integral del cultivo, no han sido suficientes para disminuir los daños que causa, siendo muchas veces desplazado por el control químico, en el cual frecuentemente se hace un uso exagerado e inadecuado de plaguicidas. De esta manera se ha afectado severamente el medio ambiente al alterar las poblaciones de los enemigos naturales, fauna y flora asociadas; así como también al originar una serie de problemas relacionados con la contaminación ambiental del suelo, aire y agua e incluso de los mismos productos agrícolas.
El control biológico aparece como una alternativa promisoria para el control de esta plaga ya que pretende dar una solución a las debilidades de los demás métodos de control desde una perspectiva ecológica, haciendo uso de prácticas culturales y de enemigos naturales que han contribuido de manera representativa a mantener reguladas sus poblaciones (López-Ávila 1996).
Para el control biológico del gusano blanco se han utilizado los hongos entomopatógenos Beauveria bassiana, Beauveria brongniartii y Metarhizium anisopliae, los cuales se comportan como agentes efectivos de control al ser utilizados en forma individual (Torres et al. 1993). Sin embargo, teniendo en cuenta sus aspectos ecológicos similares y el hecho de poderlos encontrar en un mismo sitio geográfico, ejerciendo algún tipo de control sobre la plaga bajo la influencia de los mismos factores bióticos y abióticos, surgió la idea de evaluarlos mediante su utilización de manera combinada. Por lo tanto, la presente investigación tuvo los siguientes objetivos:
Aislar. identificar, y conservar cepas nativas de hongos entomopatógenos relacionadas con el control del gusano blanco.
Seleccionar. mediante bioensayos con adultos de la plaga, las cepas que presenten los mayores porcentajes de mortalidad acumulada corregida en el menor tiempo.
Determinar las concentraciones letales 50 (CL50) y 90 (CL90) de las cepas seleccionadas.
Comparar la actividad biocontroladora de las cepas seleccionadas sobre adultos de la plaga, al ser utilizadas de manera individual o combinada.
Materiales y Métodos
Con el fin de obtener el material biológico necesario para el desarrollo del trabajo, se definieron diferentes veredas pertenecientes a los municipios de Motavita (Boyacá) y Chocontá (Cundinamarca), determinando en éstas, sitios de almacenamiento y áreas cultivadas con una alta incidencia de la plaga.
En los sitios de almacenamiento se recolectaron insectos en diferentes estados de desarrollo con el propósito de buscar los que hubieran sido posiblemente afectados por algún hongo entomopatógeno, así como también muestras de suelo, para aislar cepas nativas promisorias para el control biológico de la plaga. Por otro lado, se seleccionaron áreas cultivadas con una alta incidencia de la plaga, con el propósito de obtener insectos en estado adulto para luego ser destinados al mantenimiento de la cría bajo condiciones de casa de malla, en el Centro de Investigación Corpoica - Tibaitatá. Esta cría permitió disponer del material necesario para llevar a cabo las pruebas de control biológico.
El aislamiento de los hongos entomopatógenos presentes sobre el cuerpo de los insectos recolectados, se llevó a cabo cultivando el hongo en medio Rosa de Bengala con antibióticos (Cloranfenicol 0.02 g y Streptomicina 0.02 g), aproximadamente 5 días a 23°C luego de haberlos mantenido por quince días en cámara húmeda a una temperatura aproximada de 20°C. Posteriormente, se hizo un repique en Agar Saboureaud Dextrosa (SDA). El aislamiento de los hongos presentes en las muestras de suelo se efectuó de la misma manera luego de hacer diluciones sucesivas para realizar la técnica de conteo en placa.
Paralelamente, todas las cepas encontradas se cultivaron en Agar Agua para observar las características microscópicas determinantes para su identificación. Posteriormente, se llevó a cabo su conservación en latencia, utilizando la técnica de conservación en suelo, para ser mantenidas en el Banco de Cepas del Programa Nacional de Manejo de Plagas (MIP) de Corpoica - Tibaitatá.
Para llevar a cabo las pruebas de patogenicidad se utilizaron cuatro cepas de Beauveria bassiana (Bv.L1, Bv.L2, Bv.L3 y Bv.L4), tres de Beauveria brongniartii (Bv.L5, Bv.L6 y Bv.L7) y una de Metarhizium anisopliae (Mt.L1), las cuales fueron evaluadas sobre adultos de Premnotrypes vorax procedentes de la cría mantenida bajo condiciones de casa de malla. Los insectos previamente desinfectados en una solución de hipoclorito de sodio al 0,5% se sumergieron durante un minuto y medio (1' 30") en una suspensión de cada una de las cepas nativas ajustadas a la concentración de 108 propágulos/ml contenidas en 25 ml de Tween 80 al 0,1%. Posteriormente, los adultos se montaron en cajas de Petri estériles que tenían en la base una toalla de papel húmeda y hojas de Solanum tuberosum para su alimentación. Estos materiales se renovaron diariamente para evitar posibles contaminaciones. Las cajas se mantuvieron bajo condiciones de laboratorio a una temperatura ambiente promedio de 18 °C.
Una vez hecha la inoculación de los adultos se hicieron observaciones diarias hasta obtener una mortalidad completa de insectos, llevando un registro de ésta, por réplica, para cada cepa. Los adultos se montaron en cámara húmeda para poder corroborar que su muerte fuera ocasionada por la cepa en estudio. Los datos obtenidos permitieron hacer una selección de las mejores cepas, teniendo en cuenta la mayor mortalidad acumulada corregida producida en el menor tiempo. con base al testigo tratado, siguiendo la fórmula de Abbott. También se tuvieron en cuenta dos testigos, un testigo absoluto (Ta) y un testigo tratado (Tt) en el cual los insectos se suspendieron en 50 ml de Tween 80 al 0,1%. Para llevar a cabo este procedimiento, se emplearon un total de 40 adultos por tratamiento, 10 para cada réplica.
Posteriormente, se llevó a cabo la determinación de las concentraciones letales 50 (CL50) y 90 (CL90) empleando una cepa nativa de Beauveria bassiana (Bv.L2) y otra de Metarhizium anisopliae (Mt.LI) las cuales fueron seleccionadas por sus altos efectos biocontroladores en un menor tiempo respecto a las cepas restantes. De cada una de las dos cepas se hicieron diluciones a las concentraciones seriadas desde 105 hasta 109 propágulos/ml. La infección de los adultos, su montaje, las observaciones y toma de datos diarios de mortalidad se llevaron a cabo del mismo modo descrito anteriormente. Luego de la muerte de los insectos y de su posterior montaje en cámara húmeda, se llevó a cabo un registro diario del crecimiento micelial sobre el cuerpo del insecto para cada cepa y en cada concentración. Para esto, se manejaron las categorías de uno (1), la cual correspondió a un porcentaje de cubrimiento entre el 1% y 25% del cuerpo del insecto, la dos (2) a un porcentaje de cubrimiento entre el 26% y el 50%, la tres (3) a un porcentaje de cubrimiento entre el 51% y el 75% y la cuatro (4) a un porcentaje de cubrimiento entre el 76 % y el 100%. Este registro sirvió para determinar si el cubrimiento micelial de las dos cepas estaba relacionado con la cantidad de inóculo aplicado y con los porcentajes de mortalidad obtenidos para cada tratamiento.
Para llevar a cabo la determinación de las concentraciones letales 50 (CL50) y 90 (CL90) de las cepas seleccionadas, se evaluaron 12 tratamientos y tres réplicas, cada una de ellas con 10 adultos. Los datos obtenidos fueron analizados por el método Probit.
Adicionalmente, se evaluó la actividad biocontroladora de las dos cepas seleccionadas al ser utilizadas de manera combinada, ajustando cada una de ellas a su respectiva concentración letal 50 (CL50), la cual fue de 55 x 105 propágulos/ml para la cepa Bv.L2 de B. bassiana y de 13 x 105 propágulos/ml para la cepa Mt.L1 de M. anisopliae. Posteriormente, las dos cepas se mezclaron en una vaso de precipitado en el cual se sumergieron los adultos previamente desinfectados. También se emplearon controles de comparación, para poder realizar posteriormente el análisis de la actividad biocontroladora de cada cepa de manera individual y combinada. Para ello, cada cepa se ajustó al doble de su respectiva concentración letal 50 (CL50), es decir, la cepa Bv.L2 se ajustó a una concentración de 11 x 106 propágulos/ml y la cepa Mt.L1 a una concentración de 26 x 105 propágulos/ml. En este procedimiento los testigos utilizados, el número de insectos, número de réplicas por tratamiento y el montaje de los ensayos se realizó de la misma manera descrita anteriormente.
Se hicieron anotaciones de la mortalidad diaria de los insectos tratados durante 25 días. Luego de montar los adultos muertos en cámara húmeda, se anotó el momento en que se observó el inicio del crecimiento micelial de cada cepa sobre el cuerpo del insecto. De igual manera, se manejaron las categorías tenidas en cuenta en la determinación de las concentraciones letales 50 (CL50) y 90 (CL90), registrando, de ser posible para cada una de ellas, el porcentaje de cubrimiento correspondiente a cada cepa.
Para todos los casos se utilizó un diseño experimental completamente al azar y los datos de mortalidad obtenidos, fueron analizados mediante un análisis de varianza y una prueba de comparación múltiple Duncan (p = 0.05).
Resultados y Discusión
Se definieron para su estudio diez fincas pertenecientes a cinco veredas, dos de ellas ubicadas en el municipio de Motavita (Boyacá) y las restantes en el muncipio de Chocontá (Cundinamarca). En estas fincas se localizaron los sitios de almacenamiento de papa, los cuales se caracterizaron por poseer un piso de tierra y por presentar tubérculos sanos y afectados por larvas del "gusano blanco" (aproximadamente el 50 % o más). Estos sitios fueron de dos tipos, uno cubierto y otro descubierto. El primero, se caracterizó por ubicarse al lado de la casa del agricultor, en éste los tubérculos presentes estaban protegidos por un techo, mientras que los sitios de almacenamiento descubiertos se caracterizaron por ubicarse lejos de las viviendas, en campo abierto o en parcelas destinadas para diferentes cultivos. La mayoría de los tubérculos allí presentes se encontraban desprotegidos y sólo pocos se vieron envueltos por lonas plásticas o por costales, con el fin de brindarles cierta protección frente a las condiciones ambientales.
Por otra parte, los sitios en los que se pudo realizar en forma óptima la recolección de adultos sanos de gusano blanco, para llevar a cabo las pruebas de patogenicidad, se caracterizaron por ser cultivos cuyas plantas se encontraron entre la germinación y el aporque. Lo anterior pudo deberse a que este periodo constituye la época del cultivo más vulnerable a la plaga. Según Calvache y Alvarado (1980), los huevos depositados en este período eclosionaran después de 60 días de edad del cultivo, o sea cuando ya se ha iniciado el periodo de tuberización y las larvas encuentran el alimento y el medio ideal para su desarrollo. Adicionalmente, estos mismo autores, establecieron que la población de adultos de Premnotrypes vorax se incrementa en un cultivo de papa a partir de la germinación. A medida que la planta se desarrolla, ésta se convierte en una excelente fuente de alimento y de protección, por lo cual, poco a poco los insectos se van congregando alrededor de la planta.
Se encontró que las toyas o plantas que surgen a partir de tubérculos abandonados en el sitio de cultivo procedentes de cosechas anteriores y de las cuales no se esperan cosechas futuras, también constituyen un sitio óptimo para la recolección de adultos de la plaga. Estos sitios se caracterizaron por tener suelos sin ninguna preparación, lo cual favorece temporalmente la ubicación de adultos debajo de terrones esperando la emergencia de las plantas, pudiendo permanecer en el lote siempre y cuando existan en su superficie tubérculos de la cosecha anterior o plantas hospederas de hoja ancha que le sirvan de alimento y de resguardo.
De cada uno de los sitios de almacenamiento, cinco cubiertos y cinco descubiertos, en lo posible se tomaron diez insectos de cada ínstar (Tabla 1). Bajo condiciones de almacenamiento descubierto situados en las fincas San Vicente, Las Quebradas, La Aurora, El Porvenir y Tres esquinas, se obtuvieron insectos sólo en estado larval. Las 38 larvas allí recolectadas, se caracterizaron por estar muertas, por tener coloración negruzca y por no presentar ningún indicio de crecimiento micelial. Al ser llevadas estas larvas al laboratorio, no se observó desarrollo de ningún hongo entomopatógeno. Las causas de la muerte de los insectos podrían deberse a diferentes factores abióticos o físicos, incluyendo el clima, la luz, las condiciones del suelo y la exposición solar que influyen en las densidades poblacionales de los insectos de manera importante (De Bach 1987). Por otra parte, la coloración negruzca de las larvas podría deberse según Amaya y Bustamante (1975) al estrés al cual fueron sometidas las larvas, lo que hace que las bacterias presentes en su intestino invadan el hemocele donde se multiplican en grandes cantidades causándoles la muerte por septicemia. En estas condiciones, los tubérculos se encontraron en estado de enverdecimiento y deshidratación, lo cual pudo haber ocasionado estrés en las larvas, al no satisfacer las necesidades nutricionales requeridas para culminar satisfactoriamente su ciclo biológico.
Lugares de colección de las cepas nativas de hongos entomopatógenos
En contraste, bajo las condiciones de almacenamiento cubierto, presentes en las fincas El Santuario, Primavera, San Vicente, Buenavista y El Cerezo, los tubérculos mostraron buenas condiciones fisiológicas, por lo cual fue posible encontrar en el suelo insectos en diferentes estados de desarrollo: Larvas en último Instar, pupas y adultos en período de melanización. En estas condiciones de almacenamiento, los tubérculos presentes se vieron protegidos frente a la acción de los rayos ultravioleta del sol y frente a las demás condiciones medioambientales adversas. Es así como estos tubérculos constituyeron una buena fuente de alimento para las larvas. De esta manera, las larvas en el momento de abandonar los tubérculos para iniciar su desarrollo pupal, encontraron buenas condiciones nutricionales y adecuadas condiciones de temperatura, humedad y oscuridad para llevar a cabo su ciclo biológico.
Una vez analizados estos insectos bajo condiciones de laboratorio, se observó el crecimiento micelial de diferentes especies de hongos. En estos sitios de almacenamiento cubierto, los microorganismos encontrados correspondieron a los hongos entomopatógenos Beauveria y Metarhizium. El primero creció abundantemente sobre los diferentes estados del insecto, así como también sobre una muestra de suelo; mientras que el segundo creció abundantemente sobre el cuerpo de una sola larva recolectada.
En contraste, en los sitios de almacenamiento descubiertos, los hongos encontrados correspondieron principalmente a contaminantes pertenecientes a los géneros Aspergillus y Penicillium, siendo este último inhibidor de Beauveria bassiana según estudios realizados por Lingg y Donaldson (1981). En ninguna de las muestras se encontraron hongos entomopatógenos.
Las cepas de entomopatógenos identificadas correspondieron cuatro a B. bassiana, tres a Beauveria brongniartii y una a M. anisopliae. Todas las cepas fueron aisladas a partir de insectos colectados en sitios de almacenamiento cubierto y en casa de malla, con excepción de una de Beauveria bassiana que fue aislada a partir de una muestra de suelo (Tabla 2). La diferencia relacionada con la obtención de los hongos entomopatógenos bajo las condiciones de almacenamiento cubierto, con respecto al aislamiento negativo de éstos bajo condiciones de almacenamiento descubierto, podría estar relacionada con el hecho de que las condiciones climáticas favorables a los tubérculos y a los hongos, ayudan a determinar en gran medida las densidades poblacionales de estos últimos y de sus hospederos. Cuando el clima contribuye a la supervivencia del huésped, la densidad poblacional de su patógeno aumentará, y para eliminar los excesos de hospederos producidos se necesitará de más inóculo microbiano (De Bach 1987).
Cepas nativas de Beauveria spp. y Metarhizium anisopliae aisladas a partir de insectos y de una muestra de suelo provenientes de los municipios de Motavita, Chocontá y Mosquera
Por otro lado, la escasa obtención de entomopatógenos a partir de muestras de suelo podría explicarse según estudios realizados por Doberski y Tribe (1980), porque en el suelo ocurren procesos de fungistasis, estando por lo tanto, la mayoría del inóculo de los hongos entomopatógenos restringido a cadáveres de insectos hospederos, donde la esporulación puede ser profusa. Argumento que concuerda con lo encontrado por López et al. (1995) en su estudio de persistencia de B. bassiana en el suelo, en donde estableció que puede existir un aumento del inóculo del hongo en el curso de microciclos, gracias a la presión ejercida por restos de insectos presentes.
Cuando las cepas potencialmente biocontroladoras fueron evaluadas de manera individual sobre adultos de P. vorax a una única concentración de 1 x 108 propágulos/ml, se encontró que los insectos tratados con las cepas Bv.L1, Bv.L2 y M1.L1 empezaron a presentar los primeros síntomas de la enfermedad a las 72 horas (tres días) de haberse efectuado la inoculación de los adultos. En contraste, con las cepas restantes los síntomas fueron visibles al cabo de las 120 horas (cinco días). Según Alves (1986), la germinación de los conidios de B. bassiana ocurre 12 horas después de la inoculación. Posteriormente en las siguientes 12 horas, el hongo penetra a través del integumento por acción mecánica y por efectos enzimáticos. Transcurridas 72 horas de la inoculación, el insecto está totalmente colonizado. Sin embargo, cuando los insectos mueren rápidamente después de la infección, a menudo no muestran evidencia de una infección fúngica extensiva. En este caso, la muerte se relaciona con la producción de enzímas extracelulares o metabolitos secundarios, compuestos que a su vez contribuyen a la patogenicidad del hongo naron con una menor actividad locomotora, con una disminución en el consumo de follaje, así como también con una disminución en la rapidez de su respuesta al calor. Observaciones que también fueron realizadas por Torres (1996) en su estudio del control microbiano del gusano blanco con Beauveria spp. y Metarhizium sp.
A lo largo del experimento (40 días) se observaron diferencias en la pato-genicidad de las ocho cepas evaluadas (Fig. 1). Las cepas Bv.L1, Bv.L2 y Mt.LI causaron en este tiempo los mayores porcentajes de mortalidad. Las cepas Bv.Ll y Bv.L2 de B. bassiana presentaron durante los primeros 21 días un 85.29% de mortalidad y la cepa Mt.L1 de M. anisopliae produjo un 91.17% en este mismo tiempo. A pesar de que estos resultados no presentaron diferencias significativas según el análisis de varianza (f=1,28; p=3,15), las cepas Bv.L2 y Mt.L1 produjeron al final del experimento los mayores porcentajes de mortalidad acumulada corregida, alcanzando el 100% hacia los 25 días para la cepa Bv.L2 y hacia los 24 días para Mt.L1, mientras que la cepa
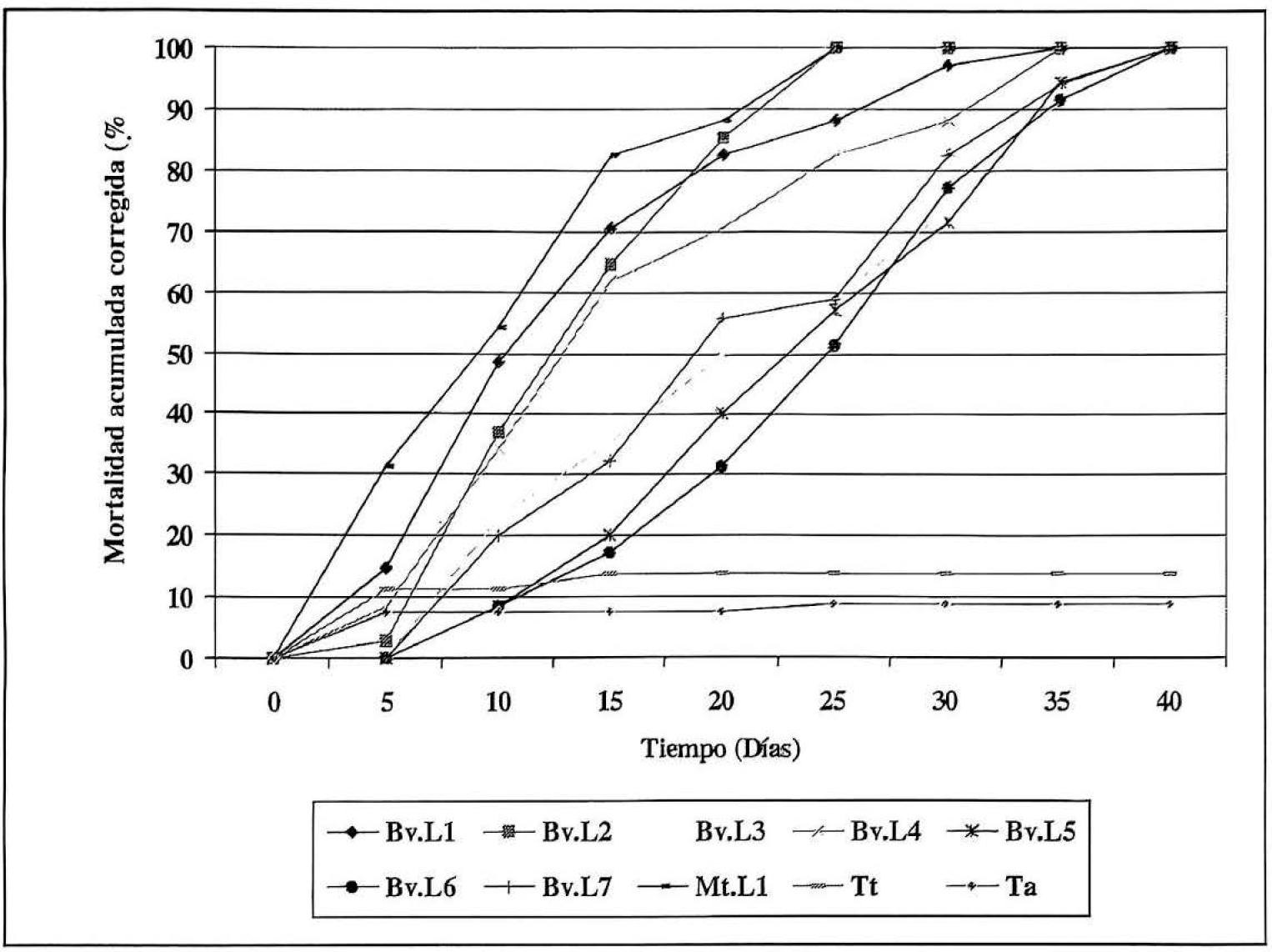
Efecto biocontrolador de las ocho cepas nativas aisladas al ser aplicadas a una única concentración de 1 x 108 propágulos/ml sobre adultos de Premnotrypes vorax
Bv.L1 presentó el 100% de mortalidad 31 días después de haber iniciado el ensayo. Debido a la rapidez en la acción de las cepas Bv.L2 y Mt.L1 éstas fueron seleccionadas para los posteriores ensayos.
Adicionalmente, se notó una deficiencia en la actividad biocontroladora por parte de las cepas Bv.L5, Bv.L6 y Bv.L7, las cuales correspondieron a Beauveria brongniartii. Estas cepas fueron las que tardaron más tiempo en causar la muerte de los insectos tratados ya que alcanzaron el 100% de la mortalidad entre los días 38 y 40.
Las diferencias encontradas en el comportamiento biocontrolador de las cepas evaluadas, podrían estar relacionadas con sus características genéticas y fisiológicas propias de éstas, ya que existen diferentes razas de una especie de hongo entomopatógeno que pueden presentar diferencias en su patogenicidad hacia un insecto hospedero determinado. Esto que ha sido aceptado como un hecho biológico dentro de la investigación moderna con estos organismos (Gillespie 1988), ha sido demostrado en cepas de B. bassiana las cuales han presentado diferencias en su virulencia contra la broca del café. De acuerdo a estudios realizados por Jiménez (1992), al evaluar bajo condiciones de laboratorio 46 aislamientos de este hongo provenientes de diferentes hospederos y de diferentes sitios de origen, se encontró que sólo cinco de ellos alcanzaron niveles de infección superiores al 80%, demostrando de esta manera los diferentes niveles de afinidad que pueden ocurrir entre un entomopatógeno y un hospedero determinado. Esta diferencia en la patogenicidad de las cepas, podría depender de un gran número de componentes tanto del patógeno como del hospedero tales como: germinación rápida, velocidad de reproducción, tasa de esporulación, tiempo de exposición, producción de toxinas y estado fisiológico del hospedero (Khachatourians 1991).
A pesar de las diferencias de patogenicidad de las ocho cepas evaluadas, todas a excepción de las cepas Bv.L5 y Bv.L6, presentaron un excelente crecimiento micelial, así como también una buena esporulación sobre el cuerpo de los insectos tratados.
A lo largo del experimento no se observaron diferencias significativas en los porcentajes de mortalidad acumulada correspondientes a los testigos: absoluto (Ta) y tratado (Tt), los cuales presentaron porcentajes de mortalidad de 8.75% y 13.75% respectivamente, al final del tratamiento. Estos bajos porcentajes de mortalidad permitieron hacer la evaluación de los tratamientos y de los signos de la enfermedad en los insectos ocasionada por cada uno de ellos.
Por otra parte, la determinación de las concentraciones letales 50 (CL50) y 90 (CL90) de las cepas Mt.L1 y Bv.L2, se llevó a cabo ajustando cada una de ellas a concentraciones sucesivas que iban desde 1 x 105 hasta 1 x 109 propágulos/ml. La toma de datos de mortalidad diaria y sus análisis se hicieron hasta el día 12, en el cual se alcanzó el 100% de mortalidad en los insectos con el tratamiento correspondiente a la cepa Mt.L1 evaluada a la concentración de 1 x 109 propágulos/ml. Se observó que a mayor concentración aplicada a los insectos, menor fue el tiempo en el que se presentó su mortalidad y mayor fue el porcentaje de mortalidad obtenido.
Al someter a un análisis de varianza los datos de mortalidad acumulada obtenidos para cada uno de los tratamientos en los ensayos de concentraciones letales, se encontraron diferencias significativas entre la mortalidad de los adultos (f=1,28; p=3,15). Es así, que cuando se aplicó B. bassiana a una concentración de 1 x 109 propágulos/ml se obtuvo el 100% de la mortalidad hacia el día 13 (Fig. 2). En contraste, con los tratamientos restantes se obtuvieron porcentajes de mortalidad por debajo del 50% con excepción del correspondiente a la cepa evaluada a la concentración de 1 x 108 propágulos/ml que produjo una mortalidad de 69.22%. Tales porcentajes fueron del 0%, 46.15% y 50% respectivamente para las concentraciones 1 x 105, 1 x 106 y 1 x 107 propágulos/ml.
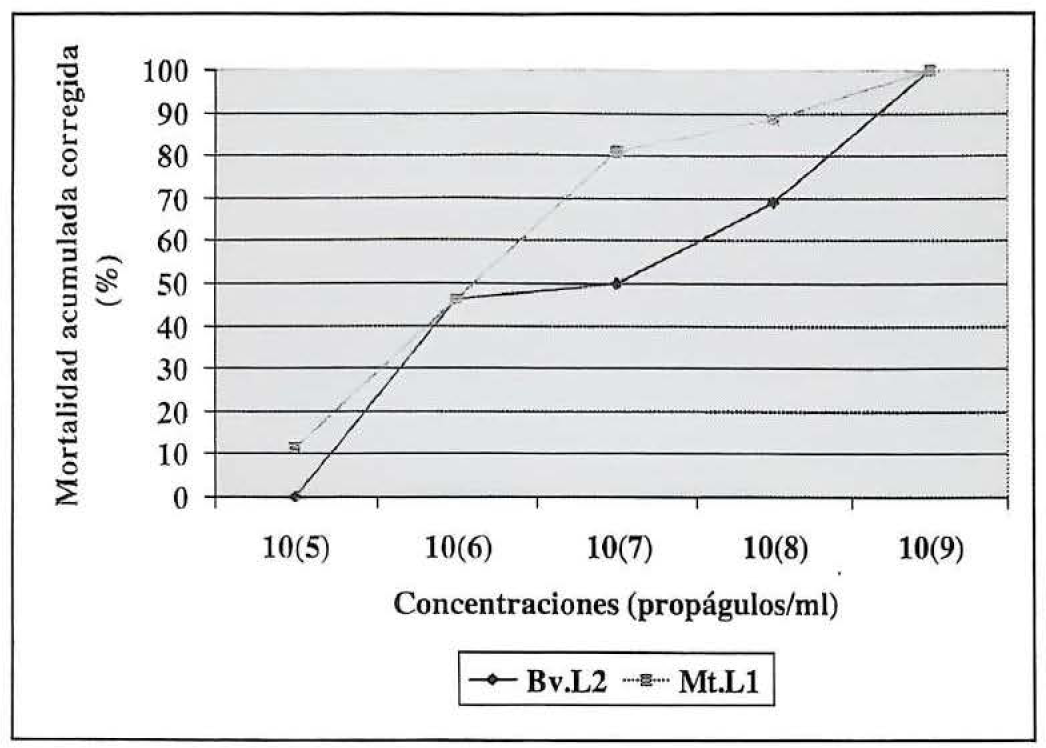
Efecto biocontrolador de diferentes concentraciones de os entomopatógenos seleccionados, sobre adultos de Premnotrypes vorax, determinado a los 12 y 13 días después de haber sido inoculados con Mt.L1 y Bv.L2, respectivamente
Cuando se aplicó M. anisopliae a la concentración de 1 x 109 propágulos/ml se obtuvo el 100% de mortalidad en los adultos de Premnotrypes vorax tratados en el día 12 (Fig. 2). Sin embargo, con los tratamientos correspondientes a las concentraciones de 1 x 107 y 1 x 108 propágulos/ml se alcanzaron en este tiempo porcentajes superiores al 50%, siendo éstos del 80%,76% y 88,46%, respectivamente. En los tratamientos restantes, se encontraron porcentajes de mortalidad de 11.53% y 46.15% en los insectos respectivamente para las concentraciones de 1 x 105 y 1 x 106 propágulos/ml de M. anisopliae. Se notó que la infección y posterior muerte de los adultos tratados, aumentó en proporción a la cantidad de propágulos infectivos del inóculo aplicado. Explicación que concuerda con los estudios realizados por Storey y Gardner (1988).
Al aplicar una prueba múltiple de Duncan se encontró que cuando las cepas Bv.L2 y Mt.L1 se evaluaron en forma individual a las concentraciones de 1 x 107 propágulos/ml y de 1 x 106 propágulos/ml, respectivamente, no presentaron diferencias significativas en sus porcentajes de mortalidad correspondientes al día 12, luego de la inoculación de los insectos (Tabla 3). De igual manera sucedió cuando se comparó el porcentaje de mortalidad correspondiente a la cepa Bv.L2 evaluada a una concentración de 1 x 105 propágulos/ml, respecto al porcentaje de mortalidad obtenido en los insectos del testigo tratado, para este mismo tiempo. En contraste, entre los tratamientos restantes si se presentaron diferencias significativas.
Prueba de Duncan aplicada en la determinación de las concentraciones letales 50 (CL50) y 90 (CL90) de las cepas Bv.L2 y Mt.L1
Promedios seguidos por la misma letra no presentaron diferencias significativas (p=0.05)
Al analizar los resultados obtenidos por el método estadístico Probit con 0.09 grados de significancia no se encontró una diferencia notoria entre las concentraciones letales 50 (CL50) correspondientes a cada cepa. Para la cepa Bv.L2 dicha concentración fue de 55 x 105 propágulos/ml y para la cepa Mt.L1 fue de 13 x 105 propágulos/ml. En contraste, si se pudo observar una diferencia notoria entre las concentraciones letales 90 (CL90), ya que para la cepa Bv.L2 ésta fue de 46 x 107 propágulos/ml, y para la cepa Mt.L1 fue de 50 x 106 propágulos/ml. De esta manera se pudo determinar que para obtener porcentajes satisfactorios de control, se requiere aplicar a los insectos una mayor cantidad de inóculo de la cepa Bv.L2, que de la cepa Mt.L1.
Los síntomas de la enfermedad presentados en los insectos sometidos a los diferentes tratamientos se observaron de manera notoria en la mayoría de aquellos que fueron tratados con altas concentraciones de los hongos entomopatógenos: 1 x 107, 1 x 108 y 1 x 109 propágulos/ml. Sin embargo, los síntomas fueron más severos en los adultos infectados con la cepa Mt.L1.
Cuando los insectos se inocularon con la cepa Bv.L2 de B. bassiana se encontró que el inicio del crecimiento micelial ocurrió de manera rápida, ya que cuando fue evaluado a una concentración de 1 x 109 propágulos/ml, éste se observó a los ocho días de inoculación alcanzando la categoría 1, y aumentó progresivamente hasta llegar a la categoría 4 en el día 23 (Tabla 4). En contraste con estos resultados y a pesar de que los diferentes tratamientos correspondientes a la cepa de Mt.L1 de M. anisopliae, fueron los responsables de los mayores porcentajes de mortalidad acumulada corregida, para el día ocho sólo fue posible observar el crecimiento micelial con una categoría 1 en los insectos muertos que habían sido sometidos al tratamiento que contenía 1 x 109 propágulos/ml.
Días correspondientes al inicio de cada categoría manejada en el registro de crecimiento micelial observada en los insectos tratados con diferentes concentraciones de los hongos entomopatógenos
De igual manera sucedió con el caso de la categoría 4. La cepa Bv.L2 alcanzó dicha categoría hacia el día 16 con el tratamiento Bv.L2 a una concentración de 109 propágulos/ml, mientras que la cepa Mt.LI la alcanzó sólo hasta el día 20 bajo la misma concentración. A pesar de que se observó una mayor rapidez en el cubrimiento micelial de las dos cepas evaluadas a la concentración de 1 x 109 propágulos/ml, con todos los tratamientos evaluados para la cepa Bv.L2 se obtuvo un porcentaje de cubrimiento hasta del 100%. En contraste, con los tratamientos de la cepa Mt.L1, el mayor cubrimiento micelial estuvo entre el 90% y 95% aproximadamente del cuerpo del insecto. De manera general se notó que a mayor concentración del hongo utilizada, más rápido fue visible el crecimiento de éste en el insecto, siendo el tiempo mayor para M. anisopliae que para B. bassiana.
En cuanto a la esporulación, ésta ocurrió en los insectos en todos los tratamientos con B. bassiana y M. anisopliae, presentándose para cada uno de ellos antes de que el crecimiento micelial representara aproximadamente el 50% del cuerpo del insecto. La manifestación de los hongos se inició en las suturas intersegmentales, patas y pico. Posteriormente, se observó la esporulación que se caracterizó por tener un aspecto polvoroso, típicamente de color blanco para la cepa Bv.L2 y verde para la cepa Mt.L1.
Finalmente, la combinación de las cepas se hizo ajustándolas a su concentración letal media, para la cepa Bv.L2 ésta fue de 55 x 105 propágulos/ml y para la cepa Mt.L1 ésta fue de 13 x 105 propágulos/ml. Adicionalmente, también se evaluaron dos controles que sirvieron como parámetro para establecer las diferencias entre la actividad biocontroladora de manera individual y combinada. Tales controles representaron el doble de la concentración letal 50 (CL50), correspondiente a cada cepa; es decir, la cepa Bv.L2 se ajustó a una concentración de 11 x 106 propágulos/ml y la cepa Mt.L1 a una concentración de 26 x 105 propágulos/ml.
Los porcentajes de mortalidad acumulada alcanzados hasta el día 10 por dichos tratamientos, incluyendo los obtenidos por los testigos tratado y absoluto, se sometieron a un análisis de varianza. Dicho análisis encontró diferencias significativas entre todos los tratamientos (f=43,62; p=2,58). Observación que fue corroborada al aplicar la prueba múltiple de Duncan (Tabla 5).
Prueba de Duncan correspondiente al promedio de mortalidad acumulada de adultos de Premnotrypes vorax causada por las cepas Bv.L2 y Mt.L1, al utilizarlas de manera individual y combinada
Promedios seguidos por la misma letra no presentaron diferencias significativas (p=0.05)
Al evaluar la actividad biocontroladora de las cepas Bv.L2 de B. bassiana y Mt.L1 de M. anisopliae de manera combinada, sobre adultos de P. vorax, se encontró un porcentaje de mortalidad acumulada corregida del 25% al día siguiente de inoculados. Porcentaje que aumentó rápidamente, hasta alcanzar un 51,84% y 100% de mortalidad hacia el cuarto y décimo día, respectivamente. En contraste, cuando se evaluaron las cepas Bv.L2 y Mt.L1 de manera individual, se observó al día siguiente de haber inoculado los insectos un porcentaje de mortalidad del 3.57% y 14,29%, respectivamente. Sin embargo, aunque dichos porcentajes aumentaron gradualmente, no fue posible alcanzar el 50% de la mortalidad al final del experimento (Fig. 3).
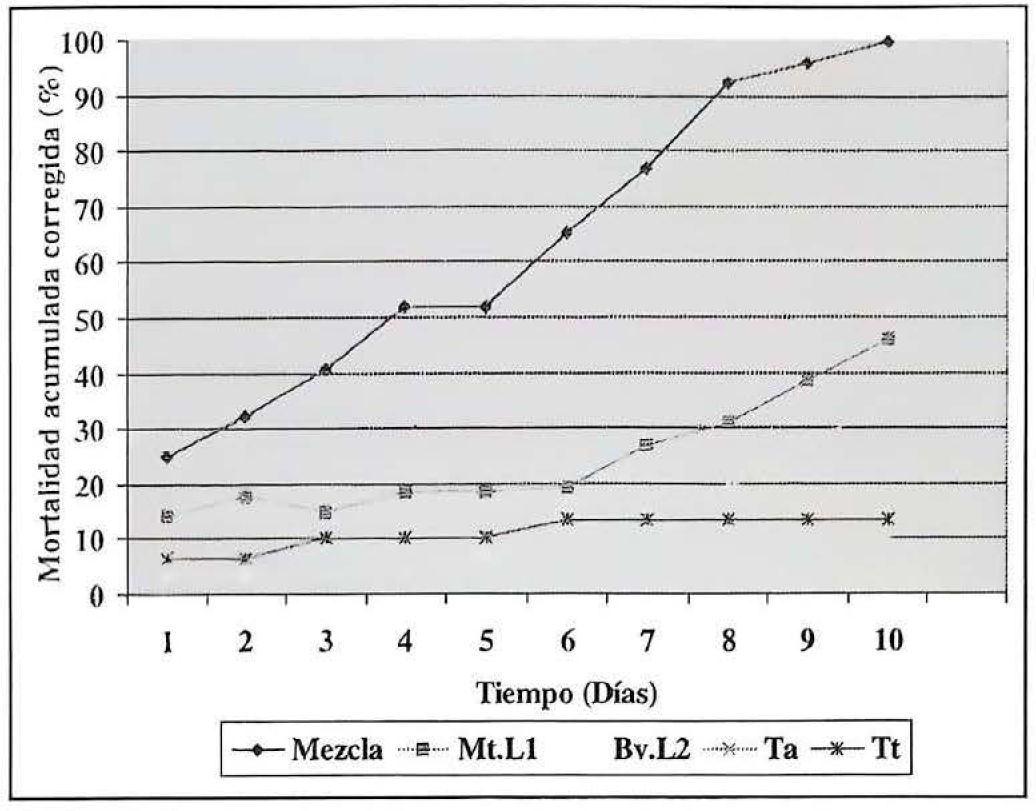
Porcentajes de mortalidad acumulada corregida obtenidos en la evaluación de la actividad biocontroladora combinada de las cepas Bv.L2 y Mt.L1 para el día 10.
La disminución, en el tiempo, de la mortalidad de los insectos tratados con la mezcla de cepas con respecto a su utilización en forma individual, podría relacionarse con un efecto sinérgico de mecanismos de acción correspondientes a cada cepa. Estos resultados podrían explicarse en parte por la acción combinada de las toxinas de B. bassiana y de M. anisopliae, a la que se sometieron los insectos.
En procesos de control biológico con Metahizium spp., normalmente se lleva a cabo una síntesis de diferentes péptidos cíclicos de variada composición amino e hidroxiácido, como es el caso de las destruxinas producidas por M. anisopliae. Toxinas que, según Proprawski et al. (1986), pueden variar en su acción sobre el hospedero. Tal es el caso de la Destruxin E, que puede actuar en el momento de la infección con un efecto tóxico que se caracteriza por producir en el insecto flacidez y parálisis del cuerpo. Adicionalmente, la cepa de B. bassiana podría haber sintetizado los compuestos denominados Beauvericin, Beauve-roliodes y Bassianolidae, los cuales están asociados con la actividad insecticida del hongo (Gillespie y Claydon 1989).
Este evento de sinergismo en la toxicidad se apoya en el hecho de haber obtenido al día siguiente de haber realizado la inoculación de los adultos con la mezcla de cepas un porcentaje de mortalidad acumulada corregida del 25%. Mientras que en este mismo día, al utilizarlas de forma individual, se obtuvieron porcentajes de mortalidad del 3,57% y 14,29% respectivamente para Bv.L2 y MI.L1. Teniendo en cuenta estos resultados el más rápido efecto de mortalidad podría atribuirse a la actividad tóxica de Mt.L1. Adicionalmente, la actividad de este hongo podría haber sensibilizado los insectos a otros mecanismos de acción correspondientes a B. bassiana o a M. anisopliae que involucran procesos mecánicos y enzimáticos. Tales mecanismos de acción se inician cuando el insecto es infectado por un hongo entomopatógeno ya que sus conidios germinan en la superficie del hospedero, producen un tubo de germinación corto en cuyo extremo forma un abultamiento llamado apresorio, luego de un crecimiento extensivo sobre los pliegues de la superficie del insecto. Posteriormente la hifa infecciosa penetra a través de la cutícula y lentamente penetra el hemocele del insecto (Khachatourians 1991).
Después del contacto entre el hongo entomopatógeno y el insecto se ofriletizan enzimas con efecto específico sobre cada uno de los componentes de la epicutícula y cutícula. La primera es desintegrada por la acción de las lipasas y la segunda por las quitinasas, N-acetil-ẞ- glucosa-minidasas y protcasas (Clakson y Charnley 1996). Dentro de estas enzimal. la serina proteasa de M. anisopliae presenta dos actividades, Prl y Pr2 responsables de la hidrólisis del 25% al 30% de las proteínas cuticulares, que con secretadas por el mismo apresorio y por la hifa que penetra la cutícula, así como también de la separación física de capas (Khachatourians 1991). Estas enzimas de M. anisopliae podrían haber causado rompimiento de la cutícula del insecto, facilitando así la penetración de B. bassiana.
Los resultados obtenidos en el presente trabajo demostraron que la actividad biocontroladora de las cepas Bv.L2 y Mt.L1 puede ser potencializada mediante su utilización de manera combinada presentándose cierta interacción entre ellas. Dicha interacción podría producirse mediante mutualismo o sintrofia. En el primer caso, los microorganismos podrían haber superado de alguna manera cualquier factor limitante en el medio, haciendo más fácil su desarrollo; mientras que en el caso de ocurrir sintrofia los dos organismos involucrados, podrían haber compartido el mismo microambiente, debido a que el producto del metabolismo de uno de ellos le sería fácilmente accesible al segundo (Campbell 1987).
Cuando los insectos se infectaron con la mezcla de cepas, al analizarlos, se encontró la presencia de micelio y de conidios pertenecientes a la cepa Bv.L2 de B. bassiana y en ningún caso se observó M. anisopliae. Tal observación se llevó a cabo desde la aparición micelial que tuvo lugar el día 4, hasta obtener un cubrimiento casi total el cual ocurrió en el día 12 (Tabla 6).
Días correspondientes al inicio de cada categoría manejada en el registro de crecimiento micelial observada en los insectos tratados con las cepas Bv.L2 y Mt.L1 utilizadas de forma individual o combinada
Esta situación podría ser explicada si se tiene en cuenta que la cepa Bv.L2 de B. bassiana, cuando fue aplicada en forma individual, siempre tardó menos tiempo en expresarse (siete días) y en cubrir el cuerpo de los insectos tratados, llegando siempre a 100% de cubrimiento. En contraste, la cepa Mt.L1 de M. anisopliae se expresó a partir de los 10 días y alcanzó en el día 24 su máximo porcentaje de cubrimiento, el cual nunca fue superior al 95%.
Según Campbell (1987), cada organismo interactúa con su entorno, modificando las características de éste en un proceso determinado. Tal es el caso de la competencia por las fuentes de nutrientes, energía y espacio escasos. De esta manera pudieron haber entrado en juego una serie de características básicamente relacionadas con unas tasas altas de crecimiento y reproducción, así como también con la capacidad de producir toxinas o enzimas. Estas características darían origen a ciertas ventajas competitivas, que para este caso se observaron entre la cepa Bv.L2 de B. bassiana y la cepa Mt.L1 de M. anisopliae.
El hecho que sólo B. bassiana se encontrara cubriendo el cuerpo de los insectos muertos, también podría explicarse por un posible fenómeno fungistático o fungicida que B. bassiana pudo haber ejercido contra M. anisopliae no permitiéndole su desarrollo, ya que según Poprawski et al. (1986), estos dos hongos entomopatógenos, tienen la capacidad de producir grandes cantidades de sustancias con dichas propiedades.
Conclusiones
Las condiciones climáticas favorables para los tubérculos y para Premnotrypes vorax, influencian la presencia de hongos entomopatógenos en los insectos.
Las cepas nativas de Beauveria bassiana y Metarhizium anisopliae evaluadas sobre adultos de la plaga presentaron una alta actividad biocontroladora.
La utilización combinada de las cepas Bv.L2 de Beauveria bassiana y Mt.L1 de Metarhizium anisopliae demostró una actividad biocontroladora significativamente superior a la producida por estos hongos al ser utilizados de manera individual sobre adultos de la plaga.