
Abstract
Bacillus thuringiensis (Berliner) is the most worldwide used entomopathogenic microorganism for the biological control of pest insect. In the same way, it is known as a very cosmopolite microbe, with the capacity to growth in a variety of ecosystems such as tropical rain forest or desert areas. Instead of the wide ecological distribution of the specie, most of the studies related with the isolation of B. thuringiensis (Bt), have been using soil samples from agricultural ecosystems, where it is found the pest insect we wish to control. In this study, we compare the presence of Bt in soil samples from the native forest of the National Natural Park Katios (PNN Katios) and different agricultural ecosystems (coffee, tobacco, grass, potato and others). In order to accomplish these aims we took seven soil samples from PNN Katios and 21 from the agricultural ecosystems which were analyzed by the methodology of Travers et al. (1987). Twenty-nine strains were obtained from 28 soil samples. Twenty-ones of these strains are from PNN Katios. The eighty three percent of the strains from the native rain forest had atypical patterns in terms of the inclusion bodies and the electrophoretic profile of the proteins of such bodies. On the other hand, from the agricultural soil samples it is important to remark the strain IBUN115A32 which presented crystals of three different shapes and the strain IBUN97B1 which presented, in the PCR reaction, a DNA band of different size which may suggest the presence of a new cry gene. The results obtained in this study confirm the importance of the screening of new Bt strains in order to know the real diversity of Bt.
Introducción
Bacillus thuringiensis (Berliner) es una bacteria Gram positiva que se caracteriza por su capacidad de producir inclusiones cristalinas paraesporales durante la fase de esporulación con formas que van desde la piramidal, que es la más frecuente, hasta la amorfa, pasando por triangular, cuadrado y rectangular. Este cristal está conformado básicamente por proteínas denominadas d-endotoxinas de diverso peso molecular (25 a 140 KDa) (Bernhard et al. 1997). Dicha toxina presenta actividad tóxica específica contra insectos plaga de diferentes órdenes tales como Lepidoptera, Homoptera, Coleoptera y Diptera además de otros invertebrados de interés en salud y/o en el sector agrícola donde cabe destacar platelmíntos, protozoos, ácaros y nemátodos (Feitelson y Kim 1992; Medrano y Quintero 1996).
Una de las principales líneas de investigación con B. thuringiensis (Bt) consiste en la búsqueda y caracterización de nuevos aislamientos que permitan ampliar el conocimiento en el espectro de acción de dicho agente entomopatógeno y/o encontrar aislamientos nativos que posean mayor actividad biopesticida que la señalada actualmente contra los organismos blanco de interés. Dentro de este contexto se han realizado diferentes trabajos tendientes a describir la presencia y diversidad de esta especie en un gran número de países a nivel mundial. Para lograr este objetivo se han diseñado diversas estrategias, teniendo como criterio la selección de las fuentes más frecuentemente utilizadas por este microorganismo como son suelo (Bernhard et al. 1997; Hossain et al. 1997; Ohba y Aratake. 1994; Chak et al. 1994; Vasquez et al. 1995; Chilcott y Wigley 1993); material vegetal (Bernhard et al. 1997; Smith y Gouche 1991); insectos (Bernhard et al. 1997; Apoloyo et al. 1995): hábitats de insectos (Bernhard et al. 1997; Chilcott y Wigley 1993) y productos o zonas de almacenamiento (Meadows et al. 1992; Burgues y Hursi 1977). La mayoría de estos estudios están dirigidos al aislamiento de cepas de un ecosistema específico y la determinación de una o varias de las características de interés para Bt como son la serotipificación de las cepas (Apoloyo et al. 1995; Vasquez et al. 1995), la forma del cristal paraesporal (Bernhard et al. 1997; Kaelin et al. 1994), determinación del peso molecular de las proteínas asociadas al cristal (Chilcott y Wigley 1993; Kaelin et al. 1994; Chak et al. 1994), la tipificación del tipo específico de gen cry (Cerón et al. 1994, 1995; Bourque et al. 1993; Carozzi et al. 1991) y la determinación de la actividad biopesticida contra diferentes especies blanco de acuerdo con las necesidades de cada país.
Pocos estudios a nivel mundial (Chilcott y Wigley 1993; Ohba y Aratake 1994; Martin y Travers 1989) han incluido dentro de su diseño experimental el análisis de la diversidad de Bt presente en los bosques nativos y menos en aquellos bosques presentes en países tropicales. Esta falta de interés en este ecosistema posiblemente se deba a la creencia de que el entomopatógeno que controle un insecto blanco de importancia económica, debe ser encontrado estrechamente asociado a éste, lo cual significaría que la mejor fuente del Bt activo contra un insecto de interés, es el ecosistema donde normalmente este habita, bien sea un cultivo agrícola o ciertas zonas pantanosas si se desea controlar insectos vectores de enfermedades tropicales. Sin embargo, muy poco se conoce en relación con las condiciones bióticas o abióticas que podrían estar favoreciendo la presencia de Bt en el ambiente. De hecho hoy en día aun no está muy claro el grado de dependencia de Bt con su insecto blanco, ya que mientras para algunos autores es evidente que el hábitat más adecuado para la proliferación del microorganismo en la naturaleza, es el cuerpo de un insecto (Luna y Peña 1996) otros demuestran que los mejores sitios como fuente de aislamientos de Bt no necesariamente son aquellos que presenten una alta actividad de insectos, sino por el contrario aquellos donde ni siquiera se detectan insectos fácilmente como son las zonas de desierto (Martin y Travers 1989). Varias colecciones de cepas de Bt han sido descritas, principalmente de Europa, Asia, Africa, Nueva Zelanda y los Estados Unidos (Hossain et al. 1997; Apoloyo et al. 1995; Chak et al. 1994; Bernhard et al. 1997; Chilcot y Wigley 1993; Martin y Travers 1989). Ninguna de estas colecciones ha incluido muestras de Latino América. Llenando este vacío Bravo et al. 1998 presentan la caracterización de la colección de cepas de Bt mexicanas en la cual señalan que el 49.5% de las cepas porta los genes cry1, con 33 diferentes tipos de perfiles, 21.5% de las cepas portan genes cry3 y el 7.9% de las cepas contienen genes cry11, cyt, resaltando la presencia de cepas que pueden portar potenciales nuevas proteínas Cry. Así mismo algunos grupos chilenos se han puesto en la tarea de estudiar colecciones de Bt, haciendo una caracterización general de los aislamientos encontrados en términos de la presencia de genes cry (Theodoluz et al. 1997) o de las proteínas asociadas al cristal (Vasquez et al. 1995), y relacionándolos con la actividad biopesticida contra una plaga de interés. Diaz et al. 1993 realizaron un muestreo en diferentes áreas de Colombia para encontrar cepas de Bt con actividad a larvas de mosquitos, haciendo la caracterización mediante pruebas bioquímicas de los aislamientos más activos. Ninguno de los estudios antes mencionados plantea dentro de sus objetivos establecer alguna relación entre los aislamientos obtenidos y el tipo de ecosistema del cual fueron extraídos.
Dentro de este trabajo, se pretende determinar la presencia de Bt en muestras de suelo provenientes de algunas zonas de bosque natural nativo del Parque Nacional Natural Katios y compararla con la de algunos ecosistemas agrícolas, con el ánimo de aportar más información respecto al tipo de hábitat preferido por Bt. El análisis de las cepas obtenidas en cada caso, consistió en la determinación de la forma del cristal o cuerpo paraesporal, el peso molecular de las proteínas asociadas al cristal, y la presencia de genes cry con actividad contra lepidópteros y coleópteros.
Materiales y Métodos
Recolección de muestras
Se recolectaron 28 muestras de suelo provenientes de zonas de bosques naturales y zonas agrícolas de diferentes partes del país (Antioquia, Cundinamarca, Huila y Meta) (Tabla 1). Para la recolección de las muestras se eliminó totalmente la vegetación que cubría el suelo de cada sitio, tomando con una pala limpia 0.5 kg de la porción superficial de suelo hasta una profundidad de 10 cm. Cada muestra se colocó en bolsas plásticas estériles y se llevó al laboratorio para ser secadas a temperatura ambiente, homogeneizadas mediante un rodillo y posteriormente tamizadas a través de un tamiz de 210 mm de poro, finalmente las muestras se procesaron para el aislamiento de Bt o se almacenaron a 4ºC hasta su utilización.
Cepas Control
Durante el desarrollo del estudio se emplearon cepas control como la HD1 Bt subesp. kurstaki la cual es portadora de genes cry1Aa, cry1Ab, cry1Ac, cry2Aa y cry2Ab. También se utilizaron las cepas con los genes cry3Aa, cry3Ba, cry3Bb, cry3Ca, todas suministradas por el banco de cepas y genes del Instituto de Biotecnología de la Universidad Nacional de Colombia.
Aislamiento de cepas nativas
Para este procedimiento se emplearon las muestras de suelo previamente procesadas siguiendo la metodología de Travers et al. (1987).
Electroforesis en geles de Poliacrilamida-SDS (SDS-PAGE)
El análisis de proteína fue hecho en geles al 12% de poliacrilamida dodecil sulfato de sodio (SDS-PAGE, de acuerdo con lo señalado por Laemmli (1970).
Análisis por PCR
Para este procedimiento las cepas de Bt se crecieron en placas de Agar Nutritivo siguiendo el procedimiento sugerido por Cerón et al. (1994; 1995). Una asada de células de una colonia se tranfirieron a 100 ul de agua y la mezcla se calentó a ebullición por 10 minutos para lisar las células. Se usaron 10 µl de esta solución como muestra de DNA para el proceso de PCR de acuerdo con lo señalado por Cerón et al. (1994: 1995). La amplificación se realizó usando un termociclador de DNA (Perkin-Elmer Mod. 480). Las condiciones para la amplificación se hicieron con primers generales para cry1 y primers generales para cry3, 7 y 8. El producto de amplificación se analizó en geles de agarosa al 3%, usando como marcador de peso molecular el plásmido pBR322 digerido con la enzima de restricción Hinf I.
Resultados y Discusión
Aislamiento de cepas nativas de Bacillus thuringiensis
Para el aislamiento de cepas nativas de Bt se tomaron muestras de suelo provenientes de 7 diferentes zonas de bosque nativo del Parque Nacional Natural Katios (PNN Katios) y 21 muestras provenientes de diversos agroecosistemas pertenecientes a los departamentos de Antioquia, Cundinamarca, Huila y Meta (Tabla 1). Se obtuvieron 29 cepas de Bt de las cuales el 82,7% procedían de zona de bosque Natural y el 17,4% restante de los ecosistemas agrícolas; es importante mencionar que el 71% de las muestras de bosque natural y únicamente el 19% de las muestras de ecosistemas agrícolas presentaron, al menos, una cepa de Bt (Tabla 1).
Este análisis claramente está evidenciando una mayor presencia de Bt en bosques naturales del PNN Katios con relación a los ecosistemas agrícolas evaluados. Dichos resultados concuerdan con el trabajo de Chilcott y Wigley (1993), donde se obtuvo un mayor porcentaje (79%), de muestras de suelo obtenidas de bosques nativos, con, al menos, una cepa de Bt en comparación con los suelos agrícolas u hortícolas cuyo porcentaje de muestras con Bt correspondió a 60% y 61%, respectivamente. Sin embargo otros trabajos, como el de Martin y Travers (1989), encuentran porcentajes de 66.7% y 87.1% para muestras de bosques de lluvia tropicales y suelos agrícolas, respectivamente, los cuales evidentemente son contradictorios con los datos mencionados anteriormente.
El análisis de los resultados sugiere que más que unas condiciones ecofisiográficas específicas, el éxito en el establecimiento de una población de Bt reside en unas condiciones microecólogicas determinadas posiblemente por el pH, nutrientes, humedad y lo que parece ser más importante para Bt, las presiones ejercidas por antagonistas microbianos, ya que Bt posee una pobre capacidad para establecerse plenamente dentro de una comunidad edáfica (Luna y Peña 1996).
De las cepas aisladas de las muestras de suelo, se seleccionaron las correspondientes a Bt por sus características macroscópicas y microscópicas. Microscópicamente corresponden a bacilos Gram positivos largos, que en su fase de esporulación producen cristales, los cuales adoptan diferente morfología, siendo éstos de forma bipiramidal, triangular, rectangular, cuadrada y amorfa. En este estudio es necesario resaltar que un porcentaje alto de aislamientos presentaron inclusiones paraesporales de tipo triangular, las cuales no son muy frecuentes dentro de la morfología típica para este tipo de inclusiones en Bt. La gran mayoría de los aislamientos con este tipo de cristales corresponden a diferentes muestras de suelo obtenidas del PNN Katios (Tabla 2). El perfil electroforético de las proteínas de las cepas aisladas presentaron bandas prominentes con pesos moleculares aproximados entre 25 y100 kDa (Fig. 1).
Es importante señalar que una cepa proveniente de cultivo de tabaco posee tres tipos diferentes de cristal: bipiramidal, cuadrado y rectangular (Fig. 2; Tabla 2). Esta característica la presentan muy pocas cepas por lo que datos semejantes son poco registrados en la literatura. El perfil electroforético de las proteínas asociadas al cristal de esta cepa mostró dos bandas de pesos moleculares entre 130-140 kDa y 60-70 kDa (Fig. 3). El encontrar cepas con características poco comunes en suelos colombianos muestra la gran diversidad de Bt, ya que tanto la cepa aislada de cultivo de tabaco, como las del PNN Katios se alejan de las características actualmente registradas en la literatura para la especie.
Distribución de Bacillus thuringiensis en las muestras de suelo analizadas
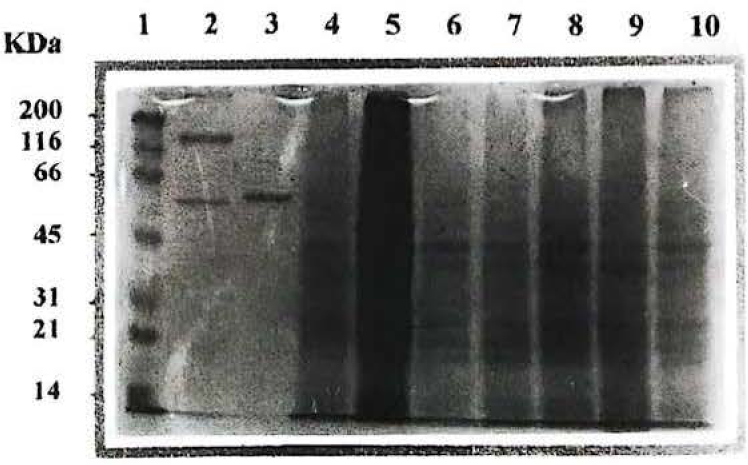
Perfil electroforético de las proteínas asociadas al cristal de algunos de los aislamientos nativos de Bacillus thuringiensis obtenidos en el PNN Katios. Línea 1 Marcador de Peso Molecular, Líneas 2 y 3 cepas control BtHD1 y Bt cry3A respectivamente. Líneas 4-10 aislamientos nativos IBUN89A2, IBUN 89A4, IBUN 89A6, IBUN89A8, 89A10, IBUN91B1 y IBUN93B1.
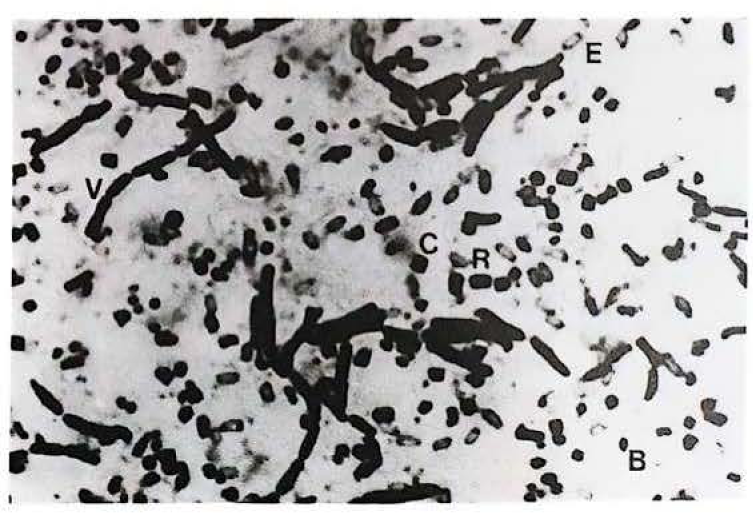
Visualización microscópica de la cepa nativa de Bacillus thuringiensis IBUN115.A32. Tres tipos diferentes de cristal paraesporal. B, bipiramidal; C cuadrado; R rectangular; esporas (E) y células vegetativas (V).
Análisis electroforético de proteínas de cepas nativas de Bacillus thuringiensis
El patrón electroforético de las proteínas asociadas al cristal permitió descartar cepas provenientes de la misma muestra de suelo, especialmente de algunas muestras del PNN Katios, las cuales se consideraron como idénticas ya que presentaron una gran similitud en términos del perfil de proteínas en geles de poliacrilamida (Fig. 1). Por esta razón, del total de 29 cepas obtenidas, se emplearon 12 para una caracterización más detallada. La posibilidad de identificar cepas iguales, provenientes de una misma muestra de suelo, es una herramienta que permite evitar la duplicación de análisis con cepas que con un alto nivel de certeza son iguales entre si (Bravo et al. 1998).
De las cepas de Bt obtenidas a partir de las muestras de ecosistemas agrícolas, es importante resaltar además de la cepa IBUN115A32, las cepas IBUN97B e IBUN104B62, los cuales mostraron un perfil electroforético definido con tres bandas de alrededor de 20, 45 y 60 kDa para la primera y 100 kDA para la segunda, lo cual no es un comportamiento típico (Hofte y Witley 1989; Feitelson y Kim 1992; Quiñones y Quintero 1996).
Análisis de genes cry por PCR
Las 12 cepas seleccionadas por su cristal paraesporal y su patrón electroforético se analizaron por PCR con oligonucleótidos generales para determinar la presencia de la familia de genes cry que codifican para la d-endotoxina con actividad biopesticida potencial (genes cry1) y contra coleópteros (cry3, cry7 y cry8). Para esta reacción de PCR se emplearon como controles la cepa HD1 y cepas que codifican para los genes cry3Aa, cry3Ba, cry3Ca y cry7Aa respectivamente (Fig. 4). Los resultados de este análisis mostraron que de las cepas seleccionadas, únicamente la cepa IBUN115A32 amplificó una banda alrededor de 300 pares de bases (pb), la cual corresponde a genes cry1 con actividad contra lepidópteros, de acuerdo con lo señalado por Cerón et al. (1995), mientras que la cepa IBUN91A1 amplificó con los oligos generales para genes cry con actividad a coleópteros, una banda alrededor de 700 pb. Particularmente la cepa IBUN97B12 amplificó con este mismo set de oligos, una banda de un peso molecular diferente al esperado, de alrededor de 160 pb (Fig. 4), lo cual posiblemente esté indicando la presencia de un nuevo gen cry perteneciente a la familia de genes que codifican proteínas con actividad en coleópteros, aunque análisis más detallados serán realizados para corroborar este resultado.
La gran mayoría de estudios dirigidos hacia la búsqueda de nuevas cepas de Bt muestra que la familia de genes cry, predominante en el ambiente, es aquella que confiere actividad contra lepidópteros (Martin y Travers, 1989; Chilcott y Wigley 1993; Bernhard et al. 1997; Carozzi et al. 1991). Un alto porcentaje de estos estudios además posee sistemas in vivo para la determinación de dicha actividad biopesticida, eso significa que evalúan las diferentes colecciones de Bι, contra una serie de insectos que en el mejor de los casos presentan máximo 3 especies diferentes de cada orden de insecto (Bernhard et al.. 1997). El sistema que se empleó en este estudio permite detectar por PCR, con un alto grado de confianza, todos aquellos genes con actividad a lepidópteros y coleópteros (excepto cry1Ha, cry1Ja, cry1Ka, cry8Ba y cry8Ca). De acuerdo con esto se puede decir que se cuenta con un sistema más sensible para detectar dichos genes que los estudios in vivo antes mencionados. A pesar de esto, únicamente se encontró un aislamiento portador de genes cry1 y 2 con actividad potencial a coleópteros, es importante resaltar que son pocos los registros en la literatura sobre genes-protemas con actividad contra coleópteros. El resto de cepas que no reaccionaron positivamente con los «primers» generales para los genes cry evaluados poseen otro tipo de genes cry que deben ser evaluados más en detalle ya que podrían tener interés en otras áreas como el control de insectos vectores de enfermedades. Los resultados obtenidos confirman la necesidad de realizar estudios más amplios de este tipo en nuestro país, con el ánimo de conocer el verdadero potencial de nuestra biodiversidad microbiana.
Es importante mencionar que la cepa IBUN115A32, única cepa encontrada con genes cry1, se evaluó en ensayos in vivo contra larvas de primer ínstar de Spodoptera frugiperda, Trichloplusia ni y Heliothis virescens no encontrándose actividad contra estos insectos de interés (resultados no mostrados).
Distribución de tipos de inclusiones paraesporales por lugar de origen.
Clasificación de tipos morfológicos de inclusiones paraesporales:
A. amorfo; A-B. amorfo - bipiramidal: A-T, amorfo - triangular;
B-C-R. bipiramidal, cuadrado, rectangular.
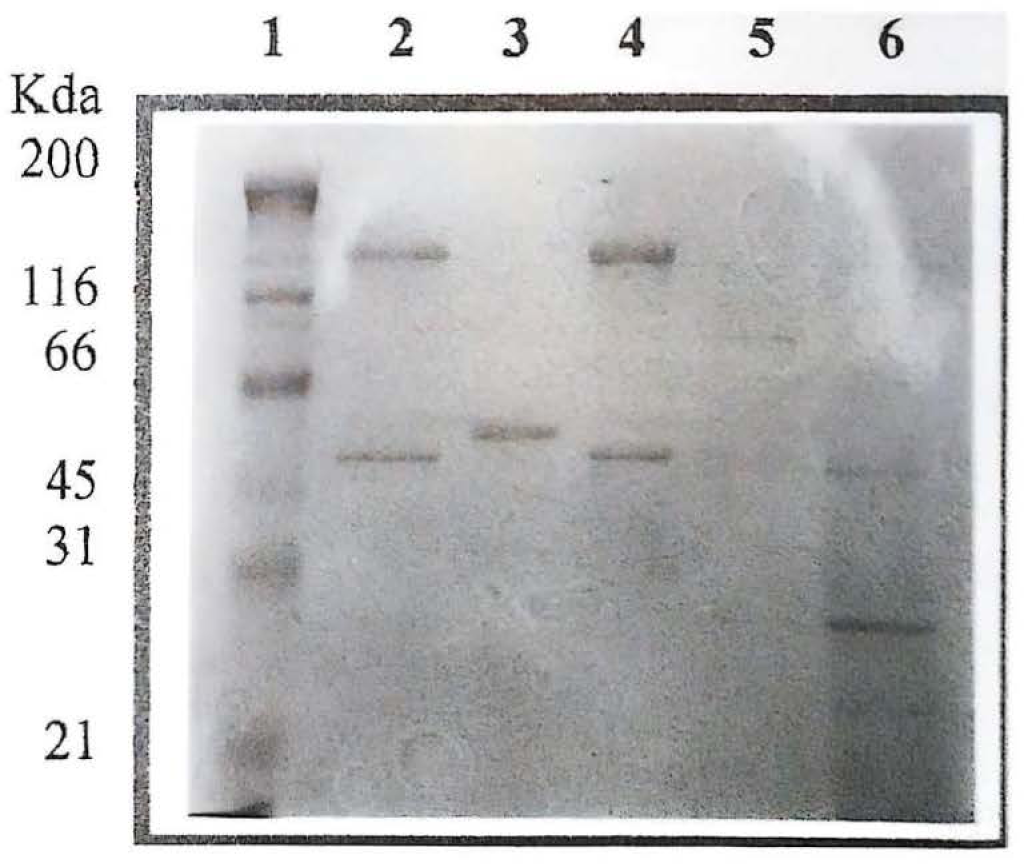
Perfil electroforético (SDS-PAGE) de las proteínas asociadas al cristal de los aislamientos nativos de B thuringiensis. Línea 1 Marcador de Peso Molecular. Línea 2 y 3 aislamientos control Bt HD1 y cry3A respectivamente; Línea 4 IBUN115A32; Línea 5 IBUN104B62; Línea 6 IBUN97B1.
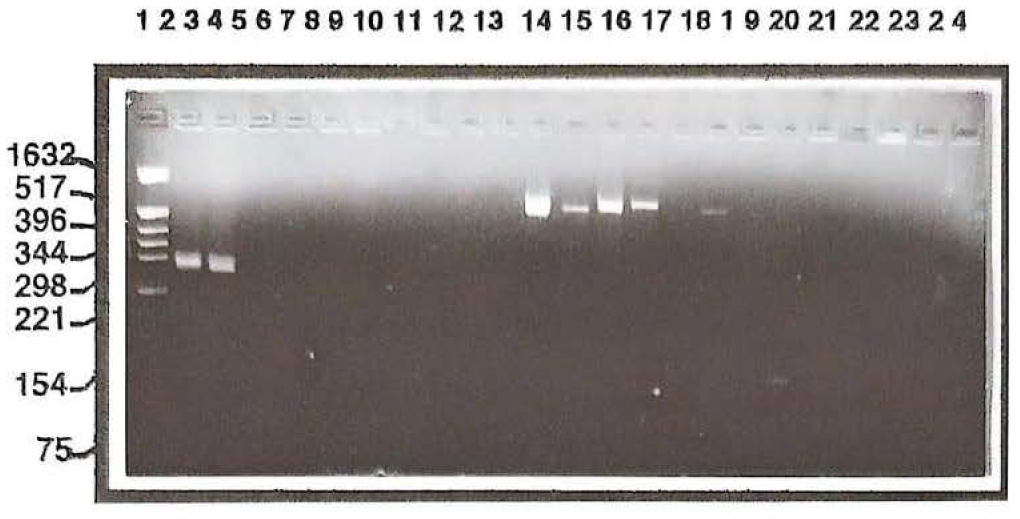
Electroforesis en gel de agarosa de los productos de la PCR obtenidos con los oligos generales para genes con actividad contra lepidopteros genes cry1 (líneas 2 al 11) y con actividad contra coleópteros genes cry3, cry7 y cry8 (líneas 12 a 24). Línea 1 Marcador de Peso Molecular, Línea 2 Cepa control para genes cry1 HD1, Línea 3 y 24 aislamiento IBUN115A32, Línea 4 y 18 IBUN93B1, Línea 5 y 19 IBUN97B12, Línea 6 y 23 IBUN104.54, Línea 7 y 22 IBUN98A4, Línea 9 y 20 IBUN94B2, Línea 10 y 17 IBUN91A1, Línea 11 y 16 IBUN 89A12. Líneas 12 a 15 cepas control para genes con actividad contra coleópteros (genes cry3Aa1, cry3Ba1, cry3Ca1 y cry7Aa1 respectivamente).
Conclusiones
La metodología presentada permite obtener de forma relativamente rápida y con alto nivel de caracterización, aislamientos nativos de Bt provenientes de muestras de suelo. Dentro de este diseño experimental se deben resaltar dos elementos, que llevan a la optimización del tiempo así como a los recursos empleados, variables estas muy importantes al momento de realizar estudios de este tipo: En primer lugar, la selección de aislamientos idénticos, provenientes de la misma muestra de suelo, permite evitar el realizar dos o más veces el análisis de un mismo aislamiento. En segundo lugar, el empleo de la técnica de PCR optimiza la selección de los aislamientos que deberán ser evaluados en los bioensayos in vivo, empleando sólo aquellos aislamientos que realmente tengan un potencial concreto de presentar actividad contra un insecto blanco de interés.
Las muestras de suelo evaluadas, provenientes tanto de bosques naturales como de ecosistemas agrícolas, son una buena fuente de aislamientos de Bt, especialmente en términos de la diversidad biológica, asociada a los factores de patogenicidad de este microorganismo, como son los cuerpos de inclusión, las proteínas que conforman dichos cuerpos y por su puesto los genes que codifican para estas últimas. Las muestras del PNN Katios se destacan en este sentido por la cantidad de cepas obtenidas y lo atípico de sus patrones en cada una de las características evaluadas (cristal, perfil de proteínas y genes cry por PCR), lo cual confirma que los bosques naturales son una buena fuente de diversidad microbiana que debe ser estudiada y conocida como parte del patrimonio nacional. Cabe mencionar además que el ecosistema agrícola es igualmente un buen lugar para buscar dicha diversidad si se consideran las cepas IBUN115A32 y IBUN97B12 las cuales también se alejan de los patrones típicos para la especie.
Agradecimientos
Para la realización de este trabajo se han obtenido recursos de COLCIENCIAS mediante la financiación del proyecto 1101-12-339-95 y del Instituto de Biotecnología de la Universidad Nacional de Colombia. Los autores desean agradecer sus importantes aportes.