
Abstract
The art for rearing insects that produce silk and the utilization of the latter is known as Sericulture. Bombyx mori (L.) is the insect that produces the largest amount of silk. This insect is attacked by protozoa, bacteria, viru- ses and fungi. Once the coffee berry borer Hypothenemus hampei (Ferrari) was establi- shed in Colombia, some research was con- ducted in order to control this pest by using entomopathogenic fungi. As a result of this strategy, there is some risk related to the effect of spraying the fungi on coffee planta- tions, just near the place where the mulberry crop is established, since the latter, is the only food consumed by the silkworm through- out its life span. The main aim of this ex- periment was to determine the differences in pathogenicity of four isolates of B. bassiana to the silkworm. The isolates Bb-9002, Bb- 9022, Bb-9105 and Bb-9107 from H. hampei, Nephotettix cincticeps (Ulher), Perileucop- tera coffeella (Guérin - Méneville) and B. mori, respectively, were used. Groups of 50 larvas (five replicates) were inoculated with a concentration of 1x107 conidia/ml, of each isolate, respectively. During the evaluation, the mortality caused by a virus, which produ- ces the Nuclear Polyhedrosis (VNP) was registered. Larvae inoculated with the isolate Bb-9107 showed the highest percentage of mortality (10%), followed by the isolate Bb- 9105. Both isolates were obtained from lepi- doptera. The highest percentage of viral mor- tality observed was also on larvae infected with the isolate Bb-9107.
Introducción
La técnica para la cría de insectos que producen seda y el aprovechamiento de esta fibra para uso humano se conoce como Sericicultura (García 1970). Esta industria tuvo origen en China 2.600 A.C., se extendió al Japón en el siglo III D.C. y alcanzó su máximo apogeo en Francia en 1853 (Bertin 1980). Según Perea (1988), la sericicultura llegó a Colombia en 1868 traída por el médico Manuel Vicente de la Roche.
El insecto que da mayor rendimiento en la producción de seda se conoce como «gusano de seda», Bombyx mori (L.) (Lepidoptera: Bombycidae), aún cuando existen otros lepidópteros de la familia Saturniidae como Antheraea pernyi, A. mylita, A. yama-mai, que también son buenos productores de seda.
Dentro de los diferentes patógenos que afectan la cría del gusano de seda, según Krishnaswami et al. (1973), se encuentran: protozoarios, de los cuales el más importante es Nosema bombysis Nag., causante de la enfermedad conocida como Pebrina; bacterias como Streptococcus bombycis Sart. & Pacc. y S. pastorianus causantes de la Flacherie o « flacidez del gusano de seda»; virus causantes de enfermedades como la Poliedrosis citoplasmática y la poliedrosis nuclear o « Grasserie», transmitidas generalmente por ingestión de alimento contaminado con el virus o por transmisión transovárica; hongos causantes de enfermedades conocidas como Muscardinas o ««tizas»» dentro de los que se encuentran Beauveria bassiana (Bals.) Vuill., Metarhizium anisopliae (Metch.) Sorokin, Paecilomyces sp., Aspergillus sp., Penicillium sp., entre otros. Algunos estudios han clasificado estas enfermedades según la incidencia que tienen sobre una cría de gusano de seda, encontrando que las enfermedades en larvas alcanzan una incidencia del 17%., dentro del cual la Grasserie predomina, causando un 38,5% del total de las larvas enfermas, seguida por las Muscardinas con un 20,3% y la Flacherie con 11,9%. (Chae et al. 1984).
Con la llegada de la broca del café, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) a Colombia, en 1988, y con las investigaciones relacionadas con los diferentes métodos para su control, dentro de los cuales está el control biológico, basado principalmente en el uso del hongo entomopatógeno B. bassiana (Bustillo 1991), se han creado expectativas sobre el efecto que puedan tener las aspersiones del hongo en los cafetales atacados por la broca sobre la contaminación con conidias de los cultivos aledaños de morera (Morus indica), fuente de alimento de las larvas de B. mori, lo que se convertiria en un obstáculo para el desarrollo de la sericicultura, la cual ha mostrado ser promisoria en el Programa de Diversificación de Cultivos de la Federación Nacional de Cafeteros.
De esta forma, el objetivo de este estudio fue determinar si existe diferencia, en cuanto a patogenicidad se refiere, de los aislamientos de B. bassiana en el gusano de seda. Es importante anotar que este estudio hace parte de un proyecto encaminado a establecer el impacto de B. bassiana sobre crías del gusano de seda y que los resultados obtenidos son preliminares.
Materiales y Métodos
El experimento se realizó en el laboratorio de Patología de Insectos y en la unidad de cría de gusano de seda de CENICAFE en Chinchiná (Caldas). Para la realización del experimento se utilizaron larvas del gusano de seda de tercero, cuarto y quinto instares, suministradas por la empresa « Sedas Colombianas y Coreanas» (Cokosilk) ubicada en Pereira. Como alimento, para cada grupo de insectos en cada instar, se utilizaron hojas de morera (Morus indica var. kanva-2), ya que esta variedad responde muy bien a las condiciones climáticas y edáficas de la zona cafetera, obteniéndose buenos rendimientos de hoja (Osorio et al. 1990). Los aislamientos de B. bassiana utilizados fueron Bb-9002, Bb-9022, Bb-9105, Bb-9107, los cuales fueron aislados a partir de H. hampei, Nephotettix cinctipes (Uhler) (Homoptera: Cicadellidae), Perileucoptera coffeella (Guérin-Méneville) (Lepidoptera: Lyonettidae) y B. mori, respectivamente, y fueron suministrados por el laboratorio de Patología de Insectos.
El medio de cultivo utilizado para la preservación de los aislamientos de B. bassiana fue el Agar- Sabouraud-Dextrosa (SDA) acidificado con ácido láctico (44%) al 1%, y el antibiótico cloranfenicol al 0,1% para evitar la proliferación bacteriana.
Para facilitar la remoción y el lavado de las conidias se utilizó una mezcla de agua destilada estéril y Tween 80 al 0,5%.
Obtención del inóculo
Una vez obtenido el crecimiento masivo y esporulado de cada aislamiento, en cajas de petri, se procedió a adicionar un volumen final de 50 ml de la mezcla de agua destilada estéril y Tween 80 en volúmenes de 10 ml. Con la ayuda de un pincel estéril se realizó la remoción de las conidias, facilitando su recolección en un beaker de 250 ml (solución madre). Con el producto de lavado de las conidias se procedió a hacer diluciones seriadas en tubos de ensayo, adicionando 1 ml de esta solución madre en 9 ml de agua destilada estéril (dilución 10-1) y repitiendo el mismo procedimiento hasta llegar a la dilución 10-3. A partir de esta dilución y con la ayuda de un hemocitómetro (cámara de recuento) y un microscopio de luz, se realizó el recuento de conidias (40x).
Recuento de conidias
Con 10 µl se llenan los dos compartimentos de la cámara de recuento, colocando una laminilla de cuarzo para observar al microscopio. El recuento de conidias se llevó a cabo en el cuadrante central de 25 cuadros (área utilizada para el recuento de glóbulos rojos). Este montaje se realiza tres veces (seis lecturas), obteniéndose un promedio, el cual se multiplica por el factor de corrección de la cámara de recuento (104) y por el recíproco de la dilución que se utilizó para realizar el recuento (103).
Porcentaje de germinación
Para cuantificar la viabilidad del inóculo, antes de entrar en contacto con las larvas, se realizó la siguiente prueba con cada uno de los aislamientos: de la dilución 10−3 se tomaron siete alícuotas de 4 ml cada una y se depositaron en cajas de petri con SDA, marcando previamente los sitios donde serían colocadas las alícuotas. Las cajas de petri se llevaron a incubación por 24 horas (luz constante) a 23°C. Al cabo de este tiempo, el trozo de agar previamente marcado con la alícuota se seccionó y se colocó en una lámina de vidrio, se le adicionó azul de lactofenol y se hizo el montaje con una laminilla cubreobjetos. El porcentaje de germinación se obtuvo teniendo en cuenta el número de conidias germinadas y el número total de conidias. De cada alícuota se observaron cinco campos para un total de cinco porcentajes de germinación, los cuales se promediaron para obtener el «promedio del porcentaje de germinación». Como conidia germinada se considera aquella que presente una longuitud del tubo germinal de aproximadamente el doble o más del diámetro de la conidia.
Infección de larvas
Todas las larvas de tercero, cuarto y quinto instar, utilizadas para el experimento, se recibieron en el período de dormancia de segundo a tercer instar. Al segundo día del tercer instar, se seleccionaron las larvas que serían infectadas con cada uno de los inóculos de los aislamientos, dejando un número igual de larvas, para el cuarto y quinto instares, que fueron sometidas al mismo tratamiento.
Una vez obtenida la concentración de la solución madre (para los cuatro aislamientos), se le adicionó agua destilada estéril hasta obtener la concentración deseada del inóculo (107 conidias/ml). Posteriormente, cada larva se sumergió en esta solución, hasta que quedaron totalmente impregnadas. Luego, un total de 50 larvas se llevaron a bandejas metálicas de 5 x 28 x 34 cm, las cuales se colocaron en estantes metálicos, y allí se observó el desarrollo de las larvas. Las larvas en el cuarto instar se infectaron el primer día de ese instar y las de quinto, en el segundo día.
La evaluación se realizó dos veces al día, desde el momento de la infección hasta que el 80% de las larvas del testigo encapullaron. Las condiciones ambientales a través del experimento fueron H.R.= 75-90%.; T=23-25°C. Estas condiciones son las requeridas para el desarrollo del insecto, pero varían dependiendo del instar. Durante este período se retiraron las larvas muertas por el hongo. Las larvas que presentaron el signo del ataque del hongo (Fig. 1 y 2) se llevaron a cámara húmeda para facilitar el desarrollo del hongo, y luego se procedió a la identificación microscópica.

A. Larva de B. mori con signos de la enfermedad causada por B. bassiana, B. Larva de B. mori con signos de la enfermedad causada por el Virus de Poliedrosis Nuclear, VPN.
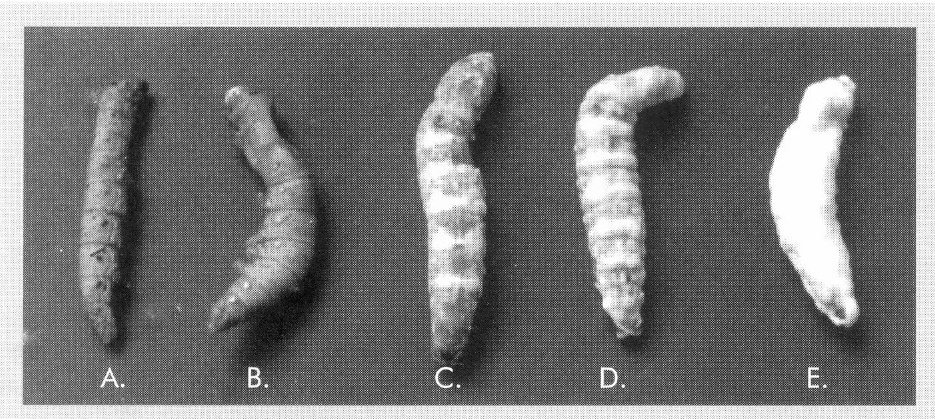
Desarrollo del hongo Beauveria bassiana sobre larvas de Bombyx mori. A. endurecimiento y coloración rojiza de la larva infectada por el hongo; B. aparición de micelio en la cabeza, patas anteriores, patas posteriores y espiráculos; C. cubrimiento parcial de los anillos de la larva por el hongo; D. cubrimiento de la totalidad del cuerpo de la larva por el hongo; E. esporulación del hongo en la larva atacada.
Además se registró la mortalidad causada por otros agentes tales como el virus de la «Poliedrosis Nuclear» (VPN), el cual se caracteriza por causar un aumento anormal en el tamaño de los anillos (segmentos) de la larva, dando una apariencia acuosa de ellos; posteriormente, toda la larva toma una coloración parduzca, se produce una licuefacción completa de su interior y hay secreción de un exudado de coloración oscura (Fig.1). Estos exudados se recolectaron de larvas moribundas y se observaron directamente al microscopio de luz para evidenciar la presencia de estructuras poliédricas, típicas del agente causal de dicha enfermedad.
El efecto de los aislamientos sobre la mortalidad en las larvas de B. mori se evaluó a través de las tasas de crecimiento en la mortalidad de las larvas por medio de una regresión lineal simple. El promedio de mortalidad para cada punto muestral, en cada tratamiento, incluyendo el testigo, provino de cinco repeticiones. A su vez, cada repetición estuvo formada por grupos de 50 larvas colocadas en las bandejas metálicas (unidad experimental). La mortalidad se determinó con base en el número de insectos muertos, respecto al número de insectos por unidad experimental. El efecto de los tratamientos se evaluó a través de un análisis de varianza y la prueba de comparación de dichos coeficientes se hizo por medio de la prueba de «t» para comparación de promedios.
Resultados
Obtención del inóculo y recuento de conidias
El recuento de conidias inicial para los aislamientos Bb-9002, Bb-9022, Bb-9105 y Bb-9107 varió entre 2 x 108 y 5 x 108 conidias/ml. Esto se llevaron a una concentración de 1 x 107 conidias/ml para con estos inóculos asperjar los diferentes grupos de larvas.
Porcentaje de germinación
Los resultados de porcentaje de germinación se presentan en la Tabla 1.
Porcentanje de germinación inicial de los aislamientos de Beauveria bassiana, utilizados en la infección de larvas de Bombyx mori.
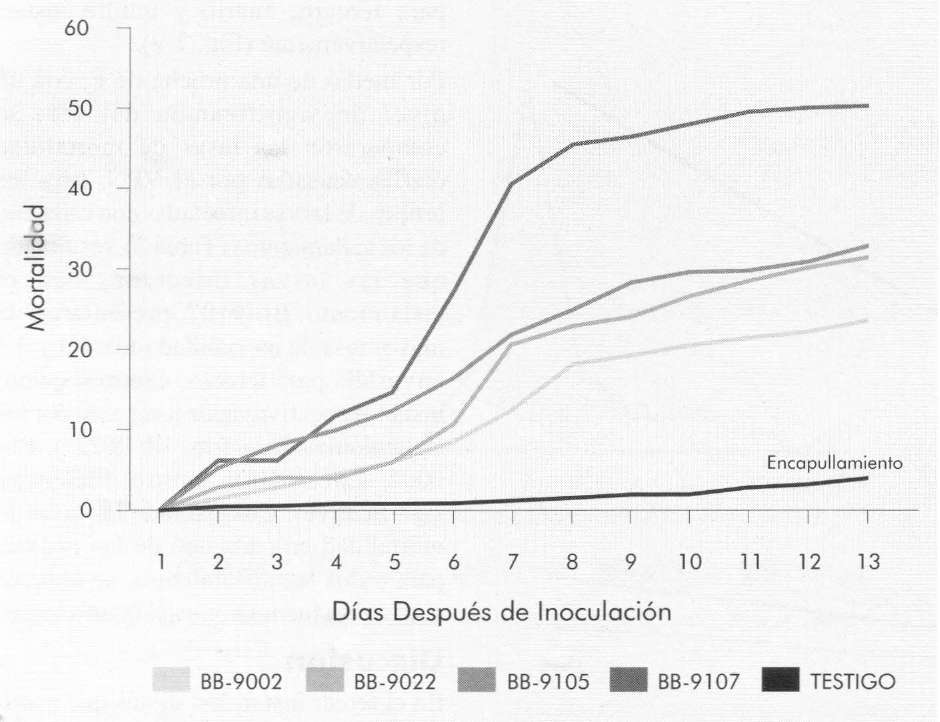
Las Figuras 3 y 4 muestran la mortalidad diaria y acumulada, respectivamente, de cada uno de los aislamientos de B. bassiana en las larvas de tercer instar de B. mori. La mortalidad se presentó a partir del sexto día después de la infestación de las larvas con el hongo. Por medio de una prueba de F, con un nivel de significancia del 1%, se compararon las tasas de mortalidad diarias para cada uno de los aislamientos (Tabla 2). El aislamiento Bb-9107 presentó la mayor tasa de mortalidad (1,07 larvas/día), seguido por los aislamientos Bb-9105, Bb-9002 y Bb-9022 respectivamente, a la concentración de 1 x 107 conidias/ml, presentando diferencias significativas. La Figura 5 muestra los ajustes de regresión donde se observa dicha tendencia. Se realizó una prueba de «t» (p=0,01) para comparar las pendientes de las regresiones para cada aislamiento, y se observaron diferencias significativas entre todas las comparaciones, a excepción de la comparación entre los aislamientos Bb-9002 y Bb-9105.
Tasa diaria del porcentaje de mortalida causada por Beauveria bassiana en larvas de Bombyx mori, infectadas en tercer instar.
Diferencias significativas
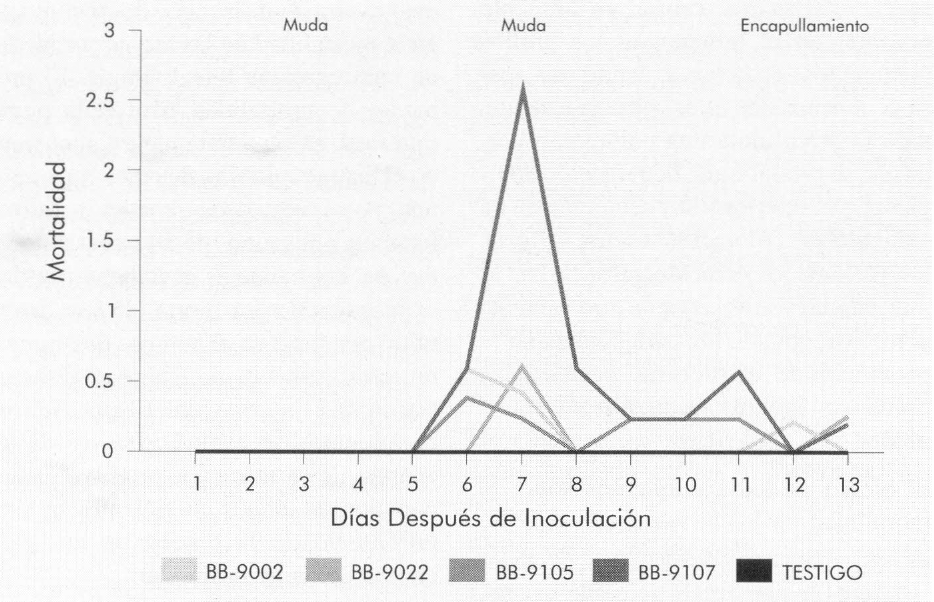
Porcentaje de mortalidad media diaria en larvas de B. mori de tercer instar causada por B. bassiana.
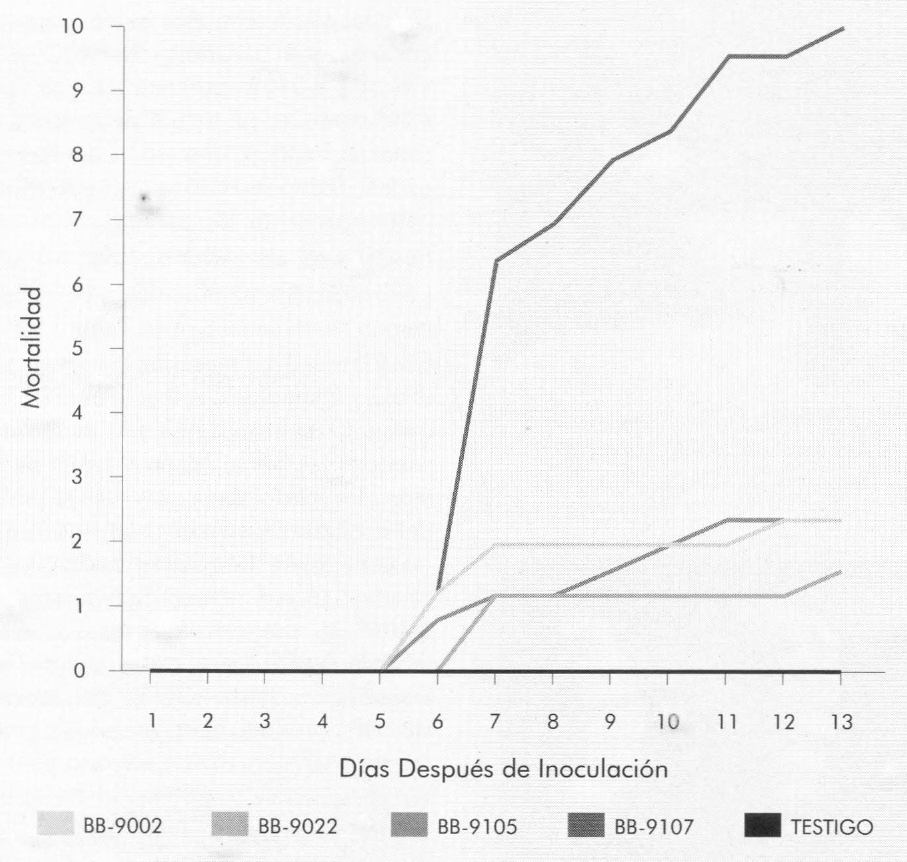
Porcentaje de mortalidad diario acumulado causado por B. bassiana en larvas de tercer intar de B. mori.
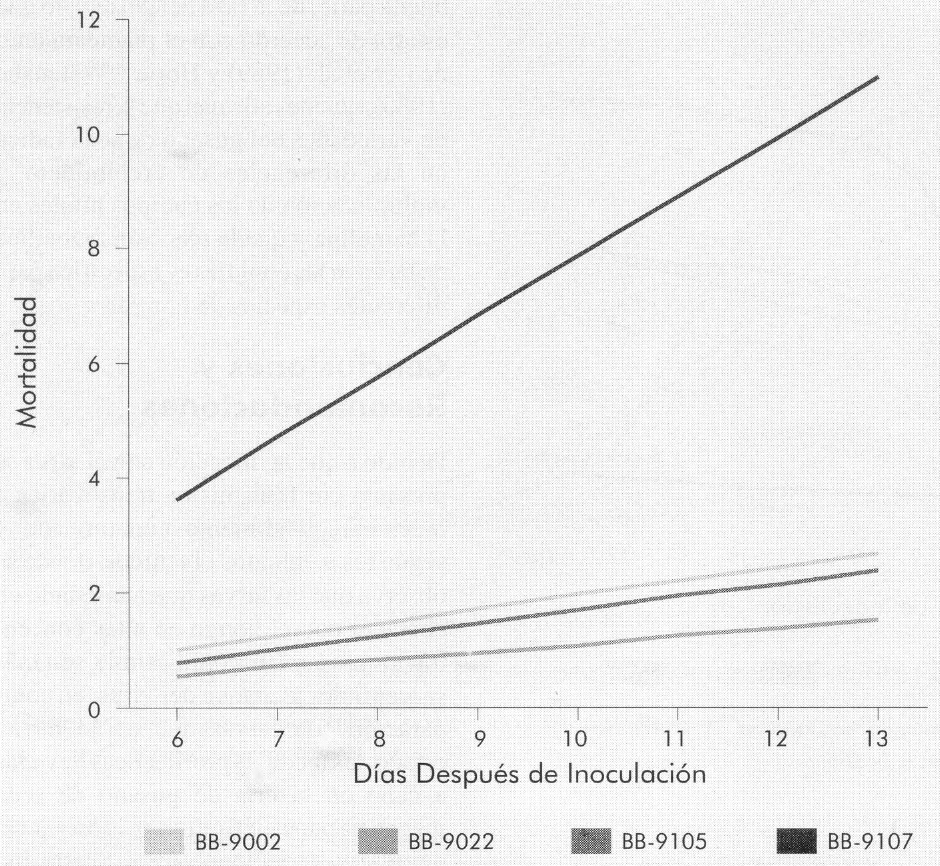
Ajuste de regresión del porcentaje de mortalidad causado por B. bassiana en larvas de B. mori de tercer instar.
La Figura 6 muestra el porcentaje de mortalidad diaria acumulada en larvas de cuarto instar causada por cada uno de los aislamientos de B. bassiana, observándose que el único aislamiento que causó mortalidad en las larvas de B. mori fue el Bb-9107 (5,2%). La mortalidad presentada por el testigo (0,4%) puede atribuirse a contaminación durante el experimento o al efecto del hongo presente en el medio ambiente, ya que este entomopatógeno se encuentra normalmente en el campo.
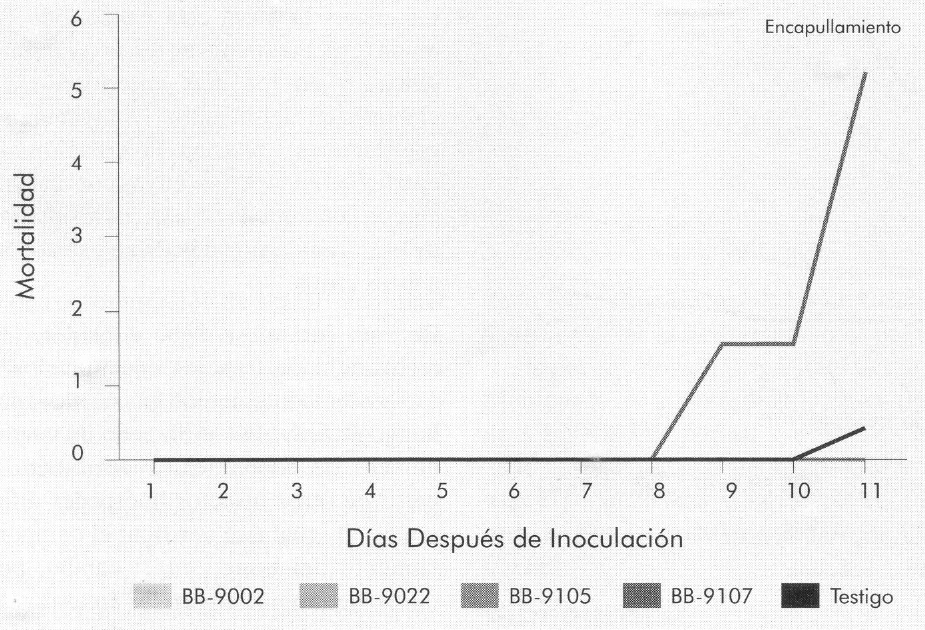
Porcentaje de mortalidad diario acumulado causado por B. bassiana en larvas de cuarto instar de B. mori.
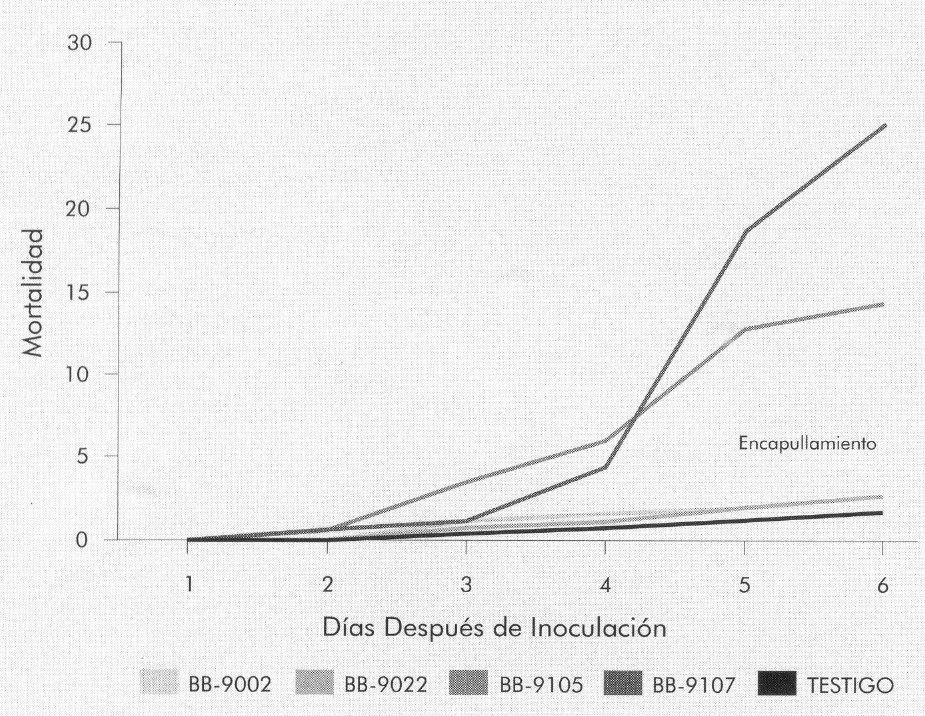
Las Figuras 7 a 9 muestran la mortalidad acumulada causada por el VPN, registrada para cada uno de los instares evaluados. En el tercer instar (Fig. 7), esta mortalidad se presentó a partir del segundo día después de inoculación (para todos los aislamientos), con una diferencia de cuatro días respecto a la aparición de los signos de muerte causada por el hongo (Fig. 2 y 4).
Porcentaje de mortalidad diario acumulado, causado por VPN, en larvas de B. mori de tercer instar, infectadas con B. bassiana.
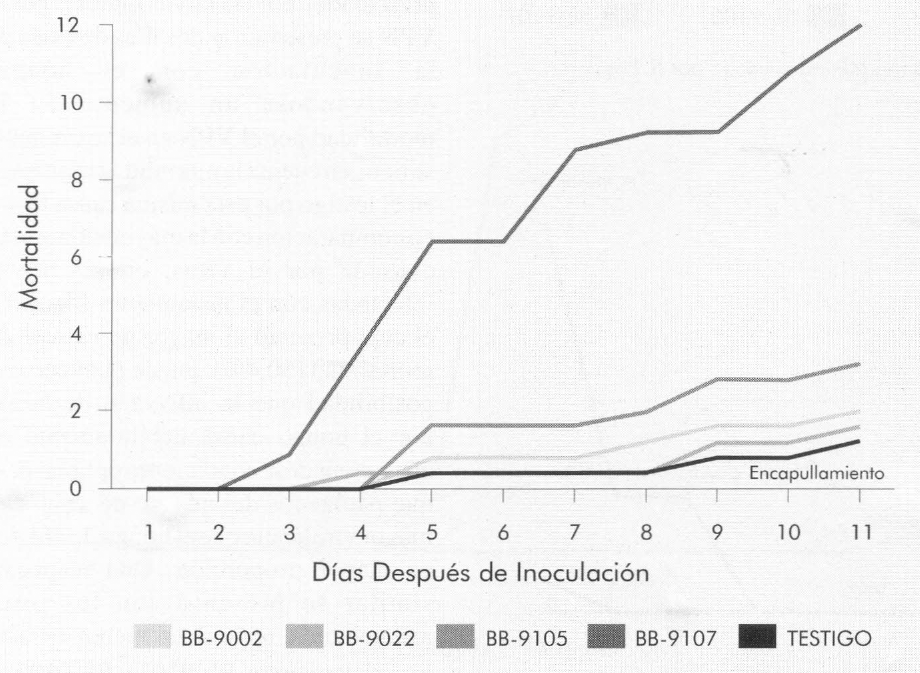
Porcentaje de mortalidad diario acumulado, causado por VPN, en larvas de B. mori de cuarto instar, infectadas con B. bassiana.
Porcentaje de mortalidad diario acumulado, causado por VPN, en larvas de quinto instar de B. mori, infectadas con B. bassiana.
Para las larvas de cuarto instar infestadas con el aislamiento Bb-9107, la mortalidad por VPN comenzó a registrarse partir del tercer día después de inoculación, con una diferencia de cinco días respecto a la aparición de los signos de muerte causada por hongo (Fig. 6 y 8). Para el resto de aislamientos, esta mortalidad se registró a partir del quinto día.
Para las larvas infectadas en el quinto instar, la mortalidad por VPN comenzó a presentarse a partir del segundo día después de infectar las larvas con cada uno de los aislamientos del hongo (Fig. 9).
La mortalidad por VPN presentada en el testigo para larvas del tercero, cuarto y quinto instar fue de 4%, 1,2% y 1,6%, respectivamente, mientras que el mayor porcentaje de mortalidad en los instares evaluados correspondió a las larvas infectadas con el aislamiento Bb-9107, con porcentajes de 50,4%, 12,4% y 25,2% para tercero, cuarto y quinto instar, respectivamente (Fig. 7-9).
Por medio de una prueba de F, con un nivel de significancia del 1%, se compararon las tasas de mortalidad diarias causadas por el VPN, para los grupos de larvas infectadas con cada uno de los aislamientos (Tabla 3), resultando que las larvas infectadas con el aislamiento Bb-9107 presentaron la mayor tasa de mortalidad (4,9; 1,3 y 5,3 larvas/día para tercero, cuarto y quinto instar, respectivamente), seguido por los aislamientos Bb-9105, Bb-9022 y Bb-9002. Este análisis mostró diferencias significativas. Comparando las tasas de mortalidad en cada uno de los instares para todos los aislamientos, se observó que la tasa fue menor en el cuarto instar.
Tasa diaria del porcentaje de mortalidad causada por el virus de la poliedrosis nuclear, en larvas de Bombyx mori de tercero, cuarto y quinto instar, infectadas con Beauveria bassiana.
Diferencias significativas
Discusión
En el tercer instar, los signos que manifiestan las larvas muertas por el hongo se presentaron a partir del sexto día después de la inoculación, mientras que los signos presentados por las larvas muertas por el VPN se presentaron dos días después de la inoculación con el hongo, observándose un aumento en la mortalidad por el VPN en el sexto día; si se tiene en cuenta la mortalidad registrada en el testigo por esta misma causa (4%), en comparación con la mayor mortalidad causada por el virus, en las larvas infectadas con el aislamiento Bb-9107, el cual presentó el mayor porcentaje de mortalidad (50,4%), puede plantearse la posibilidad que la infección de larvas por el hongo causa debilitamiento en ellas, creándose condiciones óptimas para que patógenos del gusano de seda, con mayor virulencia que el hongo, lo afecten en mayor proporción. Una respuesta similar se presentó con los otros aislamientos, aunque en menor escala. Los aislamientos Bb-9107 y Bb-9105 de B. bassiana (ambos aislados de lepidópteros) fueron más virulentos que los aislamientos Bb-9002 y Bb-9022, alcanzando un porcentaje de mortalidad máximo del 10 y 2,4, respectivamente, bajo las condiciones de la unidad de cría de gusano de seda de CENICAFE. Esto podría sugerir que el aislamiento del hongo proveniente del mismo insecto sobre el cual se desea conocer su efecto es más patogénico que los aislamientos del hongo obtenidos de insectos diferentes, susceptibles al ataque del entomopatógeno. De esta manera se plantea la existencia de cierta especificidad entre el hongo y su huésped, lo que estaría de acuerdo con los registros de especificidad de algunos aislamientos de B. bassiana hacia los grupos de insectos de los cuales el hongo ha sido aislado inicialmente (Goette y Johnson 1982; Horie y Watanabe 1980; Prior 1991). Además puede pensarse que la interacción hongo-gusano de seda depende también, en buena parte, de la raza del gusano, lo que estaría de acuerdo con el planteamiento de Lee et al. (1989) y Horie y Watanabe (1980), quienes afirman que la resistencia de variedades del gusano de seda radica en las diferencias de crecimiento y multiplicación de los cuerpos hifales en la hemolina y que la reacción fagocítica de larvas a las conidias es específica para diferentes especies de hongos.
Conclusiones Y Recomendaciones
Debido a que la infección con el virus se presenta por transmisión transovárica e ingestión de alimento contaminado y según los resultados obtenidos, donde se observa que las larvas que han estado en contacto con el hongo en altas concentraciones (1 x 107 conidias/ml), son más susceptibles al ataque del virus, en comparación con larvas que no lo hayan estado, las medidas profilácticas llevadas a cabo en la cría de gusano de seda juegan un papel definitivo en la presencia de enfermedades y su control. Si este tipo de medidas no se realizan normalmente, se deben implementar y si existen, se deben reforzar, sobre todo en los días en los cuales la larva está en período de muda (especialmente en tercero y quinto instar) y en los días anteriores a la elaboración del capullo. Estas medidas profilácticas evitarían la creación de epizootias en las crías, cuyas consecuencias serían impredecibles y los resultados de este trabajo no se podrían extrapolar a estos casos.
De esta forma, se debe considerar la realización de trabajos encaminados a esclarecer la interacción gusano de seda-hongo B. bassiana, utilizando un mayor número de alistamientos del hongo a partir de otros insectos huéspedes diferentes, al igual que verificar el comportamiento del hongo en el campo, por medio de monitoreos en las áreas donde se realicen las aspersiones del entomopatógeno.
Ya que no todos los aislamientos del hongo B. bassiana presentan la misma patogenicidad en B. mori y considerando el porcentaje máximo de mortalidad obtenido por el aislamineto Bb-9107 (10%), a una concentración de 1 x 107 conidias/ ml, se podría sugerir una coordinación de las prácticas serícolas y de las aspersiones del hongo, con el fin de no obstaculizar el desarrollo de ninguna de las dos importantes labores.
Para trabajos posteriores, el instar de la larva que se recomienda a trabajar es el tercero, ya que es el que permite apreciar completamente el efecto del hongo a través de todo su desarrollo.
Footnotes
Agradecimientos
El autor expresa sus agradecimientos al Dr. Alex Enrique Bustillo Pardey, Jefe de la Disciplina Entomología de CENICAFE, por su colaboración en el desarrollo del trabajo; a la Dra. Esther Cecilia Montoya R., de la Disciplina de Biometría de CENICAFE, por su asesoría en el análisis estadístico de los resultados y a todo el personal de auxiliares del Laboratorio de Patología de Insectos de CENICAFE.